
Oral Administration of Lactobacillus sakei CVL-001 Improves Recovery from Dextran Sulfate Sodium-Induced Colitis in Mice by Microbiota Modulation
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation and Identification of L. sakei CVL-001
2.2. Animals
2.3. In Vivo Experiments
2.4. Histological Evaluation
2.5. Real-Time PCR (qPCR)
2.6. Sequencing of Bacterial DNA Extracted from Mouse Feces
2.7. Operational Taxonomic Unit (OTU) Analysis for MiSeq
2.8. Statistical Analysis
3. Results
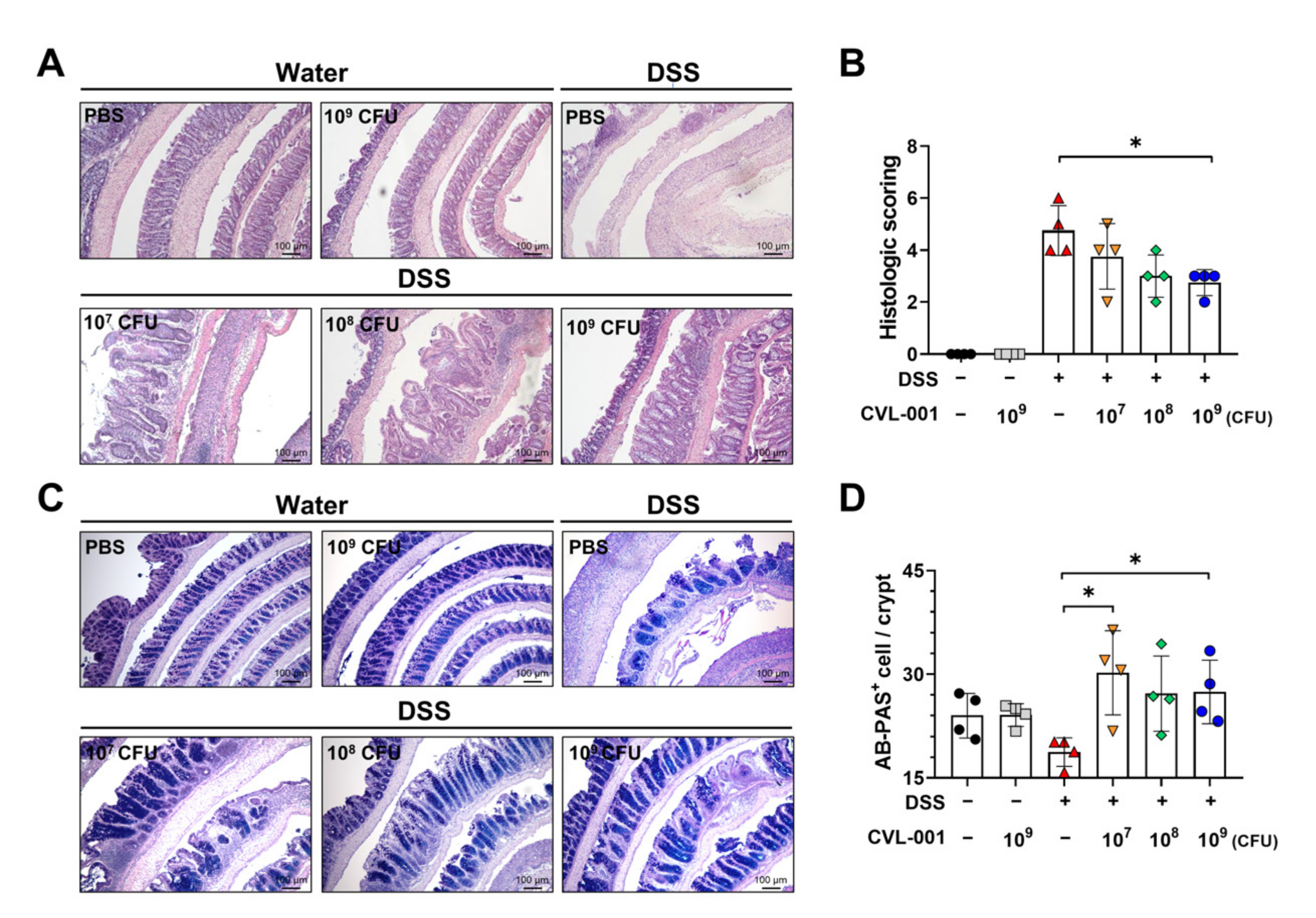
3.1. Oral Administration of L. sakei CVL-001 Ameliorates DSS-Induced Colitis
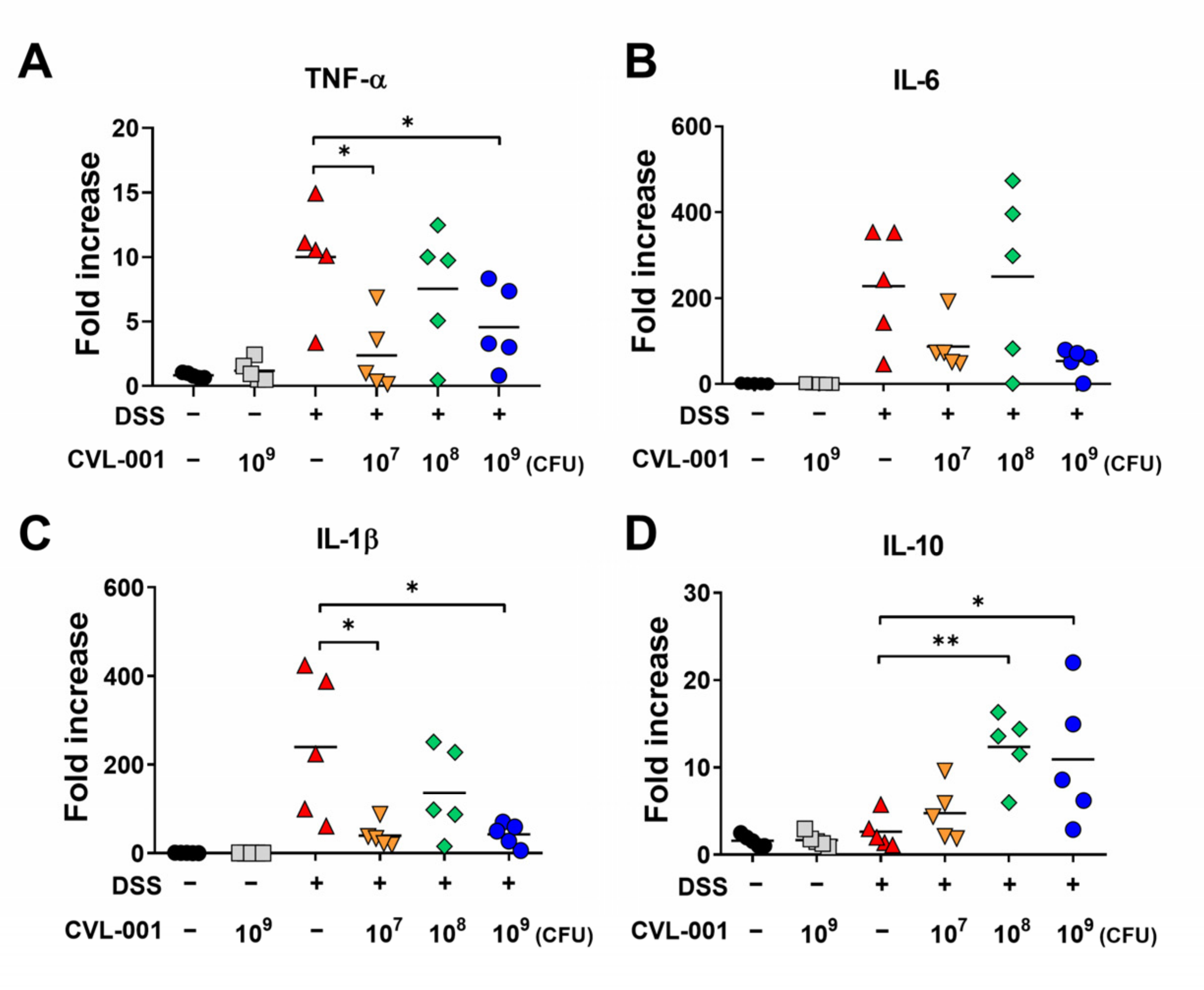
3.2. Anti-Inflammatory Effect of L. sakei CVL-001 against DSS-Induced Colitis
3.3. L. sakei CVL-001 Improves Intestinal Mucosal Barrier Function
3.4. Effect of Co-Housing on DSS-Induced Colitis in Mice Receiving L. sakei CVL-001
3.5. Administration of L. sakei CVL-001 Leads to Compositional Change of Gut Microbiota
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Mak, W.Y.; Zhao, M.; Ng, S.C.; Burisch, J. The epidemiology of inflammatory bowel disease: East meets west. J. Gastroenterol. Hepatol. 2020, 35, 380–389. [Google Scholar] [CrossRef]
- Baumgart, D.C.; Carding, S.R. Inflammatory bowel disease: Cause and immunobiology. Lancet 2007, 369, 1627–1640. [Google Scholar] [CrossRef] [PubMed]
- Irving, P.; Gearry, R.B.; Sparrow, M.P.; Gibson, P.R. Appropriate use of corticosteroids in Crohn’s disease. Aliment. Pharmacol. Ther. 2007, 26, 313–329. [Google Scholar] [CrossRef]
- Iborra, M.; Beltrán, B.; Bastida, G.; Aguas, M.; Nos, P. Infliximab and adalimumab-induced psoriasis in Crohn’s disease: A paradoxical side effect. J. Crohn’s Colitis 2011, 5, 157–161. [Google Scholar] [CrossRef]
- Hou, K.; Wu, Z.-X.; Chen, X.-Y.; Wang, J.-Q.; Zhang, D.; Xiao, C.; Zhu, D.; Koya, J.B.; Wei, L.; Li, J. Microbiota in health and diseases. Signal Transduct. Target. Ther. 2022, 7, 135. [Google Scholar] [CrossRef] [PubMed]
- DeGruttola, A.K.; Low, D.; Mizoguchi, A.; Mizoguchi, E. Current understanding of dysbiosis in disease in human and animal models. Inflamm. Bowel Dis. 2016, 22, 1137–1150. [Google Scholar] [CrossRef]
- Tamboli, C.P.; Neut, C.; Desreumaux, P.; Colombel, J.F. Dysbiosis in inflammatory bowel disease. Gut 2004, 53, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Pisani, A.; Rausch, P.; Bang, C.; Ellul, S.; Tabone, T.; Marantidis Cordina, C.; Zahra, G.; Franke, A.; Ellul, P. Dysbiosis in the gut microbiota in patients with inflammatory bowel disease during remission. Microbiol. Spectr. 2022, 10, e00616–e00622. [Google Scholar] [CrossRef] [PubMed]
- Stojanov, S.; Berlec, A.; Štrukelj, B. The influence of probiotics on the firmicutes/bacteroidetes ratio in the treatment of obesity and inflammatory bowel disease. Microorganisms 2020, 8, 1715. [Google Scholar] [CrossRef]
- Palmela, C.; Chevarin, C.; Xu, Z.; Torres, J.; Sevrin, G.; Hirten, R.; Barnich, N.; Ng, S.C.; Colombel, J.-F. Adherent-invasive Escherichia coli in inflammatory bowel disease. Gut 2018, 67, 574–587. [Google Scholar] [CrossRef]
- Salminen, S.; Ouwehand, A.; Isolauri, E. Clinical applications of probiotic bacteria. Int. Dairy J. 1998, 8, 563–572. [Google Scholar] [CrossRef]
- Liu, Y.; Alookaran, J.J.; Rhoads, J.M. Probiotics in autoimmune and inflammatory disorders. Nutrients 2018, 10, 1537. [Google Scholar] [CrossRef] [PubMed]
- Castellazzi, A.M.; Valsecchi, C.; Caimmi, S.; Licari, A.; Marseglia, A.; Leoni, M.C.; Caimmi, D.; Miraglia del Giudice, M.; Leonardi, S.; La Rosa, M. Probiotics and food allergy. Ital. J. Pediatr. 2013, 39, 47. [Google Scholar] [CrossRef]
- Bai, A.; Ouyang, Q. Probiotics and inflammatory bowel diseases. Postgrad. Med. J. 2006, 82, 376–382. [Google Scholar] [CrossRef] [PubMed]
- Cristofori, F.; Dargenio, V.N.; Dargenio, C.; Miniello, V.L.; Barone, M.; Francavilla, R. Anti-inflammatory and immunomodulatory effects of probiotics in gut inflammation: A door to the body. Front. Immunol. 2021, 12, 578386. [Google Scholar] [CrossRef] [PubMed]
- Wirtz, S.; Neufert, C.; Weigmann, B.; Neurath, M.F. Chemically induced mouse models of intestinal inflammation. Nat. Protoc. 2007, 2, 541–546. [Google Scholar] [CrossRef] [PubMed]
- Hendrickson, B.A.; Gokhale, R.; Cho, J.H. Clinical aspects and pathophysiology of inflammatory bowel disease. Clin. Microbiol. Rev. 2002, 15, 79–94. [Google Scholar] [CrossRef]
- Qin, D.; Ma, Y.; Wang, Y.; Hou, X.; Yu, L. Contribution of Lactobacilli on Intestinal Mucosal Barrier and Diseases: Perspectives and Challenges of Lactobacillus casei. Life 2022, 12, 1910. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Ji, H.; Wang, S.; Liu, H.; Zhang, W.; Zhang, D.; Wang, Y. Probiotic Lactobacillus plantarum promotes intestinal barrier function by strengthening the epithelium and modulating gut microbiota. Front. Microbiol. 2018, 9, 1953. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, B.; Delgado, S.; Blanco-Míguez, A.; Lourenço, A.; Gueimonde, M.; Margolles, A. Probiotics, gut microbiota, and their influence on host health and disease. Mol. Nutr. Food Res. 2017, 61, 1600240. [Google Scholar] [CrossRef]
- Do Carmo, F.L.; Rabah, H.; De Oliveira Carvalho, R.D.; Gaucher, F.; Cordeiro, B.F.; da Silva, S.H.; Le Loir, Y.; Azevedo, V.; Jan, G. Extractable bacterial surface proteins in probiotic–host interaction. Front. Microbiol. 2018, 9, 645. [Google Scholar] [CrossRef] [PubMed]
- Ren, C.; Cheng, L.; Sun, Y.; Zhang, Q.; de Haan, B.J.; Zhang, H.; Faas, M.M.; de Vos, P. Lactic acid bacteria secrete toll like receptor 2 stimulating and macrophage immunomodulating bioactive factors. J. Funct. Foods 2020, 66, 103783. [Google Scholar] [CrossRef]
- Yu, Q.; Yuan, L.; Deng, J.; Yang, Q. Lactobacillus protects the integrity of intestinal epithelial barrier damaged by pathogenic bacteria. Front. Cell. Infect. Microbiol. 2015, 5, 26. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, S.K. Bacteriocin-producing probiotic lactic acid bacteria in controlling dysbiosis of the gut microbiota. Front. Cell. Infect. Microbiol. 2022, 12, 415. [Google Scholar]
- Vemuri, R.; Gundamaraju, R.; Eri, R. Role of lactic acid probiotic bacteria in IBD. Curr. Pharm. Des. 2017, 23, 2352–2355. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Zhou, C.; Huang, J.; Kuai, X.; Shao, X. The potential therapeutic role of Lactobacillus reuteri for treatment of inflammatory bowel disease. Am. J. Transl. Res. 2020, 12, 1569. [Google Scholar]
- Li, M.-C.; He, S.-H. IL-10 and its related cytokines for treatment of inflammatory bowel disease. World J. Gastroenterol. 2004, 10, 620. [Google Scholar] [CrossRef] [PubMed]
- Jang, S.E.; Min, S.W. Lactobacillus sakei S1 Improves Colitis Induced by 2,4,6-Trinitrobenzene Sulfonic Acid by the Inhibition of NF-κB Signaling in Mice. J. Microbiol. Biotechnol. 2020, 30, 71–78. [Google Scholar] [CrossRef] [PubMed]
- Chapot-Chartier, M.-P.; Kulakauskas, S. Cell wall structure and function in lactic acid bacteria. Microb. Cell Factories 2014, 13, S9. [Google Scholar] [CrossRef]
- Menard, S.; Candalh, C.; Bambou, J.; Terpend, K.; Cerf-Bensussan, N.; Heyman, M. Lactic acid bacteria secrete metabolites retaining anti-inflammatory properties after intestinal transport. Gut 2004, 53, 821–828. [Google Scholar] [CrossRef] [PubMed]
- Hernández-González, J.C.; Martínez-Tapia, A.; Lazcano-Hernández, G.; García-Pérez, B.E.; Castrejón-Jiménez, N.S. Bacteriocins from lactic acid bacteria. A powerful alternative as antimicrobials, probiotics, and immunomodulators in veterinary medicine. Animals 2021, 11, 979. [Google Scholar] [CrossRef] [PubMed]
- Seo, H.; Seong, H.; Kim, G.Y.; Jo, Y.M.; Cheon, S.W.; Song, Y.; Ryu, B.H.; Kang, H.; Han, N.S. Development of Anti-inflammatory Probiotic Limosilactobacillus reuteri EFEL6901 as Kimchi Starter: In vitro and In vivo Evidence. Front. Microbiol. 2021, 12, 760476. [Google Scholar] [CrossRef] [PubMed]
- Seo, S.; Shin, J.-S.; Lee, W.-S.; Rhee, Y.K.; Cho, C.-W.; Hong, H.-D.; Lee, K.-T. Anti-colitis effect of Lactobacillus sakei K040706 via suppression of inflammatory responses in the dextran sulfate sodium-induced colitis mice model. J. Funct. Foods 2017, 29, 256–268. [Google Scholar] [CrossRef]
- Rather, I.A.; Bajpai, V.K.; Ching, L.L.; Majumder, R.; Nam, G.-J.; Indugu, N.; Singh, P.; Kumar, S.; Hajrah, N.H.; Sabir, J.S. Effect of a bioactive product SEL001 from Lactobacillus sakei probio65 on gut microbiota and its anti-colitis effects in a TNBS-induced colitis mouse model. Saudi J. Biol. Sci. 2020, 27, 261–270. [Google Scholar] [CrossRef] [PubMed]
- Chelakkot, C.; Ghim, J.; Ryu, S.H. Mechanisms regulating intestinal barrier integrity and its pathological implications. Exp. Mol. Med. 2018, 50, 1–9. [Google Scholar] [CrossRef]
- Okamoto, R.; Watanabe, M. Role of epithelial cells in the pathogenesis and treatment of inflammatory bowel disease. J. Gastroenterol. 2016, 51, 11–21. [Google Scholar] [CrossRef]
- Chen, J.; Zhang, S.; Feng, X.; Wu, Z.; Dubois, W.; Thovarai, V.; Ahluwalia, S.; Gao, S.; Chen, J.; Peat, T. Conventional co-housing modulates murine gut microbiota and hematopoietic gene expression. Int. J. Mol. Sci. 2020, 21, 6143. [Google Scholar] [CrossRef]
- Liu, C.; Huang, S.; Wu, Z.; Li, T.; Li, N.; Zhang, B.; Han, D.; Wang, S.; Zhao, J.; Wang, J. Cohousing-mediated microbiota transfer from milk bioactive components-dosed mice ameliorate colitis by remodeling colonic mucus barrier and lamina propria macrophages. Gut Microbes 2021, 13, 1903826. [Google Scholar] [CrossRef]
- Spalinger, M.R.; Schwarzfischer, M.; Hering, L.; Shawki, A.; Sayoc, A.; Santos, A.; Gottier, C.; Lang, S.; Bäbler, K.; Geirnaert, A. Loss of PTPN22 abrogates the beneficial effect of cohousing-mediated fecal microbiota transfer in murine colitis. Mucosal Immunol. 2019, 12, 1336–1347. [Google Scholar] [CrossRef] [PubMed]
- Kwon, M.-S.; Lim, S.K.; Jang, J.-Y.; Lee, J.; Park, H.K.; Kim, N.; Yun, M.; Shin, M.-Y.; Jo, H.E.; Oh, Y.J. Lactobacillus sakei WIKIM30 ameliorates atopic dermatitis-like skin lesions by inducing regulatory T cells and altering gut microbiota structure in mice. Front. Immunol. 2018, 9, 1905. [Google Scholar] [CrossRef]
- Magne, F.; Gotteland, M.; Gauthier, L.; Zazueta, A.; Pesoa, S.; Navarrete, P.; Balamurugan, R. The firmicutes/bacteroidetes ratio: A relevant marker of gut dysbiosis in obese patients? Nutrients 2020, 12, 1474. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, K.; Nishida, A.; Fujimoto, T.; Fujii, M.; Shioya, M.; Imaeda, H.; Inatomi, O.; Bamba, S.; Andoh, A.; Sugimoto, M. Reduced abundance of butyrate-producing bacteria species in the fecal microbial community in Crohn’s disease. Digestion 2016, 93, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Won, S.-M.; Chen, S.; Lee, S.Y.; Lee, K.E.; Park, K.W.; Yoon, J.-H. Lactobacillus sakei ADM14 induces anti-obesity effects and changes in gut microbiome in high-fat diet-induced obese mice. Nutrients 2020, 12, 3703. [Google Scholar] [CrossRef] [PubMed]
- Friswell, M.; Campbell, B.; Rhodes, J. The role of bacteria in the pathogenesis of inflammatory bowel disease. Gut Liver 2010, 4, 295. [Google Scholar] [CrossRef] [PubMed]
- Shin, N.-R.; Whon, T.W.; Bae, J.-W. Proteobacteria: Microbial signature of dysbiosis in gut microbiota. Trends Biotechnol. 2015, 33, 496–503. [Google Scholar] [CrossRef]
- Chen, C.-S.; Sullivan, S.; Anderson, T.; Tan, A.C.; Alex, P.J.; Brant, S.R.; Cuffari, C.; Bayless, T.M.; Talor, M.V.; Burek, C.L. Identification of novel serological biomarkers for inflammatory bowel disease using Escherichia coli proteome chip. Mol. Cell. Proteom. 2009, 8, 1765–1776. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Hoedt, E.C.; Liu, Q.; Berendsen, E.; Teh, J.J.; Hamilton, A.; O’Brien, A.W.; Ching, J.Y.; Wei, H.; Yang, K. Elucidation of Proteus mirabilis as a key bacterium in Crohn’s disease inflammation. Gastroenterology 2021, 160, 317.e311–330.e311. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Score | Weight Loss | Stool Consistency | Blood |
|---|---|---|---|
| 0 | None | Normal | Normal |
| 1 | 1–5% | Soft but still formed | Slight hemoccult |
| 2 | 6–10% | Soft | Positive hemoccult |
| 3 | 11–18% | Wet, very soft | Blood traces in stool |
| 4 | >18% | Watery diarrhea | Gross rectal bleeding |
| Score | Epithelial Integrity | Lamina Propria Inflammatory Cell Infiltration |
|---|---|---|
| 0 | None | Infrequent |
| 1 | Focal epithelial damage | Increased, some neutrophils |
| 2 | Mucosal erosions and ulceration | Submucosal presence of inflammatory cell clusters |
| 3 | Extensive epithelial damage | Transmural cell infiltration |
| Genes | Sequence 5′-3′ |
|---|---|
| IL-6 | F: GCT ACC AAA CTG GAT ATA ATC AGG A |
| R: CCA GGT AGC TAT GGT ACT CCA GAA | |
| TNF-α | F: TCT GTC TAC TGA ACT TCG GGG TGA |
| R: TTG TCT TTG AGA TCC ATG CCG TT | |
| IL-1β | F: CAA CCA ACA AGT GAT ATT CTC CAT G |
| R: ATC CAC ACT CTC CAG CTG CA | |
| IL-10 | F: TAA CTG CAC CCA CTT CCC AG |
| R: AGG CTT GGC AAC CCA AGT AA | |
| GAPDH | F: CGG AGT CAA CGG ATT TGG TCG TAT |
| R: AGC CTT CTC CAT GGT GGT GAA GAC | |
| E-cadherin | F: CAG GTC TCC TCA TGG CTT TGC |
| R: CTT CCG AAA AGA AGG CTG TCC | |
| Occludin | F: CAC ACT TGC TTG GGA CAG AG |
| R: TAG CCA TAG CCT CCA TAG CC | |
| Claudin-3 | F: AAC TGC GTA CAA GAC GAG ACG |
| R: ATC CCT GAT GAT GGT GTT GG | |
| ZO-1 | F: CCT AAG ACC TGT AAC CAT CT |
| R: CTG ATA GAT ATC TGG CTC CT | |
| MUC2 | F: TGC TGA CGA GTG GTT GGT GAA TG |
| R: TGA TGA GGT GGC AGA CAG GAG AC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, D.-Y.; Lee, T.-S.; Jung, D.-H.; Song, E.-J.; Jang, A.-R.; Park, J.-Y.; Ahn, J.-H.; Seo, I.-S.; Song, S.-J.; Kim, Y.-J.; et al. Oral Administration of Lactobacillus sakei CVL-001 Improves Recovery from Dextran Sulfate Sodium-Induced Colitis in Mice by Microbiota Modulation. Microorganisms 2023, 11, 1359. https://doi.org/10.3390/microorganisms11051359
Kim D-Y, Lee T-S, Jung D-H, Song E-J, Jang A-R, Park J-Y, Ahn J-H, Seo I-S, Song S-J, Kim Y-J, et al. Oral Administration of Lactobacillus sakei CVL-001 Improves Recovery from Dextran Sulfate Sodium-Induced Colitis in Mice by Microbiota Modulation. Microorganisms. 2023; 11(5):1359. https://doi.org/10.3390/microorganisms11051359
Chicago/Turabian StyleKim, Dong-Yeon, Tae-Sung Lee, Do-Hyeon Jung, Eun-Jung Song, Ah-Ra Jang, Ji-Yeon Park, Jae-Hun Ahn, In-Su Seo, Seung-Ju Song, Yeong-Jun Kim, and et al. 2023. "Oral Administration of Lactobacillus sakei CVL-001 Improves Recovery from Dextran Sulfate Sodium-Induced Colitis in Mice by Microbiota Modulation" Microorganisms 11, no. 5: 1359. https://doi.org/10.3390/microorganisms11051359