

Petroleum-Degrading Fungal Isolates for the Treatment of Soil Microcosms
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Decaying Wood Sampling
2.3. Soil Sampling and Characterisation
2.4. Isolation of Wood-Decay Decomposing Fungi and Indigenous Soil Fungi
2.5. Selection of Hydrocarbon-Degrading Fungal Isolates
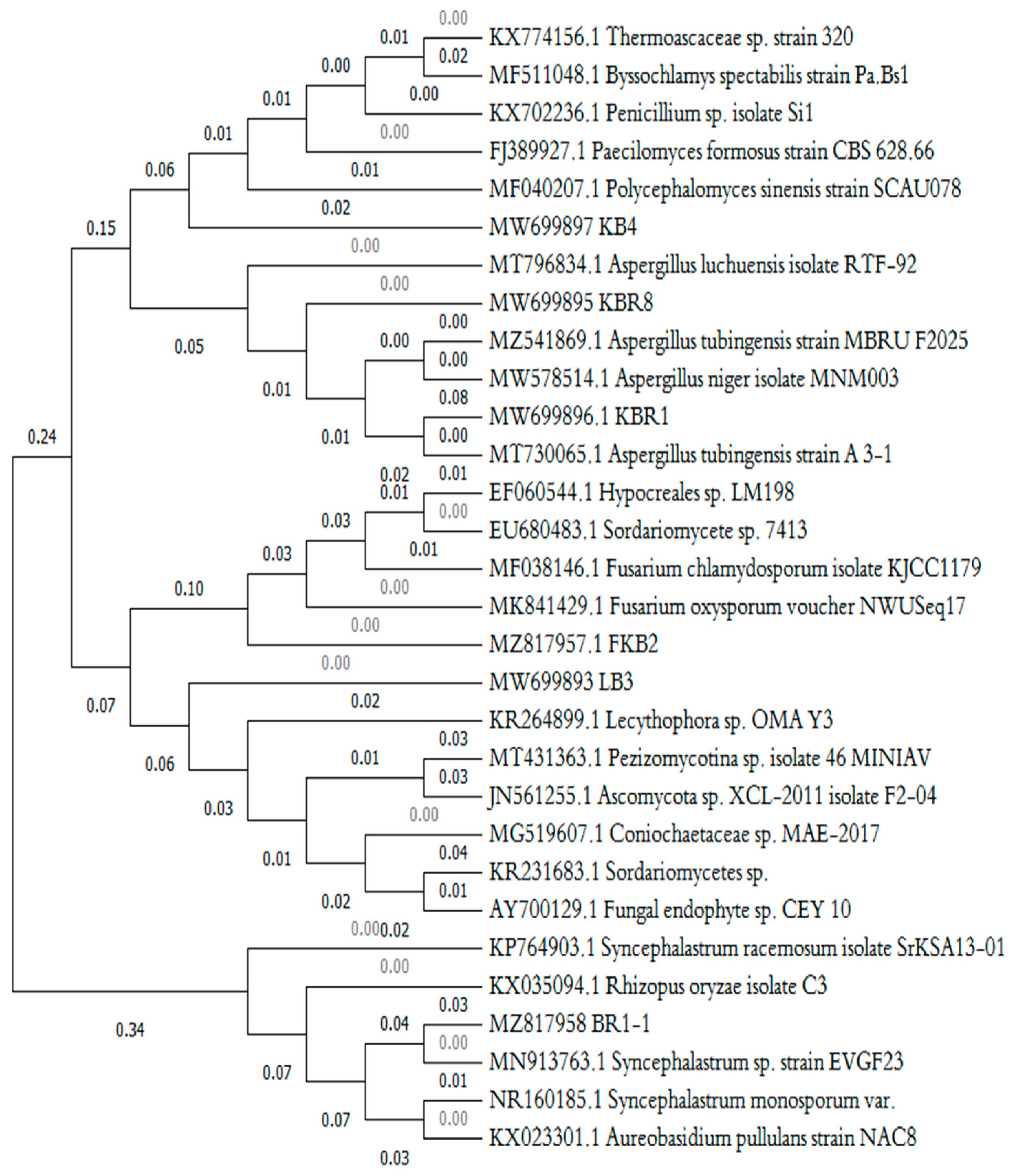
2.6. Molecular Identification and Phylogenetic Tree of Fungal Isolates
2.7. Soil Batch Microcosm Reactors Assays
2.7.1. Soil Preparation
2.7.2. Fungal Inoculum Preparation
2.7.3. Soil Microcosm Treatments
2.7.4. Analysis of Soil Batch Microcosm Parameters
2.7.5. Statistical Analysis
3. Results and Discussion
3.1. PHA-Contaminated Soil Sampling
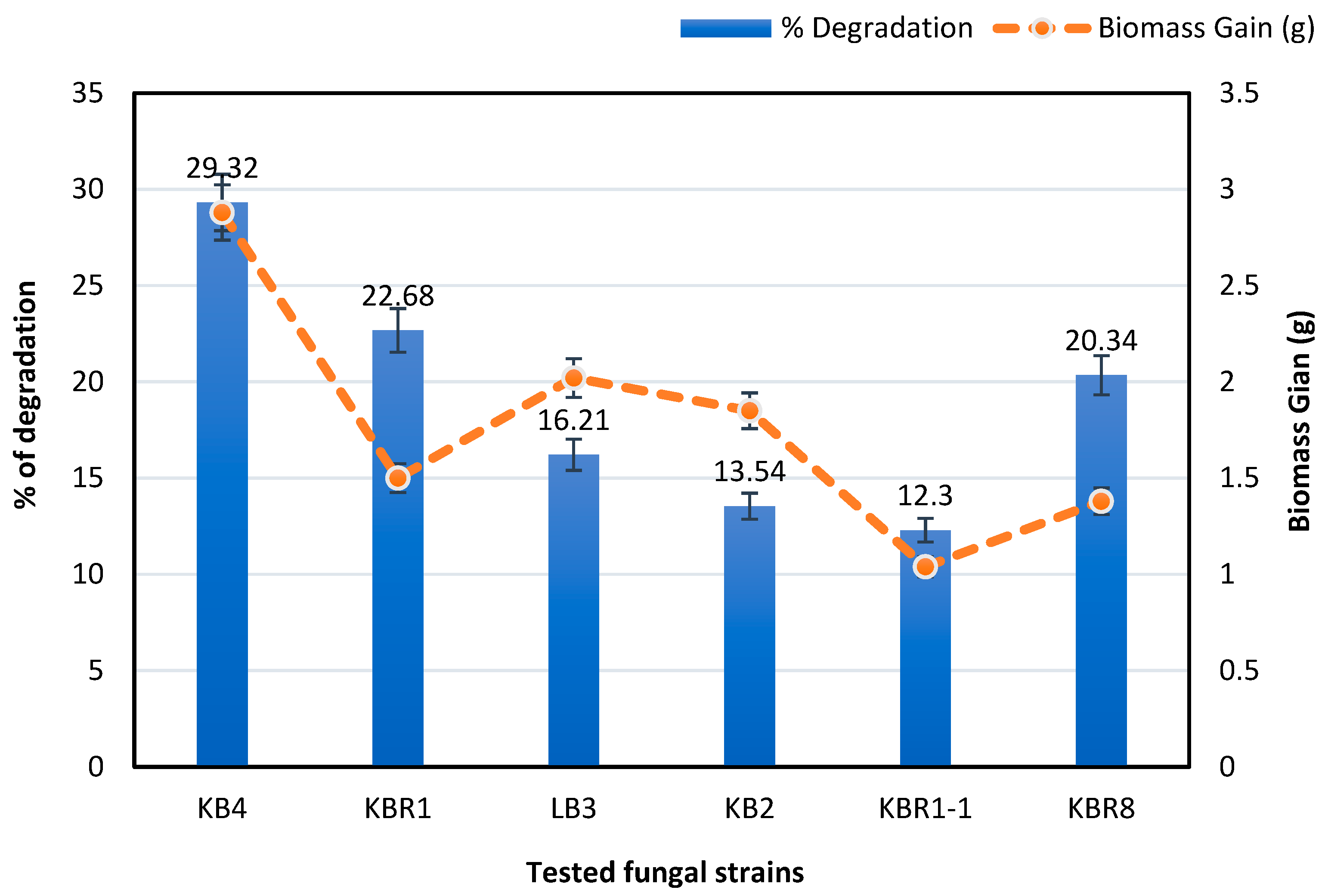
3.2. Isolation and Selection of Hydrocarbon-Degrading Fungal Isolates
3.3. Molecular Identification and Phylogenetic Analysis of the Selected Petroleum Hydrocarbon-Degrading Fungi
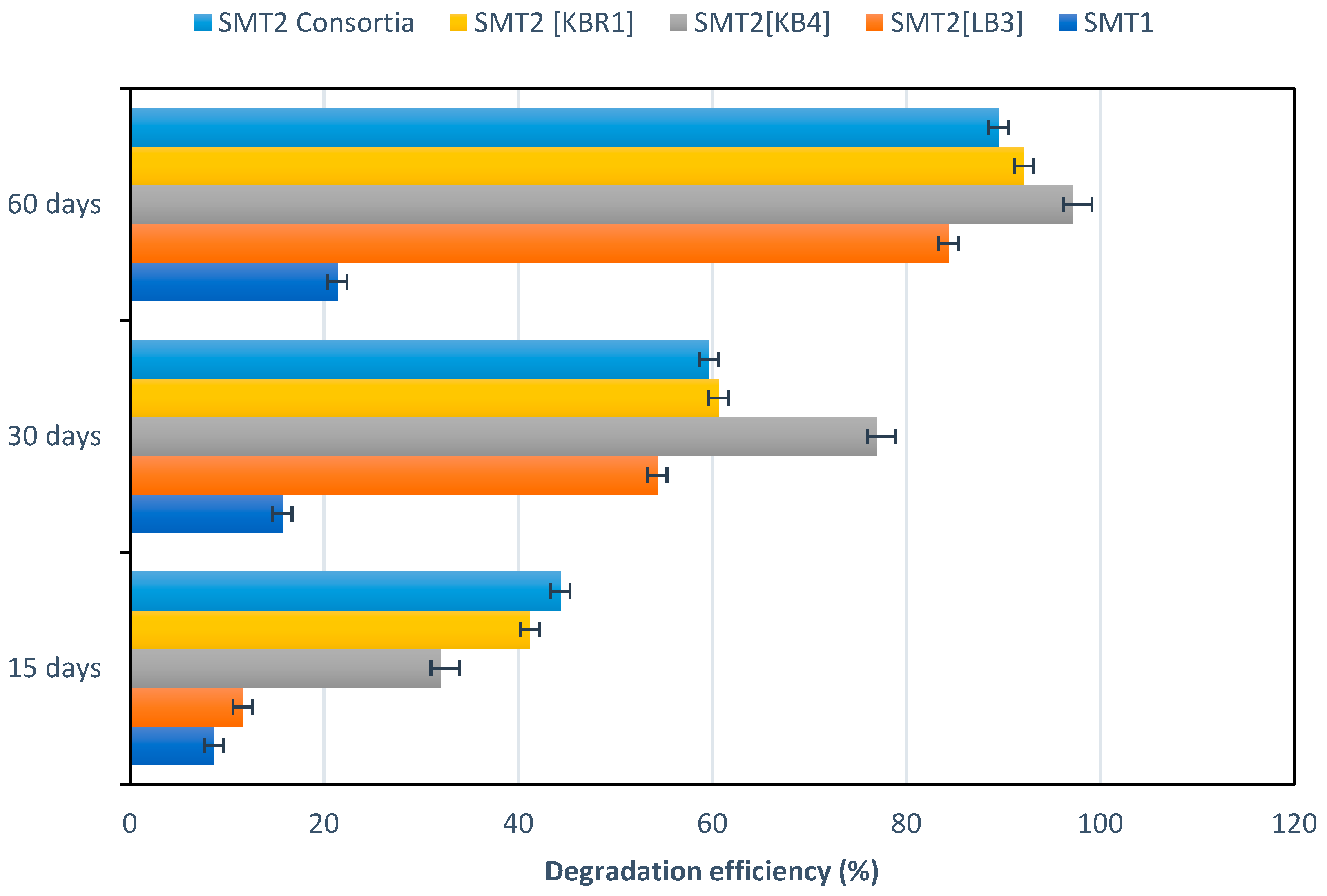
3.4. Microcosms for Petroleum-Contaminated Soil Remediation
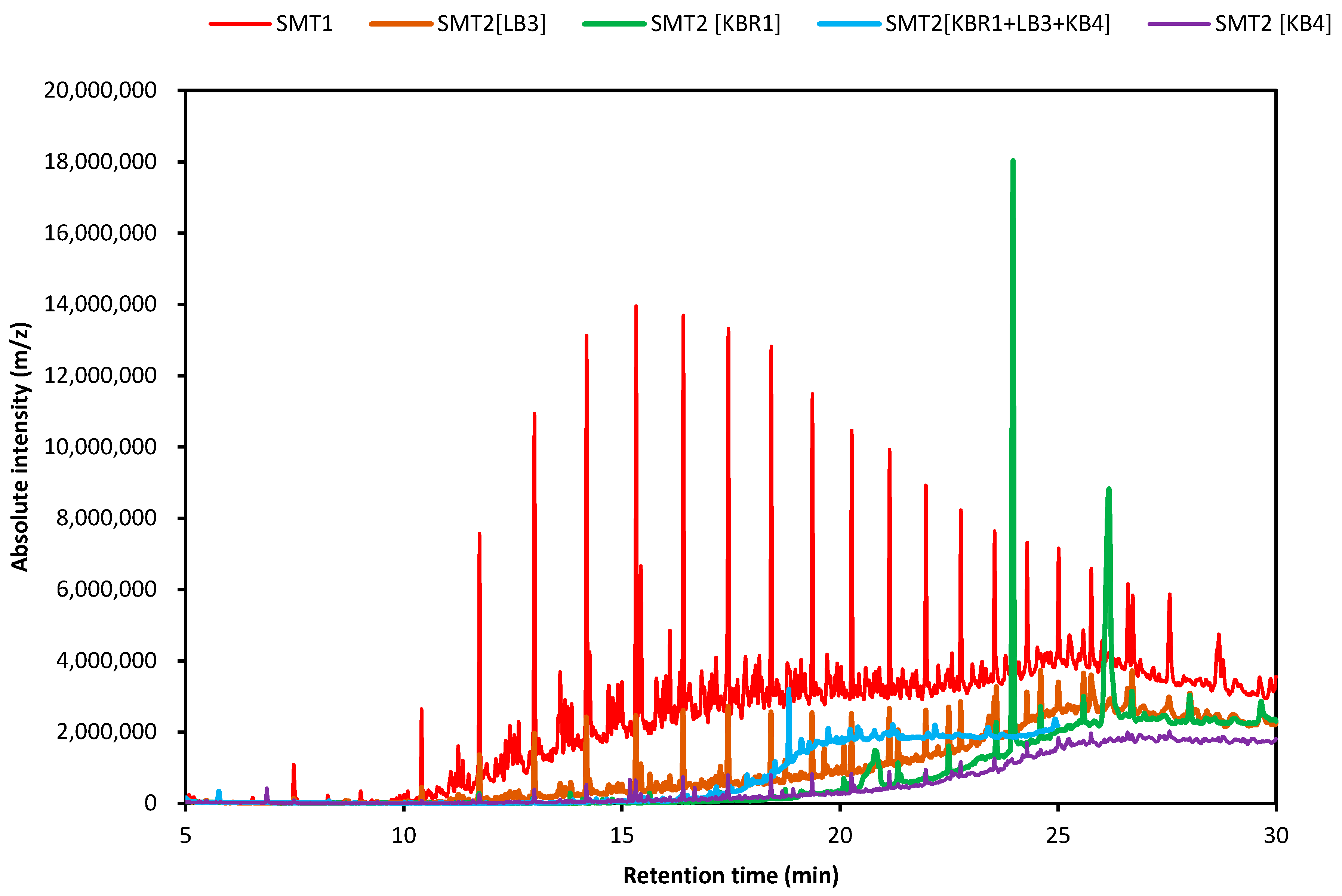
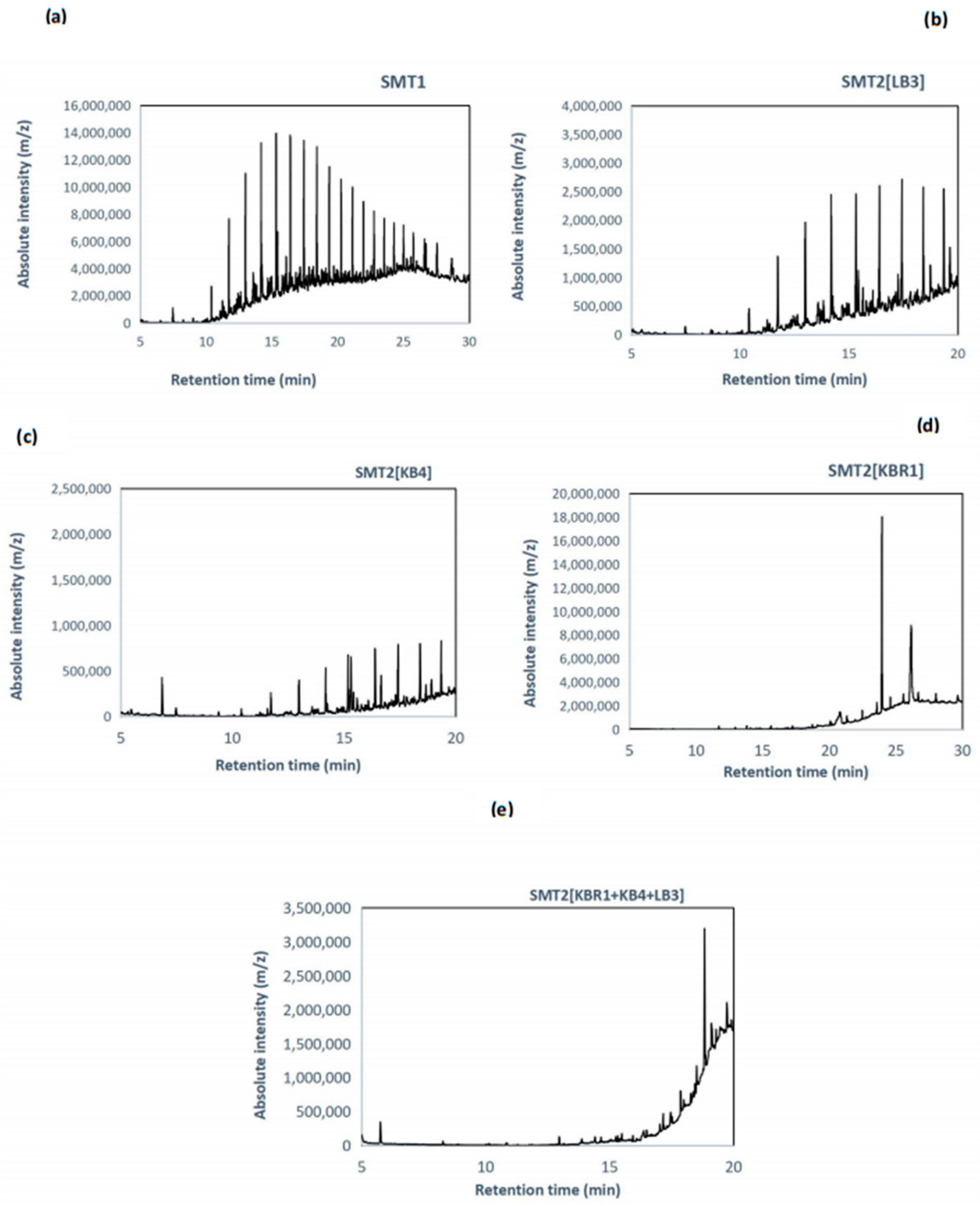
3.5. GC-MS Analysis for Soil Microcosms
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Asif, Z.; Chen, Z.; An, C.; Dong, J. Review Environmental Impacts and Challenges Associated with Oil Spills on Shorelines. J. Mar. Sci. Eng. 2022, 10, 762. [Google Scholar] [CrossRef]
- Ambade, B.; Sethi, S.S.; Kurwadkar, S.; Mishra, P.; Tripathee, L. Accumulation of polycyclic aromatic hydrocarbons (PAHs) in surface sediment residues of Mahanadi River Estuary: Abundance, source, and risk assessment. Mar. Pollut. Bull. 2022, 183, 114073. [Google Scholar] [CrossRef]
- Ismail, N.A.; Kasmuri, N.; Hamzah, N. Microbial Bioremediation Techniques for Polycyclic Aromatic Hydrocarbon (PAHs)—A Review. Water Air Soil Pollut. 2022, 233, 124. [Google Scholar] [CrossRef]
- Du, J.; Liu, J.; Jia, T.; Chai, B. The relationships between soil physicochemical properties, bacterial communities and polycyclic aromatic hydrocarbon concentrations in soils proximal to coking plants. Environ. Pollut. 2022, 298, 118823. [Google Scholar] [CrossRef] [PubMed]
- Daâssi, D.; Almaghrabi, F.Q. Petroleum-contaminated soil: Environmental occurrence and remediation strategies. 3 Biotech 2022, 12, 139. [Google Scholar] [CrossRef]
- Klamerus-Iwan, A.; Błońska, E.; Lasota, J.; Kalandyk, A.; Waligórski, P. Influence of Oil Contamination on Physical and Biological Properties of Forest Soil After Chainsaw Use. Water Air Soil Pollut. 2015, 226, 389. [Google Scholar] [CrossRef] [PubMed]
- Picariello, E.; Baldantoni, D.; De Nicola, F. Annual dynamics of indigenous microbial communities of forest soils after severe PAH contamination. Appl. Soil Ecol. 2023, 186, 104812. [Google Scholar] [CrossRef]
- Wu, D.; Kan, H.; Zhang, Y.; Wang, T.; Qu, G.; Zhang, P.; Jia, H.; Sun, H. Pyrene contaminated soil remediation using microwave/magnetite activated persulfate oxidation. Chemosphere 2022, 286 Pt 2, 131787. [Google Scholar] [CrossRef]
- Dutta, V.; Devasia, J.; Chauhan, A.; Jayalakshmi, M.; Vasantha, V.L.d.; Jha, A.; Nizam, A.; Andrew Lin, K.Y.; Ghotekar, S. Photocatalytic nanomaterials: Applications for remediation of toxic polycyclic aromatic hydrocarbons and green management. Adv. Chem. Eng. 2022, 11, 100353. [Google Scholar] [CrossRef]
- Kumar, L.; Chugh, M.; Kumar, S.; Kumar, K.; Sharma, J.; Bharadvaja, N. Remediation of petrorefinery wastewater contaminants: A review on physicochemical and bioremediation strategies. Process Saf. Environ. Prot. 2022, 159, 362–375. [Google Scholar] [CrossRef]
- Ławniczak, Ł.; Wozniak-Karczewska, M.; Loibner, A.P.; Heipieper, H.J.; Chrzanowski, Ł. Review Microbial Degradation of Hydrocarbons—Basic Principles for Bioremediation: A Review. Molecules 2020, 25, 856. [Google Scholar] [CrossRef] [PubMed]
- Picariello, E.; Baldantoni, D.; De Nicola, F. Investigating natural attenuation of PAHs by soil microbial communities: Insights by a machine learning approach. Restor. Ecol. 2022, 30, e13655. [Google Scholar] [CrossRef]
- Nwankwegu, A.S.; Zhang, L.; Xie, D.; Onwosi, C.O.; Muhammad, W.I.; Odoh, C.K.; Sam, K.; Idenyi, J.N. Bioaugmentation as a green technology for hydrocarbon pollution remediation. Problems and prospects. J. Environ. Manag. 2022, 304, 114313. [Google Scholar] [CrossRef]
- Volkoff, S.J.; Rodriguez, D.L.; Singleton, D.R.; McCumber, A.W.; Aitken, M.D.; Stewart, J.R.; Gunsch, C.K. Identifying bioaugmentation candidates for bioremediation of polycyclic aromatic hydrocarbons in contaminated estuarine sediment of the Elizabeth River, VA, USA. Appl. Microbiol. Biotechnol. 2022, 106, 1715–1727. [Google Scholar] [CrossRef]
- Kumar, B.L.; Gopal, D.V.R.S. Effective role of indigenous microorganisms for sustainable environment. 3 Biotech 2015, 5, 867–876. [Google Scholar] [CrossRef] [PubMed]
- Ezekoye, C.C.; Chikere, C.B.; Okpokwasili, G.C. Fungal diversity associated with crude oil-impacted soil undergoing in-situ bioremediation. Sustain. Chem. Pharm. 2018, 10, 148–152. [Google Scholar] [CrossRef]
- Kiamarsi, Z.; Kafi, M.; Soleimani, M.; Nezami, A.; Lutts, S. Conjunction of Vetiveria zizanioides L. and oil-degrading bacteria as a promising technique for remediation of crude oil-contaminated soils. J. Clean. Prod. 2020, 253, 119719. [Google Scholar] [CrossRef]
- Košnář, Z.; Wernerová, J.; Frühbauer, P.; Tlustoš, P. Chapter Bioremediation: Removal of Polycyclic Aromatic Hydrocarbons from Soil. Book Agricultural Biocatalysis, 1st ed.; ImprintJenny Stanford Publishing: Dubai, United Arab Emirates, 2022; p. 44. [Google Scholar]
- Wu, M.; Wu, J.; Zhang, X.; Ye, X. Effect of bioaugmentation and biostimulation on hydrocarbon degradation and microbial community composition in petroleum-contaminated loessal soil. Chemosphere 2019, 237, 124456. [Google Scholar] [CrossRef]
- Medaura, M.C.; Guivernau, M.; Moreno-Ventas, X.; Prenafeta-Boldú, F.X.; Viñas, M. Bioaugmentation of Native Fungi, an Efficient Strategy for the Bioremediation of an Aged Industrially Polluted Soil with Heavy Hydrocarbons. Front. Microbiol. 2021, 12, 626436. [Google Scholar] [CrossRef] [PubMed]
- Jackson, M.L. Soil Chemical Analysis; Prentice-Hall of India Pvt. Ltd.: New Delhi, India, 1967; p. 498. [Google Scholar]
- National Environment Protection NEP(Assessment of Site Contamination) Measure 1999 (as amended 2013).
- Daâssi, D.; Zouari-Mechichi, H.; Belbahri, L.; Barriuso, J.; Jesús Martínez, M.; Nasri, M.; Mechichi, T. Phylogenetic and metabolic diversity of Tunisian forest wood-degrading fungi: A wealth of novelties and opportunities for biotechnology. 3 Biotech 2016, 6, 46. [Google Scholar] [CrossRef]
- Warcup, J.H. The soil-plate method for isolation of fungi from soil. Nature 1950, 166, 117–118. [Google Scholar] [CrossRef]
- Agrawal, N.; Verma, P.; Shahi, S.K. Degradation of polycyclic aromatic hydrocarbons (phenanthrene and pyrene) by the ligninolytic fungi Ganoderma lucidum isolated from the hardwood stump. Bioresour. Bioprocess. 2018, 5, 11. [Google Scholar] [CrossRef]
- Daâssi, D.; Nasraoui-Hajaji, A.; Awasir, S.; Frikha, F.; Mechichi, T. Biodegradation of C20 carbon clusters from diesel fuel by Coriolopsis gallica: Optimization, metabolic pathway, phytotoxicity. 3 Biotech 2021, 11, 214. [Google Scholar] [CrossRef]
- Daâssi, D.; Zouari-Mechichi, H.; Martınez, M.J.; Nasri, M.; Mechichi, T. Purification and biochemical characterization of a new alkali-stable laccase from Trametes sp. Isolated in Tunisia: Role of the enzyme in olive mill wastewater treatment. World J. Microbiol. Biotechnol. 2013, 29, 2145–2155. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.D.; Higgins, D.G.; Gibsonet, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [PubMed]
- Saitou, N.; Nei, M. The Neighbour-joining method-A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [CrossRef]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Tchobanoglous, G.T.H.; Vigil, S. Integrated solid waste management. In Engineering Principles and Management Issues; McGraw Hill: New York, NY, USA, 1993; Volume 684–704, p. 992. [Google Scholar]
- Potin, O.; Rafin, C.; Veignie, E. Bioremediation of an aged polycyclic aromatic hydrocarbons (PAHs)-contaminated soil by filamentous fungi isolated from the soil. Int. Biodeter. Biodegr. 2004, 54, 45–52. [Google Scholar] [CrossRef]
- EPA 821-B-94-004; Method 1664: N-hexane Extractable Material and Silica Gel. Treated n-Hexane Extractable Material (SGT-HEM) by Extraction and Gravimetry (Oil and Grease and Total Petroleum Hydrocarbons). US EPA (United States Environmental Protection Agency): Washington, DC, USA, 1995.
- US EPA (United States Environmental Protection Agency). Method 3540C. Soxhlet Extraction, Part of Test Methods for Evaluating Solid Waste, Physical/Chemical Methods; US EPA (United States Environmental Protection Agency): Washington, DC, USA, 1996.
- Siddique, T.; Rutherford, P.M.; Arocena, J.M.; Thring, R.W. A proposed method for rapid and economical extraction of petroleum hydrocarbons from contaminated soils. Can. J. Soil Sci. 2005, 86, 725–728. [Google Scholar] [CrossRef]
- Bishnoi, N.R.; Mehta, U.; Sain, U.; Pandit, G.G. Quantification of polycyclic aromatic hydrocarbons in tea and coffee samples of Mumbai city (India) by high performance liquid chromatography. Environ. Monit. Assess. 2005, 107, 399–406. [Google Scholar] [CrossRef]
- Abbassi, B.E.; Shquirat, W.D. Kinetics of Indigenous Isolated Bacteria used for Ex-Situ Bioremediation of Petroleum Contaminated Soil. Water Air Soil Pollut. 2008, 192, 221–226. [Google Scholar] [CrossRef]
- Suarez, M.P.; Rifai, H.S. Biodegradation rates for fuel hydrocarbons and chlorinated solvents in groundwater. Bioremediat. J. 2005, 8, 37–41. [Google Scholar] [CrossRef]
- Torres, L.G.B.; Climent, M.; Saquelares, J.; Bandala, E.R.; Urquiza, G.; Iturbe, R. Characterization and treatability of a contaminated soil from an oil exploration zone. Int. J. Environ. Sci. Technol. 2007, 4, 311–322. [Google Scholar] [CrossRef]
- Abu Khamsin, S.A. Environmental Regulations for Drilling Operations in Saudi Arabia; SPE-39256-MS. In Proceedings of the SPE/IADC Middle East Drilling Technology Conference, Bahrain, 23 November 1997; Available online: https://onepetro.org/SPEMEDT/proceedings-abstract/97MEDT/All-97MEDT/SPE-39256-MS/189354 (accessed on 23 April 2023).
- Department of Petroleum Resources (DPR). Environmental Guidelines and Standards for the Petroleum Industry in Nigeria (EGASPIN); Country Nigeria EGASPIN; Department of Petroleum Resources: Lagos, Nigeria, 2002. [Google Scholar]
- Lotfinasabasl, S.; Gunale, V.R.; Rajurkar, N.S. Assessment of petroleum hydrocarbon degradation from soil and tarball by fungi. Biosci. Discv. 2012, 3, 186–192. [Google Scholar]
- Oboh, O.B.; Ilori, M.O.; Akinyemi, J.O.; Adebusoye, S.A. Hydrocarbon degrading potential of bacteria isolated from a Nigerian bitumen (tarsand) deposit. Nat. Sci. 2006, 4, 51–57. [Google Scholar]
- Rosselló-Mora, R.; Amann, R. The species concept for prokaryotes. FEMS Microbiol. Rev. 2001, 25, 39–67. [Google Scholar] [CrossRef]
- Lopez, M.J.; Vargas-García, M.d.-C.; Suárez-Estrella, F.; Nichols, N.N.; Dien, B.S.; Moreno, J. Lignocellulose-degrading enzymes produced by the ascomycete Coniochaeta ligniaria and related species: Application for a lignocellulosic substrate treatment. Enzym. Microb. Technol. 2007, 40, 794–800. [Google Scholar] [CrossRef]
- Bainier, G. Mycothèque de l’école de pharmacie. XI Paecilomyces, genre nouveau de mucédinées. Bull. Soc. Mycol. Fr. 1970, 23, 26–27. [Google Scholar]
- Al-Hawash, A.B.; Alkooranee, J.T.; Abbood, H.A.; Zhang, J.; Sun, J.; Zhang, X.; Ma, F. Isolation and characterization of two crude oil-degrading fungi strains from Rumaila oil field, Iraq. Biotechnol. Rep. 2018, 17, 104–109. [Google Scholar] [CrossRef]
- Li, Q.; Liu, J.; Gadd, G.M. Fungal bioremediation of soil co-contaminated with petroleum hydrocarbons and toxic metals. Appl. Microbiol. Biotechnol. 2020, 104, 8999–9008. [Google Scholar] [CrossRef]
- Pathak, U.; Jhunjhunwala, A.; Singh, S.; Bajaj, N.; Mandal, T. Chapter 19—Potentiality of enzymes as a green tool in degradation of petroleum hydrocarbons Advances in Oil-Water Separation. In A Complete Guide for Physical, Chemical, and Biochemical Processes; Elsevier: Amsterdam, The Netherlands, 2022; pp. 337–351. [Google Scholar]
- Hernández-Adame, N.M.; López-Miranda, J.; Martínez-Prado, M.A.; Cisneros-de la Cueva, S.; Rojas-Contreras, J.A.; Medrano-Roldán, H. Increase in Total Petroleum Hydrocarbons Removal Rate in Contaminated Mining Soil Through Bioaugmentation with Autochthonous Fungi During the Slow Bioremediation Stage. Water Air Soil Pollut. 2021, 232, 95. [Google Scholar] [CrossRef]
- Feng, L.; Jiang, X.; Huang, Y.; Wen, D.; Fu, T.; Fu, R. Petroleum hydrocarbon-contaminated soil bioremediation assisted by isolated bacterial consortium and sophorolipid. Environ. Pollut. 2021, 273, 116476. [Google Scholar] [CrossRef]
- Chukwura, E.I.; Ojiegbu, N.M.; Nwankwegu, A.S. Hydrocarbon Degradation Potentials of Fungi Associated with Oil-Contaminated Soil from Selected Mechanic Workshops in Awka, Anambra State, Nigeria. Front. Environ. Microbiol. 2016, 2, 38–44. [Google Scholar] [CrossRef]
- Obire, O.; Aleruchi, O.; Wemedo, S.A. Fungi in Biodegradation of Polycyclic Aromatic Hydrocarbons in Oilfield Wastewater. Acta Sci. Microbiol. 2020, 4, 3220–3224. [Google Scholar]
- Yuan, H.; Huang, S.; Yuan, J.; You, Y.; Zhang, Y. Characteristics of microbial denitrification under different aeration intensities: Performance, mechanism, and co-occurrence network. Sci. Total Environ. 2021, 754, 141965. [Google Scholar] [CrossRef] [PubMed]
- Abd El-Aziz, A.R.M.; Al-Othman, M.R.; Hisham, S.M.; Shehata, S.M. Evaluation of crude oil biodegradation using mixed fungal cultures. PLoS ONE 2021, 16, e0256376. [Google Scholar] [CrossRef] [PubMed]
- Daâssi, D.; Almaghrabi, F.Q. Soil Microcosms for Bioaugmentation With Fungal Isolates to Remediate Petroleum Hydrocarbon-contaminated Soil. 2021; Preprint. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Property | Value |
|---|---|
| Physical Parameters | |
| Moisture Content (%) | 1.6 |
| pH | 6.8 |
| Total Petroleum Hydrocarbon (TPH) | |
| C6–C9 Fraction (mg kg−1) | <10 |
| C10–C14 Fraction (mg kg−1) | <50 |
| C15–C28 Fraction (mg kg−1) | 13,400 |
| C29–C36 Fraction (mg kg−1) | 7370 |
| C37–C40 Fraction (mg kg−1) | 2720 |
| C10–C40 Fraction (sum) (mg kg−1) | 23,500 |
| Volatile Organic Compounds-BTEX | |
| Benzene (mg kg−1) | 0.2 |
| Ethylbenzene (mg kg−1) | 0.5 |
| Meta- and Para-Xylene (mg kg−1) | 0.5 |
| Ortho-Xylene (mg kg−1) | 0.5 |
| Volatile Organic Compounds—Surrogates | |
| 1.2-Dichloroethane-D4 (%) | 100 |
| Toluene-D8 (%) | 99.3 |
| 4-Bromofluorobenzene (%) | 95.9 |
| Number of Isolates | Code of Isolate | Origin of Isolate | Max Identity (%) | Strains of Closed Match (Accession Number) | Identification | Accession Number | Phylum |
|---|---|---|---|---|---|---|---|
| Strain 1 | KBR1-1 | Wood | 99.58% | Syncephalastrum sp. EVGF23 (MN913763.1) | Syncephalastrum sp. | MZ817958 | Zygomycota |
| Strain 2 | KBR1 | PHs-Contaminated Soil | 95.54% | Aspergillus niger MNM003 (MW578514.1) | Aspergillus niger. | MW699896 | Ascomycota |
| Strain 3 | KB4 | Wood | 98.39% | Paecilomyces formosus CBS 628.66 (FJ389927.1) | Paecilomyces formosus | MW699897 | Ascomycota |
| Strain 4 | KB2 | Wood | 98.84% | Fusarium chlamydosporum isolate KJCC1179 (MF038146.1) | Fusarium chlamydosporum | MZ817957 | Ascomycota |
| Strain 5 | KBR8 | PHs-Contaminated Soil | 98.88% | Aspergillus niger (MW142509.1) | Aspergillus tubingensis | MW699895 | Ascomycota |
| Strain 6 | LB3 | Wood | 99.68% | Coniochaetaceae sp. MAE-2017 (MG519607.1) | Coniochaetaceae sp. | MW699893 | Ascomycota |
| Inputs | Soil Microcosms Treatments | |
|---|---|---|
| SMT1 | SMT2 | |
| Native soil (g) | + | − |
| Sterile Soil (g) | − | + |
| Nutrients C:N:P ratio of 100:15:1 | + | + |
| Water | + | + |
| Fungal strains | ||
| Aspergillus niger [MW699896] KBR1 | − | + |
| Coniochaeta sp. [MW699893] LB3 | − | + |
| Paecilomyces formosus [MW699897] KB4 | − | + |
| Fungal consortium culture: KBR1 + LB3 + KB4 | − | + |
| Removal Rate of TPH (%) | TPH Removal Rate (%) | k (1/Day) | t1/2 (Day) |
|---|---|---|---|
| SMT2 [KB4] | 97.17933 ± 1.15 | 5.95 × 10−2 ± 1.71 × 10−6 | 11.65542 |
| SMT2 [KBR1] | 95.12733 ± 1.23 | 5.04 × 10−2 ± 2.1 × 10−7 | 13.76417 |
| SMT2 [KBR1 + LB3 + KB4] | 89.487 ± 2.04 | 3.75 × 10−2 ± 0.7 × 10−6 | 18.46294 |
| SMT2 [LB3] | 84.361 ± 1.65 | 3.09 × 10−2 ± 4.3 × 10−7 | 22.41499 |
| SMT1 | 21.36 ± 2.35 | 4.01 × 10−2 ± 1.3 × 10−7 | 173.0779 |
| SMT1 | |||||
|---|---|---|---|---|---|
| RT | Area% | Height% | Compounds | Molecular Formula | Fractions |
| 11.735 | 5.34 | 1.81 | Heptadecane | C17H36 | C15–C28 |
| 12.995 | 7.64 | 1.98 | Heptadecane | C17H36 | |
| 14.191 | 8.89 | 1.93 | Heptadecane | C17H36 | |
| 15.326 | 9.2 | 1.82 | Eicosane | C20H42 | |
| 15.436 | 3.43 | 2.34 | Eicosane | C20H42 | |
| 16.407 | 8.75 | 1.81 | Eicosane | C20H42 | |
| 17.15 | 1.15 | 5.02 | Heptadecane, 2-methyl | C18H38 | |
| 17.439 | 8.5 | 2.07 | Eicosane | C20H42 | |
| 18.142 | 0.83 | 5.11 | 2-methylhexacosane | C27H56 | |
| 18.423 | 7.84 | 1.88 | Eicosane | C20H42 | |
| 19.366 | 6.7 | 1.87 | Eicosane | C20H42 | |
| 20.268 | 5.93 | 1.89 | Eicosane | C20H42 | |
| 21.136 | 5.46 | 1.9 | Tetracosane | C24H50 | |
| 21.969 | 4.44 | 1.83 | Hexatriacontane | C36H74 | C29–C36 |
| 22.771 | 3.88 | 2.01 | Hexatriacontane | C36H74 | |
| 23.544 | 3.16 | 1.82 | Hexatriacontane | C36H74 | |
| 24.289 | 2.68 | 1.89 | Hexatriacontane | C36H74 | |
| 25.011 | 2.4 | 1.96 | Hexatriacontane | C36H74 | |
| 25.76 | 2.08 | 2.18 | Tetrapentacontane | C54H110 | C37–C40 |
| 27.558 | 1.7 | 3.27 | 2-methylhexacosane | C27H56 | C15–C28 |
| SMT2 [LB3] | |||||
| 11.733 | 4.12 | 1.71 | Heptadecane | C17H36 | C15–C28 |
| 12.991 | 6.07 | 1.85 | Heptadecane | C17H36 | |
| 14.184 | 7.16 | 1.76 | Heptadecane | C17H36 | |
| 15.32 | 7.13 | 1.81 | Heneicosane | C21H44 | |
| 15.432 | 2.54 | 2.3 | Eicosane | C20H42 | |
| 16.401 | 7.28 | 1.78 | Heneicosane | C21H44 | |
| 17.432 | 7.2 | 1.8 | Eicosane | C20H42 | |
| 18.417 | 6.71 | 1.77 | Eicosane | C20H42 | |
| 19.359 | 6.18 | 1.79 | Eicosane | C20H42 | |
| 20.263 | 5.69 | 1.75 | Eicosane | C20H42 | |
| 21.131 | 5.3 | 1.89 | Eicosane | C20H42 | |
| 21.329 | 3.07 | 1.94 | Cyclononasiloxane, octadecamethyl- | C18H54O9Si9 | |
| 21.964 | 4.31 | 1.86 | Eicosane | C20H42 | |
| 22.492 | 4.03 | 1.94 | Cyclononasiloxane, octadecamethyl- | C18H54O9Si9 | |
| 22.766 | 4.4 | 1.8 | Hexatriacontane | C36H74 | C29–C36 |
| 23.539 | 3.37 | 1.89 | Tetracontane | C40H82 | C37–C40 |
| 23.584 | 4.65 | 1.9 | Cyclononasiloxane, octadecamethyl- | C18H54O9Si9 | C15–C28 |
| 24.6 | 4.17 | 1.97 | Cyclononasiloxane, octadecamethyl- | C18H54O9Si9 | |
| 25.582 | 3.19 | 2.25 | Cyclodecasiloxane, eicosamethyl- | C20H60O10Si10 | |
| 26.696 | 3.43 | 2.5 | Cyclononasiloxane, octadecamethyl- | C18H54O9Si9 | |
| SMT2 [KB4] | |||||
| 6.858 | 4.67 | 2.02 | Cyclopentasiloxane, decamethyl- | C10H30O5Si5 | C10–C14 |
| 12.99 | 4.12 | 1.73 | Hexadecane | C16H34 | C15–C28 |
| 14.184 | 5.61 | 1.78 | Heptadecane | C17H36 | |
| 15.184 | 7.13 | 1.74 | Methyl 13-methyltetradecanoate | C16H32O2 | |
| 15.318 | 6.85 | 1.83 | Heneicosane | C21H44 | |
| 16.4 | 7.51 | 1.73 | Heneicosane | C21H44 | |
| 16.669 | 4.14 | 1.86 | Hexadecanoic acid, methyl ester | C17H34O2 | |
| 17.431 | 7.44 | 1.88 | Eicosane | C20H42 | |
| 18.417 | 6.84 | 1.89 | Eicosane | C20H42 | |
| 18.667 | 1.69 | 2.89 | Methyl stearate | C19H38O2 | |
| 19.359 | 6.9 | 1.74 | Eicosane | C20H42 | |
| 20.262 | 6.15 | 1.89 | Eicosane | C20H42 | |
| 21.13 | 5.67 | 1.93 | Eicosane | C20H42 | |
| 21.962 | 4.66 | 1.95 | Eicosane | C20H42 | |
| 22.558 | 2.31 | 5.05 | Bumetrizole | C17H18ClN3O | |
| 22.765 | 4.8 | 1.84 | Eicosane | C20H42 | |
| 23.539 | 3.56 | 1.71 | Tetrapentacontane | C54H110 | C37–C40 |
| 24.284 | 4.34 | 1.6 | 1-Eicosanol, 2-hexadecyl | C36H74O | C29–36 |
| 25.005 | 2.79 | 2.02 | Hexatriacontane | C36H74 | |
| 25.753 | 2.82 | 1.96 | Hexatriacontane | C36H74 | |
| SMT2 [KBR1 + KB4 + LB3] | |||||
| 5.758 | 6.34 | 2.1 | Cyclotetrasiloxane, octamethyl- | C8H24O4Si4 | C6–C9 |
| 17.869 | 6.65 | 1.51 | Cyclononasiloxane, octadecamethyl | C18H54O9Si9 | C15–C28 |
| 18.826 | 42.13 | 1.77 | 1-benzylindole | C15H13N | |
| 19.108 | 7.45 | 2.56 | Cyclodecasiloxane, eicosamethyl | C20H60O10Si10 | |
| 19.727 | 8.31 | 2.36 | Cyclononasiloxane, octadecamethyl | C18H54O9Si9 | |
| 20.23 | 5.38 | 2.17 | 13.17.21-Trimethylheptatriacontane | C7H14 | C6-C9 |
| 20.408 | 6.45 | 1.94 | Cyclodecasiloxane, eicosamethyl | C20H60O10Si10 | C15–C28 |
| 21.209 | 6.1 | 2.8 | Tetracosamethyl-cyclododecasiloxane | C24H72O12Si | |
| 22.178 | 6.44 | 2.94 | Tetracosamethyl-cyclododecasiloxane | C24H72O12Si | |
| 23.394 | 4.75 | 3.62 | Tetracosamethyl-cyclododecasiloxane | C24H72O12Si | |
| SMT2 [KBR1] | |||||
| 20.8 | 1.39 | 5.44 | Silane, diethylheptyloxyoctadecyloxy | C29H62O2Si | C29–C36 |
| 22.489 | 2.59 | 2 | Cyclodecasiloxane, eicosamethyl- | C20H60O10Si10 | C15–C28 |
| 23.581 | 3.21 | 2.08 | Tetracosamethyl-cyclododecasiloxane | C24H72O12Si | |
| 23.966 | 58.04 | 2.78 | 1.3-Benzenedicarboxylic acid, bis(2-ethylhexyl) ester | C24H38O · | |
| 24.598 | 3.15 | 2.04 | Tetracosamethyl-cyclododecasiloxane | C24H72O12Si | |
| 25.581 | 2.58 | 2.18 | Tetracosamethyl-cyclododecasiloxane | C24H72O12Si | |
| 26.163 | 22.36 | 7.93 | Propanoic acid, 3.3’-thiobis-, didodecyl ester | C30H58O4S | |
| 26.688 | 2.56 | 2.88 | Tetracosamethyl-cyclododecasiloxane | C24H72O12Si | |
| 28.015 | 2.51 | 3.53 | Tetracosamethyl-cyclododecasiloxane | C24H72O12Si | |
| 29.654 | 1.61 | 4.06 | Tetracosamethyl-cyclododecasiloxane | C24H72O12Si | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Daâssi, D.; Almaghrabi, F.Q. Petroleum-Degrading Fungal Isolates for the Treatment of Soil Microcosms. Microorganisms 2023, 11, 1351. https://doi.org/10.3390/microorganisms11051351
Daâssi D, Almaghrabi FQ. Petroleum-Degrading Fungal Isolates for the Treatment of Soil Microcosms. Microorganisms. 2023; 11(5):1351. https://doi.org/10.3390/microorganisms11051351
Chicago/Turabian StyleDaâssi, Dalel, and Fatimah Qabil Almaghrabi. 2023. "Petroleum-Degrading Fungal Isolates for the Treatment of Soil Microcosms" Microorganisms 11, no. 5: 1351. https://doi.org/10.3390/microorganisms11051351