
Chitinolytic Enzymes of the Hyperparasite Fungus Aphanocladium album: Genome-Wide Survey and Characterization of A Selected Enzyme
 , , , , ,
, , , , ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strain and Growth Conditions
2.2. DNA Isolation and Sequencing
2.3. Genome Assembly Annotation
2.4. Analysis of Protein Sequences
2.5. Gene Cloning, Expression, and Purification of rChi1
2.6. Chitinase Activity Assay
3. Results and Discussion
3.1. A. album DNA Sequencing and Genome Assembly
3.2. Chitinases in A. album: Genome Analysis and Protein Structural Analysis
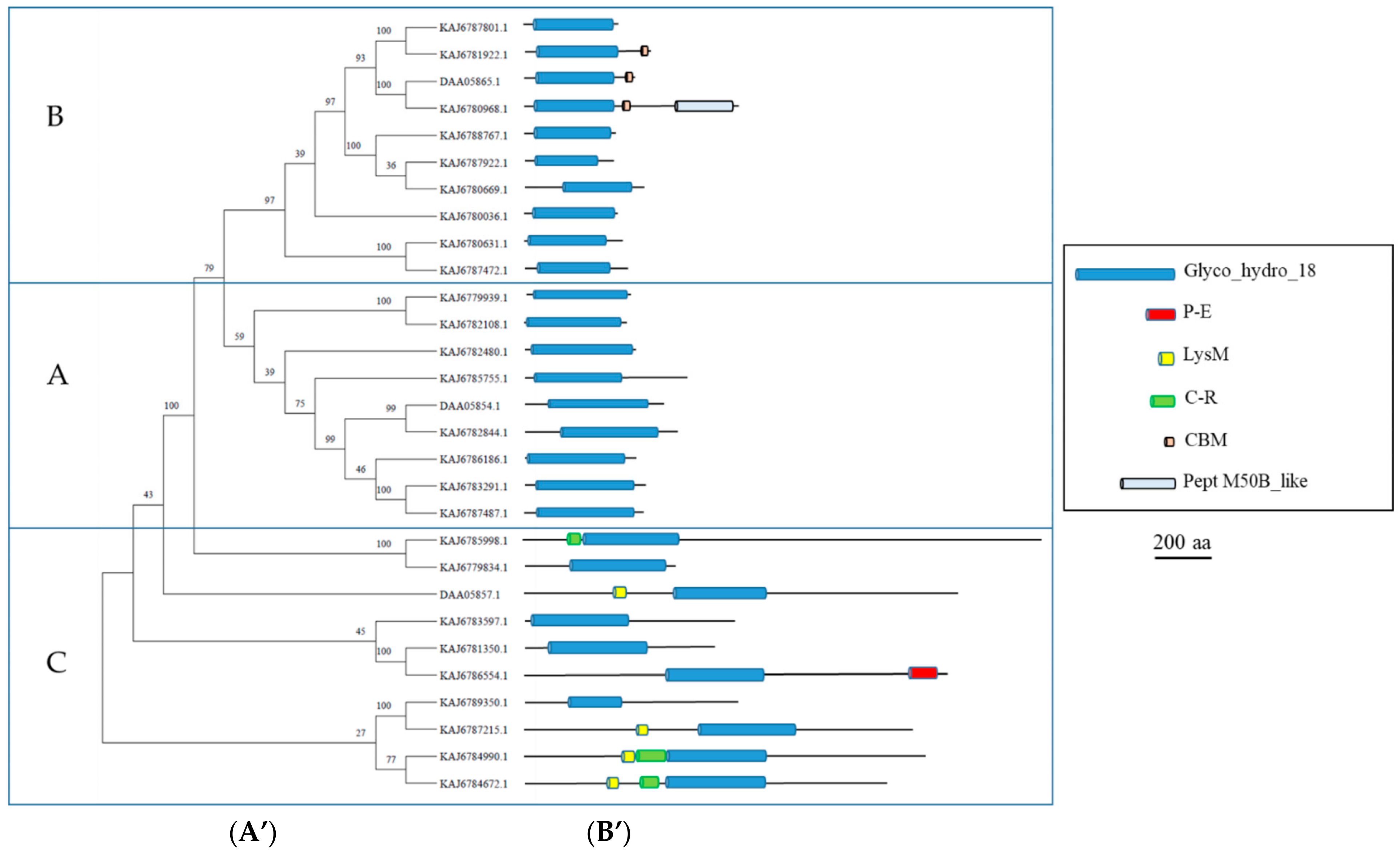
3.3. GH18 Enzymes
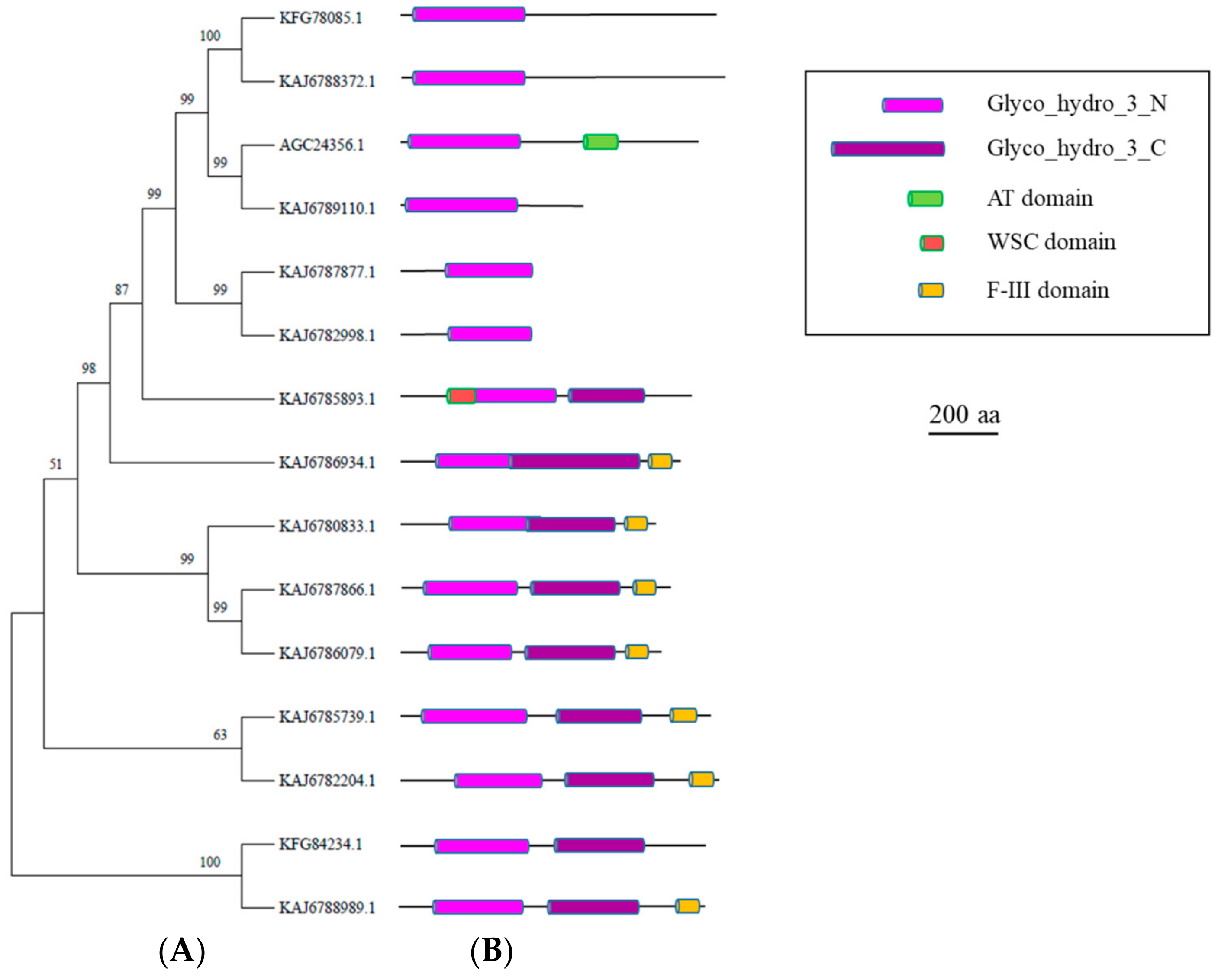
3.4. GH3 Enzymes
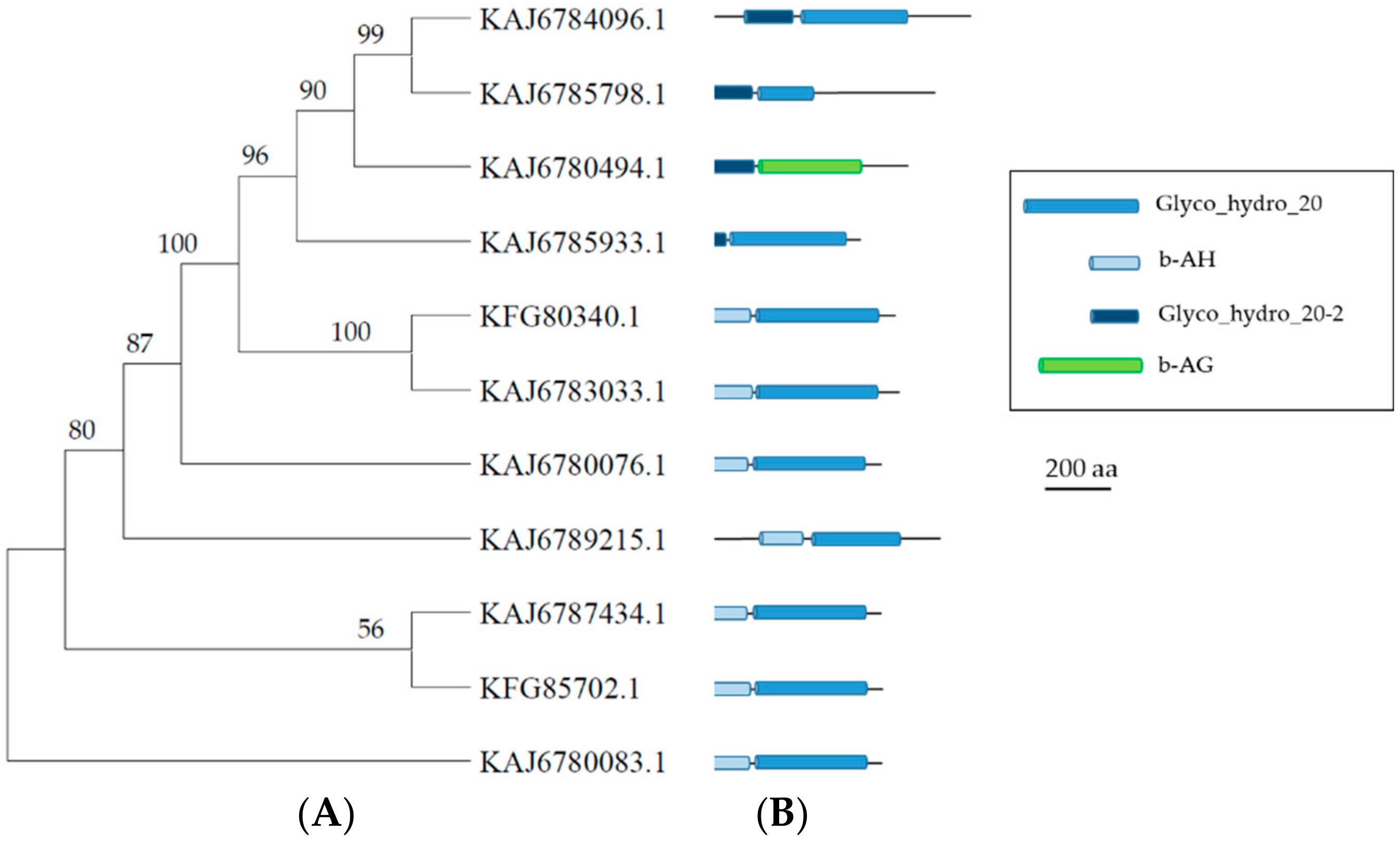
3.5. GH20 Enzymes
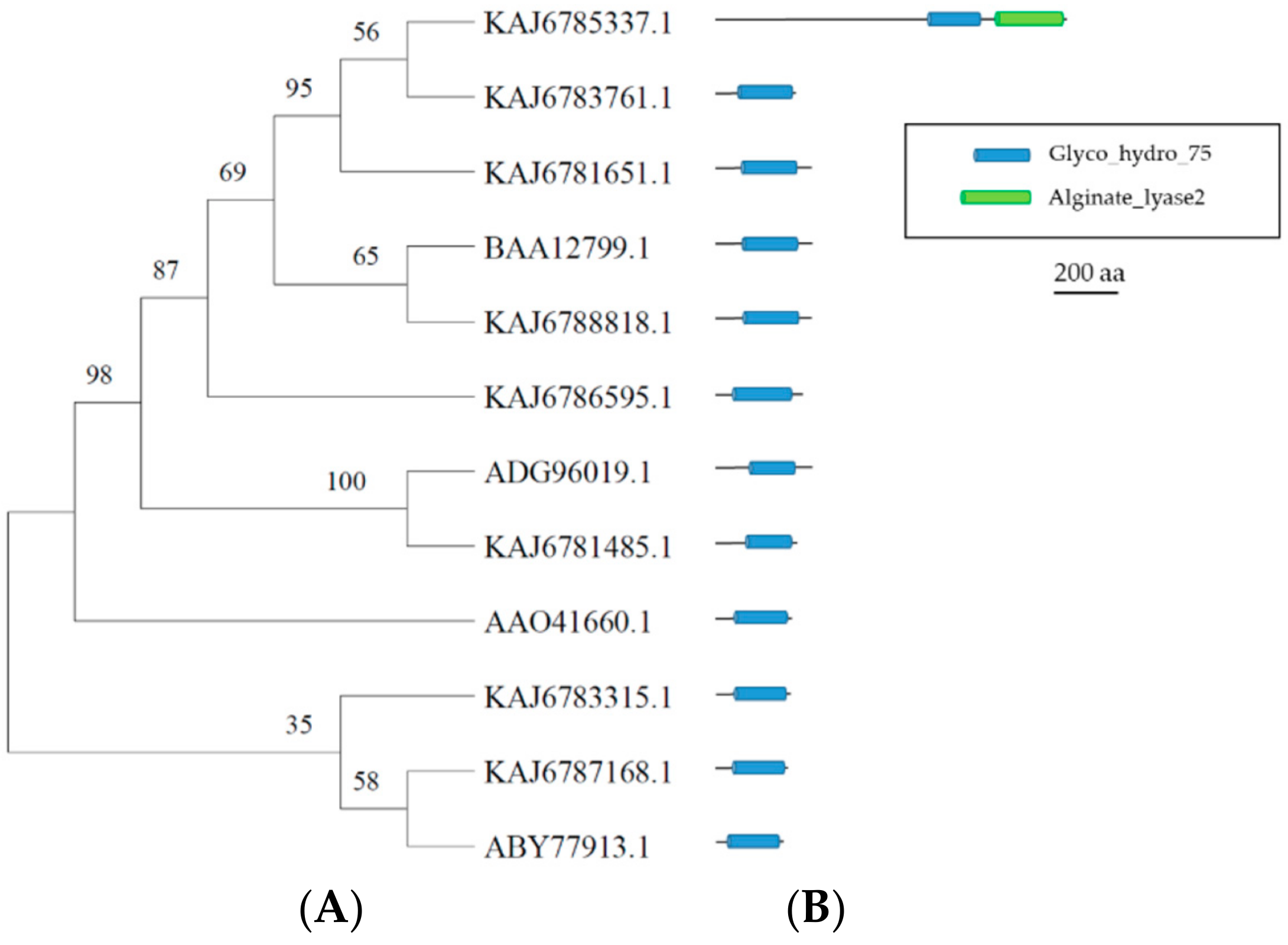
3.6. GH75 Enzymes
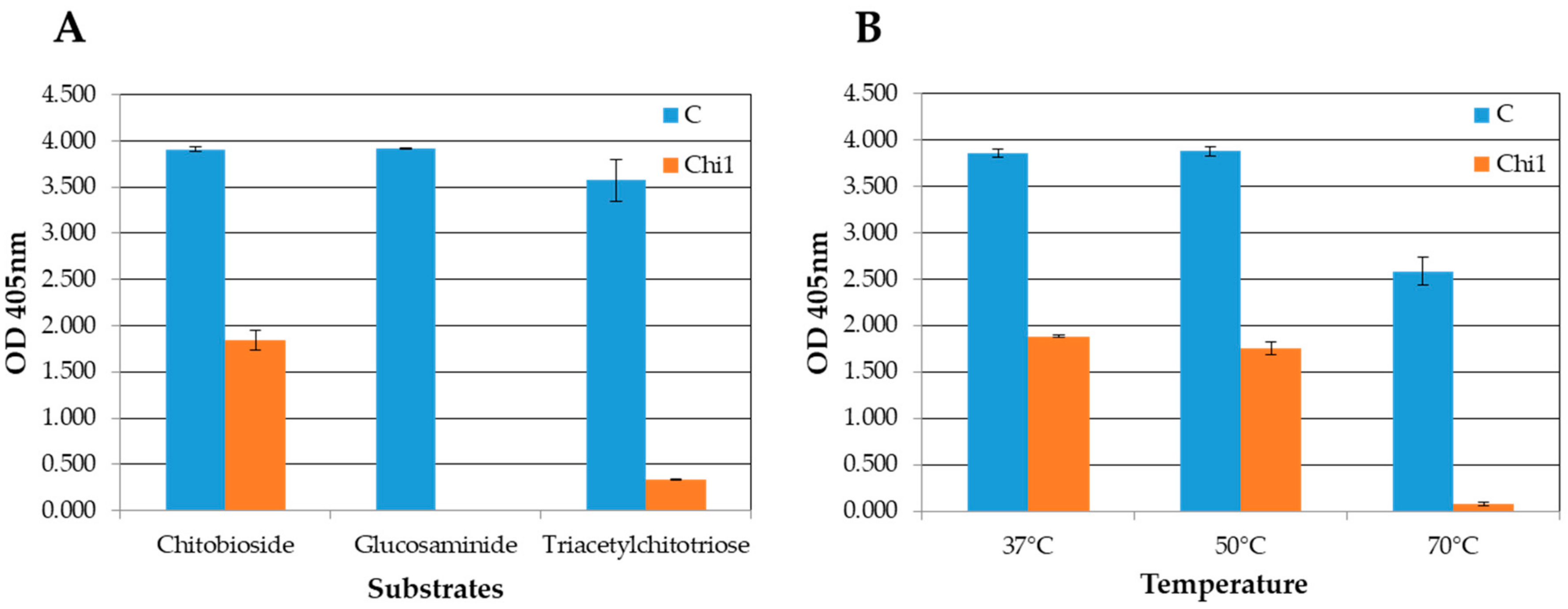
3.7. Expression and Activity Assays of the rChi1 Protein
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Biali, M.; Dinoor, A.; Eshed, N.; Kenneth, R. Aphanocladium album, a Fungus Inducing Teliospore Production in Rusts. Ann. Appl. Biol. 1972, 72, 37–42. [Google Scholar] [CrossRef]
- Blaiseau, P.L.; Kunz, C.; Grison, R.; Bertheau, Y.; Brygoo, Y. Cloning and Expression of a Chitinase Gene from the Hyperparasitic Fungus Aphanocladium album. Curr. Genet. 1992, 21, 61–66. [Google Scholar] [CrossRef] [PubMed]
- Kunz, C.; Sellam, O.; Bertheau, Y. Purification and Characterization of a Chitinase from the Hyperparasitic Fungus Aphanocladium album. Physiol. Mol. Plant Pathol. 1992, 40, 117–131. [Google Scholar] [CrossRef]
- Ciccarese, F.; Amenduni, M.; Schiavone, D.; Ambrico, A. Aphanocladium album a New Promising Biocontrol Agent against Oidiumlycopersici. In Proceedings of the 10th Congress of the Mediterranean Phytopathological Union, Montpellier, France, 1–5 June 1997; pp. 559–562. [Google Scholar]
- Ciccarese, F.; Longo, O.; Ambrica, A.; Schiavone, D. Aphanocladium album: Un promettente limitatore biologico dell’oidio del pomodoro e dello zucchino. La difesa delle colture in agricoltura biologica. Not. Mal. Piante 2001, 52, 69–71. [Google Scholar]
- Blaiseau, P.L.; Lafay, J.F. Primary Structure of a Chitinase-Encoding Gene (Chi1) from the Filamentous Fungus Aphanocladium album: Similarity to Bacterial Chitinases. Gene 1992, 120, 243–248. [Google Scholar] [CrossRef]
- Leoni, C.; Piancone, E.; Sasanelli, N.; Bruno, G.L.; Manzari, C.; Pesole, G.; Ceci, L.R.; Volpicella, M. Plant Health and Rhizosphere Microbiome: Effects of the Bionematicide Aphanocladium album in Tomato Plants Infested by Meloidogyne javanica. Microorganisms 2020, 8, 1922. [Google Scholar] [CrossRef]
- D’Ambrosio, G.; Cariddi, C.; Mannerucci, F.; Bruno, G.L. In Vitro Screening of New Biological Limiters against Some of the Main Soil-Borne Phytopathogens. Sustainaibility 2022, 14, 2693. [Google Scholar] [CrossRef]
- Moussian, B. Chitin: Structure, Chemistry and Biology. In Targeting Chitin-Containing Organisms; Yang, Q., Fukamizo, T., Eds.; Advances in Experimental Medicine and Biology; Springer: Singapore, 2019; Volume 1142, pp. 5–18. ISBN 9789811373176. [Google Scholar]
- Naveed, M.; Phil, L.; Sohail, M.; Hasnat, M.; Baig, M.M.F.A.; Ihsan, A.U.; Shumzaid, M.; Kakar, M.U.; Mehmood Khan, T.; Akabar, M.; et al. Chitosan Oligosaccharide (COS): An Overview. Int. J. Biol. Macromol. 2019, 129, 827–843. [Google Scholar] [CrossRef]
- Poria, V.; Rana, A.; Kumari, A.; Grewal, J.; Pranaw, K.; Singh, S. Current Perspectives on Chitinolytic Enzymes and Their Agro-Industrial Applications. Biology 2021, 10, 1319. [Google Scholar] [CrossRef]
- Gomaa, E.Z. Microbial Chitinases: Properties, Enhancement and Potential Applications. Protoplasma 2021, 258, 695–710. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A Flexible Trimmer for Illumina Sequence Data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Zhang, J.; Kobert, K.; Flouri, T.; Stamatakis, A. PEAR: A Fast and Accurate Illumina Paired-End ReAd MergeR. Bioinformatics 2014, 30, 614–620. [Google Scholar] [CrossRef]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A New Genome Assembly Algorithm and Its Applications to Single-Cell Sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef]
- Stanke, M.; Morgenstern, B. AUGUSTUS: A web server for gene prediction in eukaryotes that allows user-defined constraints. Nucleic Acids Res. 2005, 33, W465–W467. [Google Scholar] [CrossRef]
- Mistry, J.; Finn, R. Pfam: A Domain-Centric Method for Analyzing Proteins and Proteomes. Methods Mol. Biol. 2007, 396, 43–58. [Google Scholar] [CrossRef]
- Drula, E.; Garron, M.-L.; Dogan, S.; Lombard, V.; Henrissat, B.; Terrapon, N. The Carbohydrate-Active Enzyme Database: Functions and Literature. Nucleic Acids Res. 2022, 50, D571–D577. [Google Scholar] [CrossRef]
- Sievers, F.; Wilm, A.; Dineen, D.; Gibson, T.J.; Karplus, K.; Li, W.; Lopez, R.; McWilliam, H.; Remmert, M.; Söding, J.; et al. Fast, Scalable Generation of High-quality Protein Multiple Sequence Alignments Using Clustal Omega. Mol. Syst. Biol. 2011, 7, 539. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Potter, S.C.; Luciani, A.; Eddy, S.R.; Park, Y.; Lopez, R.; Finn, R.D. HMMER Web Server: 2018 Update. Nucleic Acids Res. 2018, 46, W200–W204. [Google Scholar] [CrossRef]
- Kelley, L.A.; Mezulis, S.; Yates, C.M.; Wass, M.N.; Sternberg, M.J.E. The Phyre2 Web Portal for Protein Modeling, Prediction and Analysis. Nat. Protoc. 2015, 10, 845–858. [Google Scholar] [CrossRef]
- Volpicella, M.; Leoni, C.; Fanizza, I.; Placido, A.; Pastorello, E.A.; Ceci, L.R. Overview of Plant Chitinases Identified as Food Allergens. J. Agric. Food Chem. 2014, 62, 5734–5742. [Google Scholar] [CrossRef] [PubMed]
- Ramírez, G.M.; Avelizapa, L.I.R.; Avelizapa, N.G.R.; Camarillo, C.R. Colloidal Chitin Stained with Remazol Brilliant Blue R®, a Useful Substrate to Select Chitinolytic Microorganisms and to Evaluate Chitinases. J. Microbiol. Methods 2004, 56, 213–219. [Google Scholar] [CrossRef] [PubMed]
- Langdon, W.B. Performance of Genetic Programming Optimised Bowtie2 on Genome Comparison and Analytic Testing (GCAT) Benchmarks. BioData Min. 2015, 8, 1. [Google Scholar] [CrossRef] [PubMed]
- Manni, M.; Berkeley, M.R.; Seppey, M.; Zdobnov, E.M. BUSCO: Assessing Genomic Data Quality and Beyond. Curr. Protoc. 2021, 1, e323. [Google Scholar] [CrossRef] [PubMed]
- Zdobnov, E.M.; Tegenfeldt, F.; Kuznetsov, D.; Waterhouse, R.M.; Simão, F.A.; Ioannidis, P.; Seppey, M.; Loetscher, A.; Kriventseva, E.V. OrthoDB v9.1: Cataloging Evolutionary and Functional Annotations for Animal, Fungal, Plant, Archaeal, Bacterial and Viral Orthologs. Nucleic Acids Res. 2017, 45, D744–D749. [Google Scholar] [CrossRef] [PubMed]
- Seidl, V. Chitinases of Filamentous Fungi: A Large Group of Diverse Proteins with Multiple Physiological Functions. Fungal Biol. Rev. 2008, 22, 36–42. [Google Scholar] [CrossRef]
- Thadathil, N.; Velappan, S.P. Recent Developments in Chitosanase Research and Its Biotechnological Applications: A Review. Food Chem. 2014, 150, 392–399. [Google Scholar] [CrossRef]
- Seidl, V.; Huemer, B.; Seiboth, B.; Kubicek, C.P. A Complete Survey of Trichoderma Chitinases Reveals Three Distinct Subgroups of Family 18 Chitinases. FEBS J. 2005, 272, 5923–5939. [Google Scholar] [CrossRef]
- Magliani, W.; Conti, S.; Gerloni, M.; Bertolotti, D.; Polonelli, L. Yeast Killer Systems. Clin. Microbiol. Rev. 1997, 10, 369–400. [Google Scholar] [CrossRef]
- Tzelepis, G.; Karlsson, M. The Fungal Chitinases. In Encyclopedia of Mycology; Elsevier: Amsterdam, The Netherlands, 2021; pp. 23–31. ISBN 978-0-323-85180-0. [Google Scholar]
- Kamerewerd, J.; Zadra, I.; Kürnsteiner, H.; Kück, U. PcchiB1, Encoding a Class V Chitinase, Is Affected by PcVelA and PcLaeA, and Is Responsible for Cell Wall Integrity in Penicillium chrysogenum. Microbiology 2011, 157, 3036–3048. [Google Scholar] [CrossRef]
- Masuda, T.; Zhao, G.; Mikami, B. Crystal Structure of Class III Chitinase from Pomegranate Provides the Insight into Its Metal Storage Capacity. Biosci. Biotechnol. Biochem. 2015, 79, 45–50. [Google Scholar] [CrossRef]
- Slámová, K.; Bojarová, P.; Petrásková, L.; Kren, V. β-N-Acetylhexosaminidase: What’s in a Name…? Biotechnol. Adv. 2010, 28, 682–693. [Google Scholar] [CrossRef]
- Yang, S.; Song, S.; Yan, Q.; Fu, X.; Jiang, Z.; Yang, X. Biochemical Characterization of the First Fungal Glycoside Hydrolyase Family 3 β-N-Acetylglucosaminidase from Rhizomucor miehei. J. Agric. Food Chem. 2014, 62, 5181–5190. [Google Scholar] [CrossRef]
- De Oliveira, E.S.; Junges, Â.; Sbaraini, N.; Andreis, F.C.; Thompson, C.E.; Staats, C.C.; Schrank, A. Molecular Evolution and Transcriptional Profile of GH3 and GH20 β-N-Acetylglucosaminidases in the Entomopathogenic Fungus Metarhizium anisopliae. Genet. Mol. Biol. 2018, 41, 843–857. [Google Scholar] [CrossRef]
- Kojima, K.; Sunagawa, N.; Mikkelsen, N.E.; Hansson, H.; Karkehabadi, S.; Samejima, M.; Sandgren, M.; Igarashi, K. Comparison of Glycoside Hydrolase Family 3 β-Xylosidases from Basidiomycetes and Ascomycetes Reveals Evolutionarily Distinct Xylan Degradation Systems. J. Biol. Chem. 2022, 298, 101670. [Google Scholar] [CrossRef]
- Zhu, X.-F.; Tan, H.-Q.; Zhu, C.; Liao, L.; Zhang, X.-Q.; Wu, M. Cloning and Overexpression of a New Chitosanase Gene from Penicillium sp. D-1. AMB Express 2012, 2, 13. [Google Scholar] [CrossRef]
- Vasseur, V.; Arigoni, F.; Andersen, H.; Defago, G.; Bompeix, G.; Seng, J.-M. Isolation and Characterization of Aphanocladium album Chitinase-Overproducing Mutants. J. Gen. Microbiol. 1990, 136, 2561–2567. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CAZy Family | ID | Amino Acids | Activity |
|---|---|---|---|
| GH18 | KAJ6784672.1 | 1279 | Exochitinase |
| KAJ6786186.1 | 394 | “ | |
| KAJ6780036.1 | 330 | Endochitinase | |
| KAJ6780669.1 | 423 | “ | |
| KAJ6787801.1 | 336 | “ | |
| KAJ6780631.1 | 348 | “ | |
| KAJ6783291.1 | 428 | Exochitinase | |
| KAJ6787472.1 | 365 | Endochitinase | |
| KAJ6787487.1 | 423 | Exochitinase | |
| KAJ6781350.1 | 671 | “ | |
| KAJ6782844.1 | 540 | “ | |
| KAJ6788767.1 | 324 | Endochitinase | |
| KAJ6784990.1 | 1418 | Exochitinase | |
| KAJ6785755.1 | 574 | “ | |
| KAJ6779939.1 | 369 | “ | |
| KAJ6780968.1 | 757 | Endochitinase | |
| KAJ6787215.1 | 1373 | Exochitinase | |
| KAJ6786554.1 | 1495 | “ | |
| KAJ6781922.1 | 445 | Endochitinase | |
| KAJ6785998.1 | 1832 | Exochitinase | |
| KAJ6782480.1 | 392 | “ | |
| KAJ6782108.1 | 363 | “ | |
| KAJ6789350.1 | 754 | “ | |
| KAJ6779834.1 | 532 | “ | |
| KAJ6783597.1 | 742 | “ | |
| KAJ6787922.1 | 315 | Endochitinase | |
| GH3 | KAJ6786934.1 | 830 | β-glucosidase |
| KAJ6780833.1 | 760 | “ | |
| KAJ6788989.1 | 883 | “ | |
| KAJ6788372.1 | 932 | β-N-acetylhexosaminidase | |
| KAJ6782204.1 | 948 | β-glucosidase | |
| KAJ6785739.1 | 893 | “ | |
| KAJ6787866.1 | 802 | “ | |
| KAJ6785893.1 | 870 | β-xylosidase | |
| KAJ6787877.1 | 370 | β-N-acetylhexosaminidase | |
| KAJ6789110.1 | 539 | “ | |
| KAJ6782998.1 | 369 | “ | |
| KAJ6786079.1 | 777 | β-glucosidase | |
| GH20 | KAJ6780076.1 | 577 | β-N-acetylhexosaminidase |
| KAJ6780083.1 | 579 | “ | |
| KAJ6785933.1 | 514 | “ | |
| KAJ6783033.1 | 633 | “ | |
| KAJ6780494.1 | 661 | GH 84 hydrolase | |
| KAJ6787434.1 | 574 | β-N-acetylhexosaminidase | |
| KAJ6784096.1 | 848 | “ | |
| KAJ6785798.1 | 741 | “ | |
| KAJ6789215.1 | 751 | “ | |
| GH75 | KAJ6781651.1 | 299 | Chitosanase |
| KAJ6783315.1 | 232 | “ | |
| KAJ6785337.1 | 1093 | “ | |
| KAJ6781485.1 | 256 | “ | |
| KAJ6788818.1 | 300 | “ | |
| KAJ6783761.1 | 252 | “ | |
| KAJ6786595.1 | 273 | “ | |
| KAJ6787168.1 | 234 | “ |
| pH | T (°C) | rChi1 Specific Activity (U/μg) * |
|---|---|---|
| 7 | 37 | 0.51 ± 0.11 |
| 7 | 50 | 0.39 ± 0.03 |
| 7 | 70 | 0.25 ± 0.13 |
| 6 | 37 | 0.18 ± 0.12 |
| 7 | 37 | 0.54 ± 0.02 |
| 8 | 37 | 0.63 ± 0.08 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Leoni, C.; Manzari, C.; Chiara, M.; Veronico, P.; Bruno, G.L.; Pesole, G.; Ceci, L.R.; Volpicella, M. Chitinolytic Enzymes of the Hyperparasite Fungus Aphanocladium album: Genome-Wide Survey and Characterization of A Selected Enzyme. Microorganisms 2023, 11, 1357. https://doi.org/10.3390/microorganisms11051357
Leoni C, Manzari C, Chiara M, Veronico P, Bruno GL, Pesole G, Ceci LR, Volpicella M. Chitinolytic Enzymes of the Hyperparasite Fungus Aphanocladium album: Genome-Wide Survey and Characterization of A Selected Enzyme. Microorganisms. 2023; 11(5):1357. https://doi.org/10.3390/microorganisms11051357
Chicago/Turabian StyleLeoni, Claudia, Caterina Manzari, Matteo Chiara, Pasqua Veronico, Giovanni Luigi Bruno, Graziano Pesole, Luigi R. Ceci, and Mariateresa Volpicella. 2023. "Chitinolytic Enzymes of the Hyperparasite Fungus Aphanocladium album: Genome-Wide Survey and Characterization of A Selected Enzyme" Microorganisms 11, no. 5: 1357. https://doi.org/10.3390/microorganisms11051357