
Chlorogenic Acid Inhibits Rahnella aquatilis KM25 Growth and Proteolytic Activity in Fish-Based Products


Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strain
2.2. Culture Conditions
2.3. Determination of the Antimicrobial Activity of Chlorogenic Acid
2.4. Morphology Analyses
2.5. RNA Isolation and RT–qPCR Analyses
2.6. In Situ Evaluation of Biopreservative Properties of Chlorogenic Acid
2.6.1. Preparation of Salmon-Based Samples
2.6.2. In Situ Antimicrobial Assay
2.6.3. Determination of TVB-N
2.6.4. Determination of TMA-N
2.7. Statistical Analysis
3. Results and Discussion
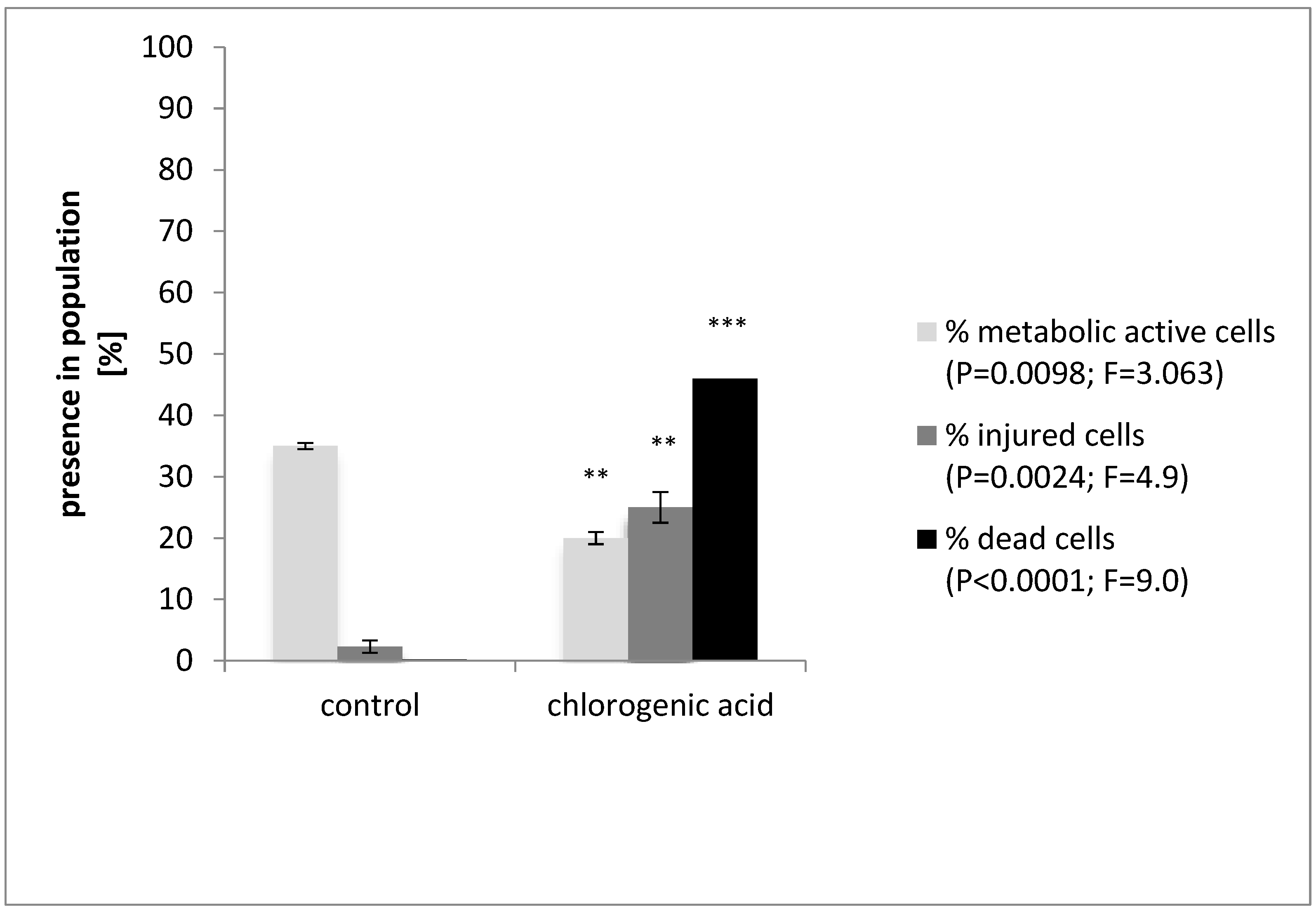
3.1. Antimicrobial Activity of Chlorogenic Acid
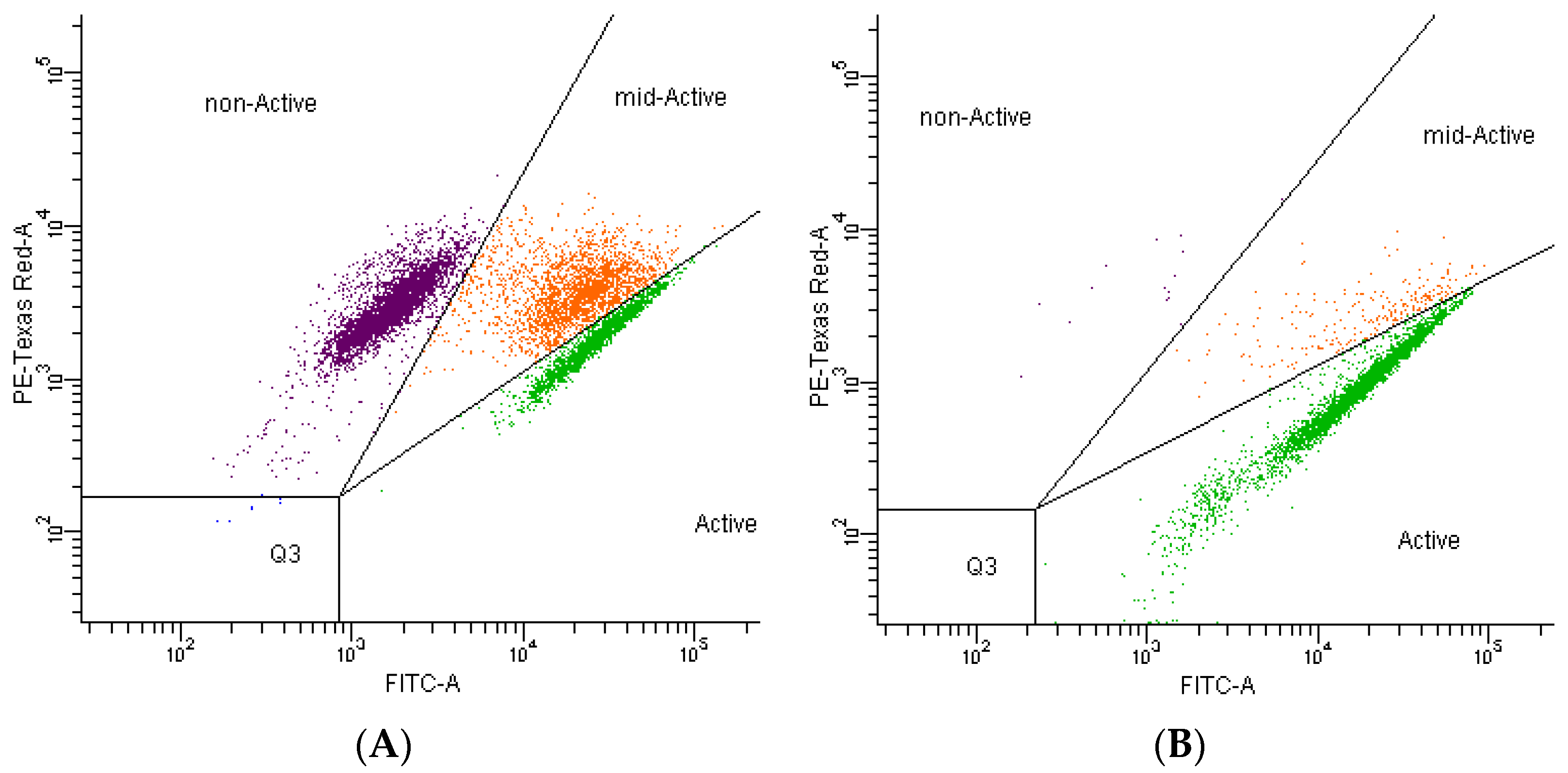
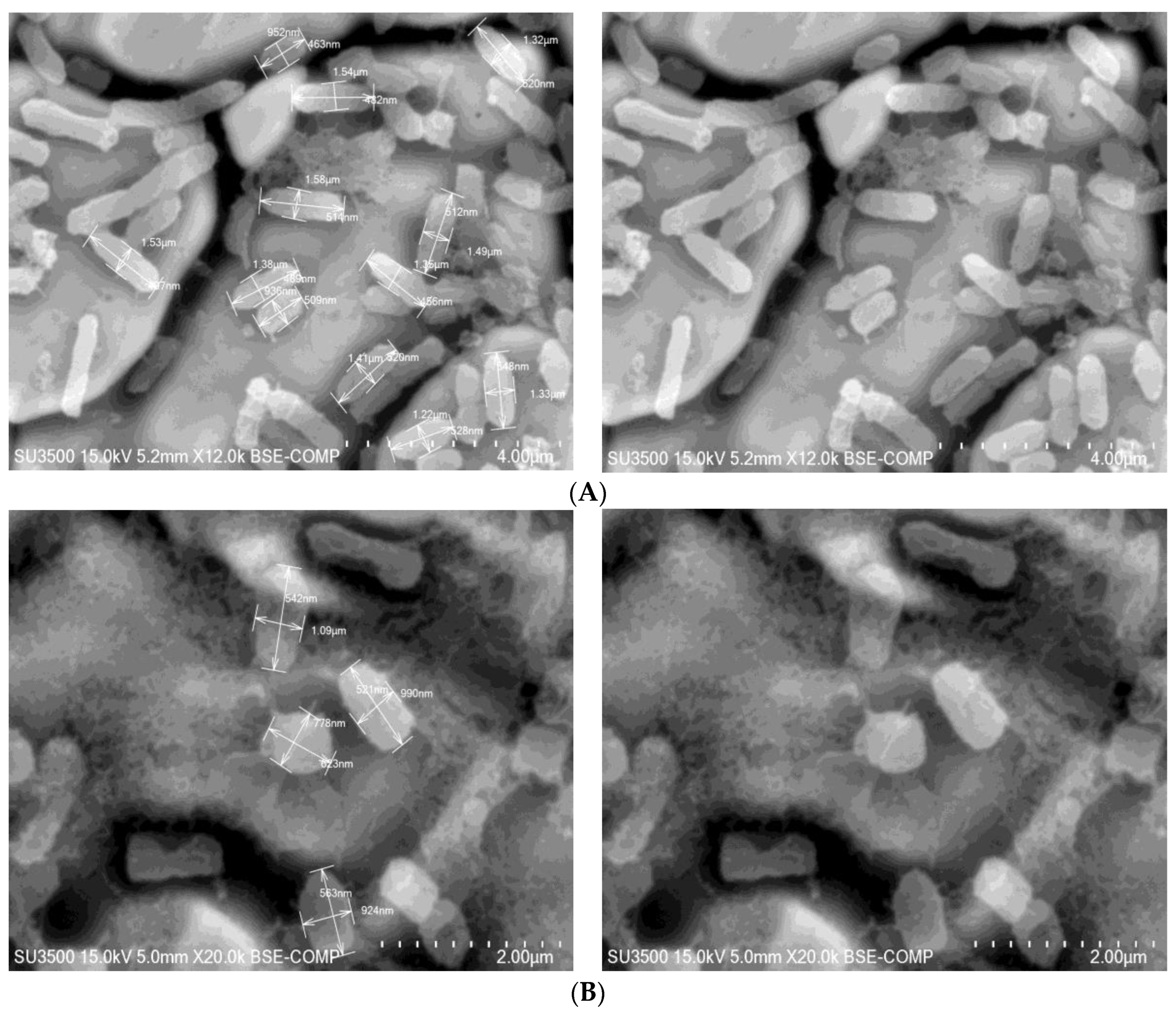
3.2. Effect of Chlorogenic Acid on Morphology of Cells
3.3. Effect of Chlorogenic Acid on ftsZ, ftsA, ftsN, tolB, and M4 Gene Expression
3.4. Effect of Chlorogenic Acid on R. aquatilis KM25 Growth in Salmon-Based Samples
3.5. Effect of Chlorogenic Acid on the Biochemical Parameters of Salmon-Based Samples
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Odeyemi, O.A.; Alegbeleye, O.O.; Strateva, M.; Stratev, D. Understanding spoilage microbial community and spoilage mechanisms in foods of animal origin. Compr. Rev. Food Sci. Food Saf. 2020, 19, 311–331. [Google Scholar] [CrossRef] [PubMed]
- Mikš-Krajnik, M.; Yoon, Y.-J.; Ukuku, D.O.; Yuk, H.-G. Volatile chemical spoilage indexes of raw Atlantic salmon (Salmo salar) stored under aerobic condition in relation to microbiological and sensory shelf lives. Food Microbiol. 2016, 53, 182–191. [Google Scholar] [CrossRef] [PubMed]
- Coppola, D.; Lauritano, C.; Esposito, F.P.; Riccio, G.; Rizzo, C.; de Pascale, D. Fish waste: From problem to valuable resource. Mar. Drugs 2021, 19, 116. [Google Scholar] [CrossRef] [PubMed]
- Paludan-Müller, C.; Dalgaard, P.; Huss, H.H.; Gram, L. Evaluation of the role of Carnobacterium piscicola in spoilage of vacuum- and modified-atmosphere-packed cold-smoked salmon stored at 5 °C. Int. J. Food Microbiol. 1998, 39, 155–166. [Google Scholar] [CrossRef]
- Skrodenyte-Arbaciauskiene, V.; Sruoga, A.; Butkauskas, D. Assessment of microbial diversity in the river trout Salmo trutta fario L. intestinal tract identified by partial 16S rRNA gene sequence analysis. Fish. Sci. 2006, 72, 597–602. [Google Scholar] [CrossRef]
- Liang, J.; Hu, X.; Lü, A.; Sun, J. First report on the characterization of pathogenic Rahnella aquatilis KCL-5 from crucian carp: Revealed by genomic and proteomic analyses. J. Fish Dis. 2020, 43, 889–914. [Google Scholar] [CrossRef]
- Botterweck, A.A.M.; Verhagen, H.; Goldbohm, R.A.; Kleinjans, J.; van den Brandt, P.A. Intake of butylated hydroxyanisole and butylated hydroxytoluene and stomach cancer risk: Results from analyses in the Netherland Cohort study. Food Chem. Toxicol. 2000, 38, 599–605. [Google Scholar] [CrossRef]
- Sun, Z.; Zhang, X.; Wu, H.; Wang, H.; Bian, H.; Zhu, Y.; Xu, W.; Liu, F.; Wang, D.; Fu, L. Antibacterial activity and action mode of chlorogenic acid against Salmonella enteritidis, a foodborne pathogen in chilled fresh chicken. World J. Microbiol. Biotechnol. 2020, 36, 24. [Google Scholar] [CrossRef]
- Li, Y.J.; Zhou, C.Y.; Qiu, C.H.; Lu, X.M.; Wang, Y.T. Chlorogenic acid induced apoptosis and inhibition of proliferation in human acute promyelocytic leukemia HL60 cells. Mol. Med.Rep. 2013, 8, 1106–1110. [Google Scholar] [CrossRef]
- Xu, J.-G.; Hu, Q.-P.; Liu, Y. Antioxidant and DNA-protective activities of chlorogenic acid isomers. J. Agric. Food Chem. 2012, 60, 11625–11630. [Google Scholar] [CrossRef]
- Liang, N.; Kitts, D.D. Role of chlorogenic acids in controlling oxidative and inflammatory stress conditions. Nutrients 2016, 8, 16. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef] [PubMed]
- Clinical and Laboratory Standards Institute. Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria That Grow Aerobically: Approved Standard, 9th ed.; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2012. [Google Scholar]
- Myszka, K.; Tomaś, N.; Wolko, Ł.; Szwengiel, A.; Grygier, A.; Nuc, K.; Majcher, M. In situ approaches show the limitation of the spoilage potential of Juniperus phoenicea L. essential oil against cold-tolerant Pseudomonas fluorescens KM24. Appl. Microbiol. Biotechnol. 2021, 105, 4255–4268. [Google Scholar] [CrossRef] [PubMed]
- Schmittgen, T.D.; Livak, K. Analyzing real-time PCR data by the comperative Ct method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]
- Goulas, A.E.; Kontominas, M.G. Effect of salting and smoking-method on the keeping quality of chub mackerel (Scomber japonicus): Biochemical and sensory attributes. Food Chem. 2005, 93, 511–520. [Google Scholar] [CrossRef]
- Ashie, I.N.A.; Smith, J.P.; Simpson, B.K. Spoilage and shelf-life extension of fresh fish and shellfish. Crit. Rev. Food Sci. Nutr. 1996, 36, 87–121. [Google Scholar]
- Zhuang, S.; Hong, H.; Zhang, L.; Luo, Y. Spoilage-related microbiota in fish and crustaceans during storage: Research progress and future trends. Compr. Rev. Food Sci. Food. Saf. 2021, 20, 252–288. [Google Scholar] [CrossRef]
- Wang, L.; Pan, X.; Jiang, L.; Chu, Y.; Gao, S.; Jiang, X.; Zhang, Y.; Chen, Y.; Luo, S.; Peng, C. The biological activity mechanism of chlorogenic acid and its applications in food industry: A review. Front. Nutr. 2022, 9, 943911. [Google Scholar] [CrossRef]
- Su, M.; Liu, F.; Luo, Z.; Wu, H.; Zhang, X.; Wang, D.; Zhu, Y.; Sun, Z.; Xu, W.; Miao, Y. The antibacterial activity and mechanism of chlorogenic acid against foodborne pathogen Pseudomonas aeruginosa. Foodborne Pathog. Dis. 2019, 16, 823–830. [Google Scholar] [CrossRef]
- Wu, Y.; Liang, S.; Zhang, M.; Wang, Z.; Wang, Z.; Ren, X. The effect of chlorogenic acid on Bacillus cereus based on metabolomics. Molecules 2020, 25, 4038. [Google Scholar] [CrossRef]
- Alikunhi, N.M.; Batang, Z.B.; AlJahdali, H.A.; Aziz, M.A.M.; Al-Suwailem, A.M. Culture-dependent bacteria in commercial fishes: Qualitative assessment and molecular identification using 16S rRNA gene sequencing. Saudi J. Biol. Sci. 2017, 24, 1105–1116. [Google Scholar] [CrossRef]
- Lingham, T.; Besong, S.; Ozbay, G.; Lee, J.-L. Antimicrobial activity of vinegar on bacterial species isolated from retail and local channel catfish (Ictalurus punctatus). J. Food. Process. Technol. 2012, 2, S11. [Google Scholar] [CrossRef]
- Wang, H.; Chu, W.; Ye, C.; Gaeta, B.; Tao, H.; Wang, M.; Qiu, Z. Chlorogenic acid attenuates virulence factors and pathogenicity of Pseudomonas aeruginosa by regulating quorum sensing. Appl. Microbiol. Biotechnol. 2019, 103, 903–915. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Lan, W.; Xie, J. Inhibitory effect of chlorogenic acid-grafted chitosan on seafood isolates Pseudomonas fluorescens and its biofilm. Lett. Appl. Microbiol. 2023, 76, ovad050. [Google Scholar] [CrossRef] [PubMed]
- Schuurmans, J.M.; Nuri Havali, A.S.; Koenders, B.B.; ter Kuile, B.H. Variations in MIC value caused by differences in experimental protocol. J. Microbiol. Met. 2009, 79, 44–47. [Google Scholar] [CrossRef] [PubMed]
- Witkowska, A.M.; Hickey, D.K.; Alonso-Gomez, M.; Wilkinson, M. Evaluation of antimicrobial activities of commercial herb and spice extract against selected food-borne bacteria. J. Food Res. 2013, 2, 37–54. [Google Scholar] [CrossRef]
- Gomes, F.; Dias, M.I.; Lima, Ă.; Barros, L.; Rodrigues, M.E.; Ferreira, I.C.F.R.; Henriques, M. Satureja montana L. and Origanum majorana L. decoctions: Antimicrobial activity, mode of action and phenolic characterization. Antibiotics 2020, 9, 294. [Google Scholar] [CrossRef]
- Blazek, A.D.; Paleo, B.J.; Weisleder, N. Plasma membrane repair: A central process for maintaining cellular homeostasis. Physiology 2015, 30, 438–448. [Google Scholar] [CrossRef]
- Abee, T.; Krockel, L.; Hill, C. Bacteriocins: Modes of actions and potentials in food preservation and control of food poisoning. Int. J. Food Microbiol. 1995, 28, 169–185. [Google Scholar] [CrossRef]
- Cai, R.; Miao, M.; Yue, T.; Zhang, Y.; Cui, L.; Wang, Z.; Yuan, Y. Antibacterial activity and mechanism of cinnamic acid and chlorogenic acid against Alicyclobacillus acidoterrestris vegetative cells in apple juice. Int. J. Food Sci. Technol. 2018, 54, 1697–1705. [Google Scholar] [CrossRef]
- Molva, C.; Baysal, A.H. Antibacterial activity of grape seed extract of Alicyclobacillus acidoterrestris DSM 3922 vegetative cells and spores in apple juice. LWT-Food Sci. Technol. 2015, 60, 238–245. [Google Scholar] [CrossRef]
- Tian, L.; Wu, M.; Guo, W.; Li, H.; Gai, Z.; Gong, G. Evaluation of the membrane damage mechanism of chlorogenic acid against Yersinia enterocolitica and Enterobacter sakazakii and its application in the preservation of raw pork and skim milk. Molecules 2021, 26, 6748. [Google Scholar] [CrossRef] [PubMed]
- Silber, N.; de Opitz, C.L.M.; Mayer, C.; Sass, P. Cell division protein FtsZ: From structure and mechanism to antibiotic target. Future Microbiol. 2020, 15, 699–831. [Google Scholar] [CrossRef]
- Mukherjee, A.; Lutkenhaus, J. Dynamic assembly of FtsZ regulated by GTP hydrolysis. EMBO J. 1998, 17, 462–469. [Google Scholar] [CrossRef] [PubMed]
- Zou, A.; Li, Y.; Ekanayake, S.B.; Dillon, J.A.R. An Escherichia coli expression model reveals the species-specific function of FtsA from Neisseria gonorrhoeae in cell division. FEMS Microbiol. Lett. 2017, 364, fnx078. [Google Scholar] [CrossRef]
- Rico, A.I.; Garcia-Ovalle, M.; Mingorance, J.; Vicente, M. Role of two essential domains of Escherichia coli FtsA in localization and progression of the division ring. Mol. Microbiol. 2004, 53, 1359–1371. [Google Scholar] [CrossRef]
- Pichoff, S.; Lutkenhaus, J. Identification of a region of FtsA required for interaction with FtsZ. Mol. Microbiol. 2007, 64, 1129–1138. [Google Scholar] [CrossRef]
- Herman, A.; Bochenek, J.; Herman, A.P. Effect of cinnamon and lavender oils on FtsZ gene expression in the Staphyloccocus aureus ATCC 29213. Appl. Biochem. Microbiol. 2013, 49, 481–484. [Google Scholar] [CrossRef]
- Mutlu, M.; Bingol, Z.; Uc, E.M.; Köksal, E.; Goren, A.C.; Alwasel, S.H.; Gulcin, I. Comprehensive metabolite profiling of cinnammon (Cinnamomum zeylanicum) leaf oil using LC-HR/MS, GC/MS, and GC-FID: Determination of antiglaucoma, antioxidant, anticholinergic, and antidiabetic profiles. Life 2023, 13, 136. [Google Scholar] [CrossRef]
- Li, G.; Wang, X.; Xu, Y.; Zhang, B.; Xia, X. Antimicrobial effect and mode of action of chlorogenic acid on Staphylococcus aureus. Eur. Food Res. Technol. 2013, 238, 589–596. [Google Scholar] [CrossRef]
- Duggirala, S.; Nankar, R.P.; Rajendran, S.; Doble, M. Phytochemicals as inhibitors of bacteria cel division protein FtsZ: Coumarins are promising candidates. Appl. Biochem. Biotechnol. 2014, 174, 283–296. [Google Scholar] [CrossRef] [PubMed]
- Lo Sciuto, A.; Fernández-Piñar, R.; Bertuccini, L.; Losi, F.; Superti, F.; Imperti, F. The periplasmic protein TolB as a potential drug target in Pseudomonas aeruginosa. PLoS ONE 2014, 9, e103784. [Google Scholar] [CrossRef] [PubMed]
- Lazzaroni, J.C.; Dubuisson, J.F.; Vianney, A. The Tol proteins in Escherichia coli and their involvement in the translocation of group A colicins. Biochimie 2002, 84, 391–397. [Google Scholar] [CrossRef] [PubMed]
- Hirakawa, H.; Suzue, K.; Tomita, H. Roles of the Tol/Pal system in bacterial pathogenesis and its application to antibacterial therapy. Vaccines 2022, 10, 422. [Google Scholar] [CrossRef]
- Li, Q.; Li, Z.; Fei, X.; Tian, Y.; Zhou, G.; Hu, Y.; Wang, S.; Shi, H. The role of TolA, TolB, and TolR in cell morphology, OMVs production, and virulence of Salmonella choleraesuis. AMB Express 2022, 12, 5. [Google Scholar] [CrossRef]
- Dubuisson, J.F.; Vianney, A.; Hugouvieux-Cotte-Pattat, N.; Lazzaroni, J.C. Tol-Pal proteins are critical cell envelope components of Erwinia chrysanthemi affecting cell morphology and virulence. Microbiologyresearch 2005, 151, 3337–3347. [Google Scholar] [CrossRef]
- Gyawali, R.; Ibrahim, S.A. Natural products as antimicrobial agents. Food Cont. 2014, 46, 412–429. [Google Scholar] [CrossRef]
- Adekoya, O.A.; Sylte, I. The thermolysin family (M4) of enzymes: Therapeutic and biotechnological potential. Chem. Biol. Drug Res. 2009, 73, 7–16. [Google Scholar] [CrossRef]
- Tian, L.; Liao, S.; Gou, W.; Wang, X.; Wu, M.; Xue, Z.; Yang, S.; Fu, J.; Liu, Z.; Gong, G. Evaluation of the membrane damage mechanism of chlorogenic acid against Bacillus cereus and Micrococcus luteus a simulation study on antibacterial growth in food. J. Food Saf. 2022, 42, e13006. [Google Scholar] [CrossRef]
- Yilmaz, H.; Subasi, B.G.; Celebioglu, H.U.; Ozdal, T.; Capanoglu, E. Chemistry of protein-phenolic interactions toward the microbiota and microbial infections. Front. Nutr. 2022, 9, 914118. [Google Scholar] [CrossRef]
- Karunanidhi, A.; Thomas, R.; von Belkum, A.; Neela, V. In vitro antibacterial and antibiofilm activities of chlorogenic acid against clinical isolates of Stenotrophomonas maltophilia including the trimethoprim/sulfamethoxazole resistant strain. Biomed. Res. Int. 2013, 2013, 392058. [Google Scholar] [CrossRef] [PubMed]
- Lou, Z.; Wang, H.; Zhu, S.; Ma, C.; Wang, Z. Antibacterial activity and mechanism of action of chlorogenic acid. J. Food Sci. 2011, 76, M398–M403. [Google Scholar] [CrossRef] [PubMed]
- Muthuswamy, S.; Vasantha-Rupasinghe, H.P. Fruit phenolics as natural antimicrobial agents: Selective antimicrobial activity of catechin, chlorogenic acid and phloridzin. J. Food Agri. Environ. 2007, 5, 81–85. [Google Scholar]
- Munekata, P.E.S.; Pateiro, M.; Domínguez, R.; Nieto, G.; Kumar, M.; Dhama, K.; Lorenzo, J.M. Bioactive compounds from fruits as preservatives. Foods 2023, 12, 343. [Google Scholar] [CrossRef]
- Arulkumar, A.; Paramasivam, S.; Miranda, J.M. Combined effect of icing medium and red alga Gracilaria verrucosa on shelf life extension of indian mackerel (Rastrelliger kanagurta). Food Bioprocess Technol. 2018, 11, 1911–1922. [Google Scholar] [CrossRef]
- Commission Regulation (EC) No 1022/2008 of 17 October 2008. Amending regulation (EC) No 2074/2005 as regards the total volatile basic nitrogen (TVB-N) limits. Off. J. EU 2008, I.277, 18–20. [Google Scholar]
- Ozyurt, G.; Kuley, E.; Balikei, E.; Kacar, C.; Gokdogan, S.; Etyemez, M.; Ozogul, F. Effect of the icing with rosemary extract on the oxidative stability and biogenic amine formation in sardine (Sardinella aurita) during chilled storage. Food Bioproc. Technol. 2012, 5, 2777–2786. [Google Scholar] [CrossRef]
- Viji, P.; Binsi, P.K.; Visnuvinayagam, S.; Bindu, J.; Ravishankar, C.N.; Srinivasa Gopal, T.K. Efficacy of mint (Mentha arvensis) leaf and citrus (Citrus aurantium) peel extract as natural preservatives for shelf life extension of chill stored Indian mackerel. J. Food Sci. Technol. 2015, 52, 6278–6289. [Google Scholar] [CrossRef]
- Arulkumar, A.; Swain, B.; Paramasivam, S. Shelf life extension of sardines (Sardinella albella) using betel leaf (Piper betle) incorporated ice. Food Bioproc. Technol. 2020, 13, 1255–1260. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence (5′→3′) | Amplified Region |
|---|---|---|
| 16S_F | GGAGACTGCCGGTGACAAAC | 16S rRNA gene (universal primers) |
| 16S_R | TGTAGCCCAGGCCGTAAGG | |
| FTSZ_F | GTAGGCCAGACGATTCAG | ftsZ gene |
| FTSZ_R | GGGCTTCACGATCTTCTT | |
| FTSA_F | CAAGTCGACGGACAGAAAA | ftsA gene |
| FTSA_R | CTGCCCACGCCAATAATA | |
| FTSN_F | GTACTGGTGGTGTTTGTC | ftsN gene |
| FTSN_R | TTCTTCTGGCTTAGGTGG | |
| TOLB_F | GTTCGCATTGAAATTACCC | tolB gene |
| TOLB_R | TACCACCCACATCTTCAG | |
| M4_F | TCCCGCCCTACATGCTTA | M4 gene |
| M4_R | CTTTGGCTGTCACGATCTT |
| Time of Incubation (h) | Log2 (Relative Quantity) | ||||
|---|---|---|---|---|---|
| ftsZ | ftsA | ftsN | tolB | M4 | |
| 24 | −2.5 | −1.5 | −2.0 | −1.5 | −1.5 |
| 72 | −2.5 | −1.0 | −2.5 | −1.5 | −1.5 |
| Storage Time (h) | Growth Inhibition (%) | |
|---|---|---|
| Salmon Fillets with Chlorogenic Acid (p = 0.105; F = 2.972) | Salmon Fillets with Benzoic Acid (p = 0.001; F = 8.163) | |
| 1 | 85 a ± 1.5 | 95 a ± 1.0 |
| 24 | 85 a ± 1.5 | 95 a ± 2.0 |
| 48 | 90 a ± 2.0 | 95 a ± 1.0 |
| 72 | 90 a ± 1.5 | 95 a ± 1.5 |
| 120 | 80 a ± 2.5 | 85 a b ± 2.0 |
| Storage Time (h) | TVB-N | TMA-N | ||||
|---|---|---|---|---|---|---|
| Salmon Fillets with Chlorogenic Acid (p = 0.0041; F = 8.225) | Salmon Fillets with Benzoic Acid (p < 0.001; F = 17.25) | Control (p = 0.0223; F = 5.725) | Salmon Fillets with Chlorogenic Acid (p < 0.0001; F = 33.52) | Salmon Fillets with Benzoic Acid (p < 0.001; F = 16.99) | Control (p < 0.0001; F = 15.39) | |
| 1 | 10 a ± 1.5 | 7.5 a ± 3.5 | 30 a ± 1.0 | 2.5 a ± 1.0 | 2.0 a ± 1.0 | 2.5 a ± 1.0 |
| 24 | 20 a ± 3.5 | 18.0 a ± 4.5 | 37 a ± 2.0 | 3.0 a ± 1.5 | 2.0 a ± 1.0 | 5.0 a ± 1.0 |
| 48 | 22 b,d ± 2.0 | 18.5 b,d ± 1.5 | 38 b,d ± 1.5 | 16.5 b,d ± 2.0 | 10.0 b,d ± 3.0 | 28.5 b,d ± 5.0 |
| 72 | 25 c,d ± 1.5 | 22 c,d ± 2.5 | 40 c,d ± 2.5 | 20.5 c,e ± 2.0 | 15.5 c,e ± 2.0 | 39.0 c,d ± 5.5 |
| 120 | 28 c,e ± 0.5 | 25.5 c,e ± 2.5 | 45 d,e ± 3.5 | 25.0 c,e ± 3.5 | 20.0 c,e ± 2.0 | 45.0 c,d ± 6.5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Myszka, K.; Tomaś, N.; Juzwa, W.; Wolko, Ł. Chlorogenic Acid Inhibits Rahnella aquatilis KM25 Growth and Proteolytic Activity in Fish-Based Products. Microorganisms 2023, 11, 1367. https://doi.org/10.3390/microorganisms11061367
Myszka K, Tomaś N, Juzwa W, Wolko Ł. Chlorogenic Acid Inhibits Rahnella aquatilis KM25 Growth and Proteolytic Activity in Fish-Based Products. Microorganisms. 2023; 11(6):1367. https://doi.org/10.3390/microorganisms11061367
Chicago/Turabian StyleMyszka, Kamila, Natalia Tomaś, Wojciech Juzwa, and Łukasz Wolko. 2023. "Chlorogenic Acid Inhibits Rahnella aquatilis KM25 Growth and Proteolytic Activity in Fish-Based Products" Microorganisms 11, no. 6: 1367. https://doi.org/10.3390/microorganisms11061367