Localization of C Cycle Enzymes in Arable and Forest Phaeozems within Levels of Soil Microstructure


Abstract
:1. Introduction
2. Materials and Methods
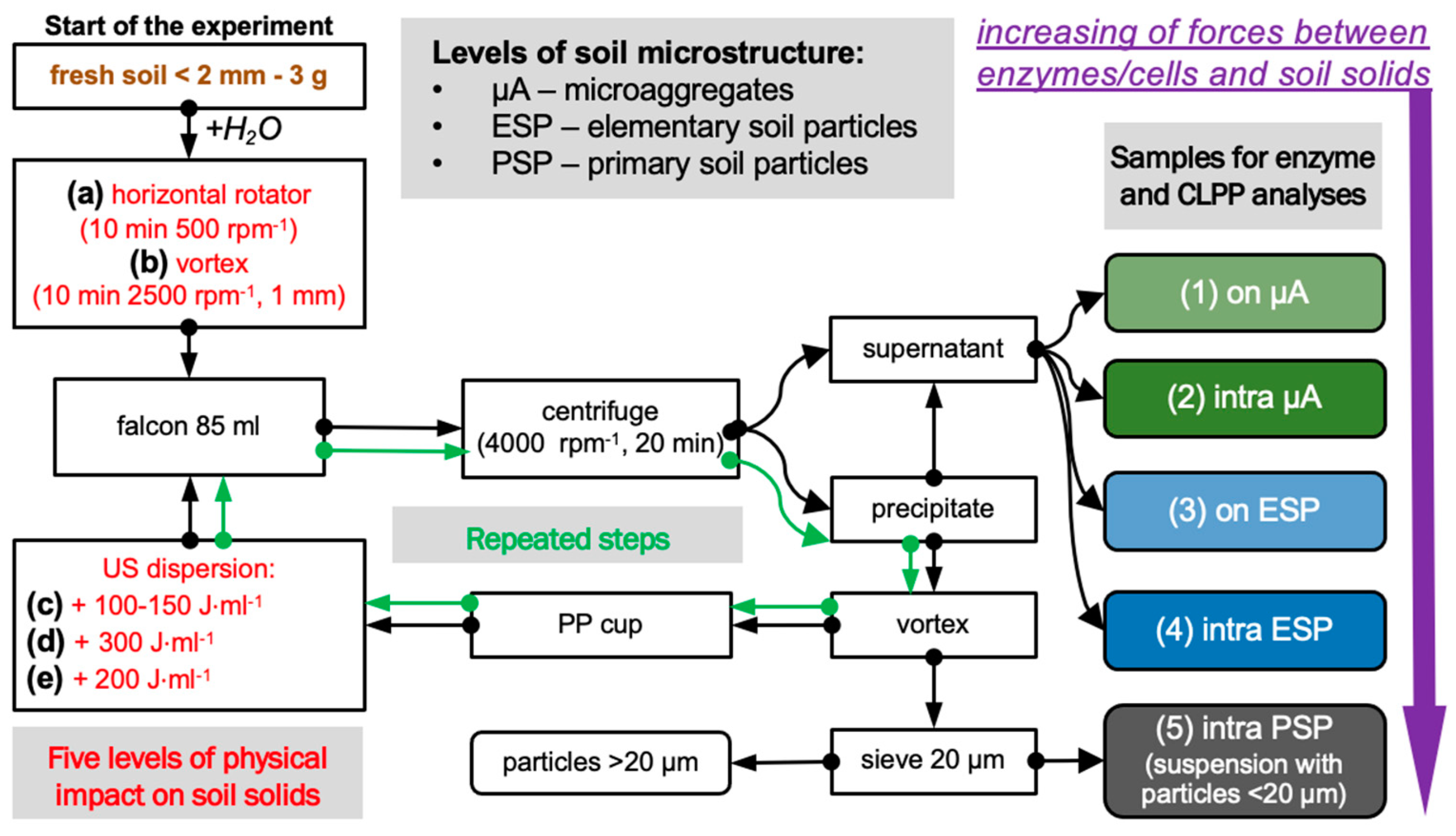
2.1. Experimental Design
2.2. Study Site, Soil Sampling, and Basic Properties
2.3. Enzyme Activity (EA) Analyses
2.4. Community-Level Physiological Profiling (CLPP)
2.5. Statistical Analyses
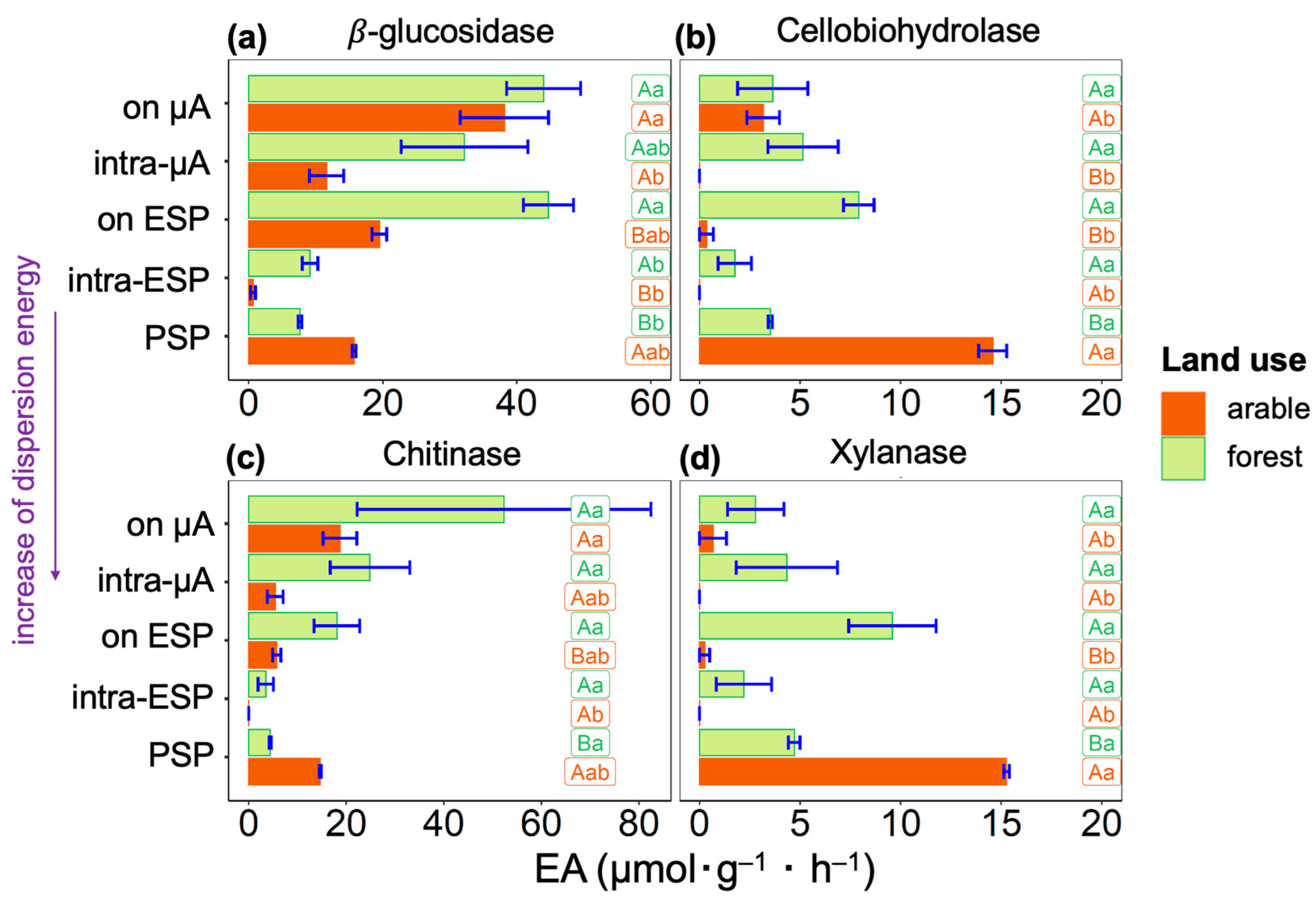
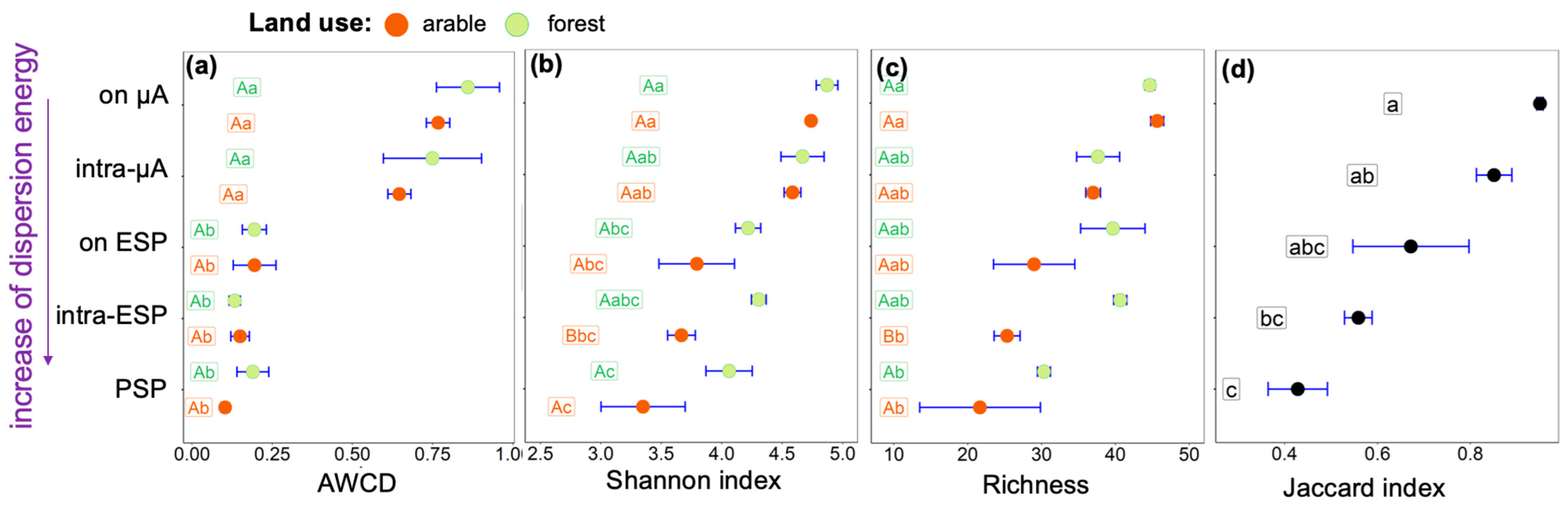
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
| Pentoses | Arabinose, ribose, xylose |
| Hexoses | Glucose, fructose, rhamnose |
| Oligoses | Cellobiose, lactose, maltose, sucrose |
| Salts of carboxylic acids | Acetate, aspartate, citrate, succinate, maleinate, pyruvate, octanoate, lactate |
| Amino acids | Glycine, proline, leucine, methionine, histidine, alanine, asparagine, valine, serine, phenylalanine, glutamine, arginine, lysine |
| Alcohols | Dulcitol, glycerol, inositol, sorbitol, mannitol |
| Polymers | Soluble starch, corn starch, Dextran 500, Tween 20, Tween 80, gelatin, pullulan |
| Miscellaneous (amides, amines phosphorylated carbons) | Creatinine, carbamide, β-glycerophosphate, glucosamine sulfate |
| Enzymes Tested | Associated with Soil Solids < 20 µm | Sum of Activities, Determined in Supernatants | ||
|---|---|---|---|---|
| Forest | Arable | Forest | Arable | |
| β-glucosidase | 7.63 ± 0.54 Ab | 15.74 ± 0.55 Bb | 130.07 ± 36.48 Aa | 69.87 ± 19.01 Ba |
| Cellobiohydrolase | 3.52 ± 0.22 Bb | 14.58 ± 1.39 Aa | 18.46 ± 2.21 Aa | 3.51 ± 0.95 Bb |
| Chitinase | 4.39 ± 0.42 Ba | 14.66 ± 0.33 Ab | 98.74 ± 88.11 Aa | 29.83 ± 8.82 Aa |
| Xylanase | 4.71 ± 0.57 Bb | 15.27 ± 0.28 Aa | 18.92 ± 10.63 Aa | 0.91 ± 1.26 Bb |
References
- Young, I.M.; Crawford, J.W. Interactions and self-organization in the soil-microbe complex. Science 2004, 304, 1634–1637. [Google Scholar] [CrossRef] [PubMed]
- Nunan, N.; Leloup, J.; Ruamps, L.S.; Pouteau, V.; Chenu, C. Effects of habitat constraints on soil microbial community function. Sci. Rep. 2007, 7, 4280. [Google Scholar] [CrossRef]
- Schimel, J.P. Life in dry soils: Effects of drought on soil microbial communities and processes. Annu. Rev. Ecol. Evol. Syst. 2018, 49, 409–432. [Google Scholar] [CrossRef]
- Tisdall, J.M.; Oades, J.M. Organic matter and water-stable aggregates in soils. J. Soil Sci. 1982, 33, 141–163. [Google Scholar] [CrossRef]
- Yudina, A.; Kuzyakov, Y. Dual nature of soil structure: The unity of aggregates and pores. Geoderma 2023, 434, 116478. [Google Scholar] [CrossRef]
- Alkorta, I.; Aizpurua, A.; Riga, P.; Albizu, I.; Amézaga, I.; Garbisu, C. Soil enzyme activities as biological indicators of soil health. Rev. Environ. Health 2003, 18, 65–73. [Google Scholar] [CrossRef]
- Nannipieri, P. Role of stabilised enzymes in microbial ecology and enzyme extraction from soil with potential applications in soil proteomics. In Nucleic Acids and Proteins in Soil; Nannipieri, P., Smalla, K., Eds.; Springer: Berlin/Heidelberg, Germany, 2006; pp. 75–94. [Google Scholar]
- Nunan, N.; Schmidt, H.; Raynaud, X. The ecology of heterogeneity: Soil bacterial communities and C dynamics. Philos. Trans. R. Soc. B 2020, 375, 20190249. [Google Scholar] [CrossRef]
- Allison, S.D.; Weintraub, M.N.; Gartner, T.B.; Waldrop, M.P. Evolutionary-economic principles as regulators of soil enzyme production and ecosystem function. In Soil Enzymology; Springer: Berlin/Heidelberg, Germany, 2011; pp. 229–243. [Google Scholar]
- McCarthy, J.F.; Ilavsky, J.; Jastrow, J.D.; Mayer, L.M.; Perfect, E.; Zhuang, J. Protection of organic carbon in soil microaggregates via restructuring of aggregate porosity and filling of pores with accumulating organic matter. Geochim. Cosmochim. Acta 2008, 72, 4725–4744. [Google Scholar] [CrossRef]
- Rabot, E.; Wiesmeier, M.; Schlüter, S.; Vogel, H.J. Soil structure as an indicator of soil functions: A review. Geoderma 2018, 314, 122–137. [Google Scholar] [CrossRef]
- Totsche, K.U.; Amelung, W.; Gerzabek, M.H.; Guggenberger, G.; Klumpp, E.; Knief, C.; Lehndorff, E.; Mikutta, R.; Peth, S.; Prechtel, A.; et al. Microaggregates in soils. J. Plant Nutr. Soil Sci. 2018, 181, 104–136. [Google Scholar] [CrossRef]
- Yudina, A.V.; Fomin, D.S.; Kotelnikova, A.D.; Milanovskii, E.Y. From the notion of elementary soil particle to the particle-size and microaggregate-size distribution analyses: A review. Eurasian Soil Sci. 2018, 51, 1326–1347. [Google Scholar] [CrossRef]
- Powlson, D.S. The effects of grinding on microbial and non-microbial organic matter in soil. J. Soil Sci. 1980, 31, 77–85. [Google Scholar] [CrossRef]
- Sollins, P.; Homann, P.; Caldwell, B.A. Stabilization and destabilization of soil organic matter: Mechanisms and controls. Geoderma 1996, 74, 65–105. [Google Scholar] [CrossRef]
- Dungait, J.A.; Hopkins, D.W.; Gregory, A.S.; Whitmore, A.P. Soil organic matter turnover is governed by accessibility not recalcitrance. Glob. Change Biol. 2012, 18, 1781–1796. [Google Scholar] [CrossRef]
- Kravchenko, A.N.; Guber, A.K. Soil pores and their contributions to soil carbon processes. Geoderma 2017, 287, 31–39. [Google Scholar] [CrossRef]
- Schimel, J.P.; Weintraub, M.N. The implications of exoenzyme activity on microbial carbon and nitrogen limitation in soil: A theoretical model. Soil Biol. Biochem. 2003, 35, 549–563. [Google Scholar] [CrossRef]
- Malik, A.A.; Puissant, J.; Goodall, T.; Allison, S.D.; Griffiths, R.I. Soil microbial communities with greater investment in resource acquisition have lower growth yield. Soil Biol. Biochem. 2019, 132, 36–39. [Google Scholar] [CrossRef]
- Nannipieri, P.; Giagnoni, L.; Renella, G.; Puglisi, E.; Ceccanti, B.; Masciandaro, G.; Fornasier, F.; Moscatelli, M.C.; Marinari, S. Soil enzymology: Classical and molecular approaches. Biol. Fertil. Soils. 2012, 48, 743–762. [Google Scholar] [CrossRef]
- Khaziev, F.K. Methods of Soil Enzymology; Nauka: Moscow, Russia, 2005; 252p. [Google Scholar]
- Perez Mateos, M.; Gonzalez Carcedo, S. Effect of fractionation on location of enzyme activities in soil structural units. Biol. Fertil. Soils 1985, 1, 153–159. [Google Scholar] [CrossRef]
- Stemmer, M.; Gerzabek, M.H.; Kandeler, E. Organic matter and enzyme activity in particle-size fractions of soils obtained after low-energy sonication. Soil Biol. Biochem. 1998, 30, 9–17. [Google Scholar] [CrossRef]
- Stemmer, M.; Gerzabek, M.H.; Kandeler, E. Invertase and xylanase activity of bulk soil and particle-size fractions during maize straw decomposition. Soil Biol. Biochem. 1998, 31, 9–18. [Google Scholar] [CrossRef]
- Kandeler, E.; Stemmer, M.; Klimanek, E.M. Response of soil microbial biomass, urease and xylanase within particle size fractions to long-term soil management. Soil Biol. Biochem. 1999, 31, 261–273. [Google Scholar] [CrossRef]
- Zhang, Q.; Zhou, W.; Liang, G.; Sun, J.; Wang, X.; He, P. Distribution of soil nutrients, extracellular enzyme activities and microbial communities across particle-size fractions in a long-term fertilizer experiment. Appl. Soil Ecol. 2015, 94, 59–71. [Google Scholar] [CrossRef]
- Le Roux, X.; Bouskill, N.J.; Niboyet, A.; Barthes, L.; Dijkstra, P.; Field, C.B.; Hungate, B.A.; Lerondelle, C.; Pommier, T.; Tang, J.; et al. Predicting the responses of soil nitrite-oxidizers to multi-factorial global change: A trait-based approach. Front. Microbiol. 2016, 7, 628. [Google Scholar] [CrossRef] [PubMed]
- Ingwersen, J.; Poll, C.; Streck, T.; Kandeler, E. Micro-scale modelling of carbon turnover driven by microbial succession at a biogeochemical interface. Soil Biol. Biochem. 2008, 40, 864–878. [Google Scholar] [CrossRef]
- Schweizer, S.A. Perspectives from the Fritz-Scheffer Awardee 2021: Soil organic matter storage and functions determined by patchy and piled-up arrangements at the microscale. J. Plant Nutr. Soil Sci. 2022, 185, 694–706. [Google Scholar] [CrossRef]
- Shoba, S.A.; Dobrovolsky, G.V.; Alyabina, I.O. National Atlas of Soils of the Russian Federation; Astrel: Moscow, Russia, 2006; pp. 130–131. [Google Scholar]
- Yudina, A.V.; Milanovskiy, Y.Y. The microaggregate analysis of soils by the method of laser diffraction: The specificities of sample preparation and result interpretation. Dokuchaev Soil Bull. 2017, 89, 3–20. [Google Scholar] [CrossRef]
- Schmidt, M.W.I.; Rumpel, C.; Kögel-Knabner, I. Evaluation of an ultrasonic dispersion procedure to isolate primary organomineral complexes from soils. Eur. J. Soil Sci. 1999, 50, 87–94. [Google Scholar] [CrossRef]
- Amelung, W.; Zech, W. Minimisation of organic matter disruption during particle-size fractionation of grassland epipedons. Geoderma 1999, 92, 73–85. [Google Scholar] [CrossRef]
- Graf-Rosenfellner, M.; Kayser, G.; Guggenberger, G.; Kaiser, K.; Büks, F.; Kaiser, M.; Mueller, C.W.; Schrumpf, M.; Rennert, T.; Welp, G.; et al. Replicability of aggregate disruption by sonication—An inter-laboratory test using three different soils from Germany. J. Plant Nutr. Soil Sci. 2018, 181, 894–904. [Google Scholar] [CrossRef]
- Fomin, D.; Timofeeva, M.; Ovchinnikova, O.; Valdes-Korovkin, I.; Holub, A.; Yudina, A. Energy-based indicators of soil structure by automatic dry sieving. Soil Tillage Res. 2021, 214, 105183. [Google Scholar] [CrossRef]
- Zhelezova, A.D.; Kutovaya, O.V.; Dmitrenko, V.N.; Tkhakhahova, A.K.; Khohlov, S.F. Estimation of DNA quantity in different groups of microorganisms within genetic horizons of the dark-gray soil. Dokuchaev Soil Bull. 2015, 78, 87–98. [Google Scholar] [CrossRef]
- Artemyeva, Z.S.; Kogut, B.M. The effect of tillage on organic carbon stabilization in microaggregates in different climatic zones of European Russia. Agriculture 2016, 6, 63. [Google Scholar] [CrossRef]
- Soil Survey Staff. Soil Taxonomy: A Basic System of Soil Classification for Making and Interpreting Soil Surveys, 2nd ed.; Natural Resources Conservation Service; U.S. Department of Agriculture Handbook; United States Deptartment of Agriculture, Naturel Resources Conservation Service: Portland, OR, USA, 1999.
- Marx, M.C.; Wood, M.; Jarvis, S.C. A microplate fluorimetric assay for the study of enzyme diversity in soils. Soil Biol. Biochem. 2001, 33, 1633–1640. [Google Scholar] [CrossRef]
- Razavi, B.S.; Blagodatskaya, E.; Kuzyakov, Y. Nonlinear temperature sensitivity of enzyme kinetics explains canceling effect—A case study on loamy haplic Luvisol. Front. Microbiol. 2015, 6, 1126. [Google Scholar] [CrossRef] [PubMed]
- Cartwright, J.; Dzantor, E.K.; Momen, B. Soil microbial community profiles and functional diversity in limestone cedar glades. Catena 2016, 147, 216–224. [Google Scholar] [CrossRef]
- Wu, Z.; Li, J.; Zheng, J.; Liu, J.; Liu, S.; Lin, W.; Wu, C. Soil microbial community structure and catabolic activity are significantly degenerated in successive rotations of Chinese fir plantations. Sci. Rep. 2017, 7, 6691. [Google Scholar] [CrossRef]
- Vorobyova, E.; Soina, V.; Gorlenko, M.; Minkovskaya, N.; Zalinova, N.; Mamukelashvili, A.; Gilichinsky, D.; Rivkina, E.; Vishnivetskaya, T. The deep cold biosphere: Facts and hypothesis. FEMS Microbiol. Rev. 1997, 20, 277–290. [Google Scholar] [CrossRef]
- Cheptsov, V.S.; Vorobyova, E.A.; Manucharova, N.A.; Gorlenko, M.V.; Pavlov, A.K.; Vdovina, M.A.; Lomasov, V.N.; Bulat, S.A. 100 kGy gamma-affected microbial communities within the ancient Arctic permafrost under simulated Martian conditions. Extremophiles 2017, 21, 1057–1067. [Google Scholar] [CrossRef]
- Christensen, B.T. Carbon and nitrogen in particle size fractions isolated from Danish arable soils by ultrasonic dispersion and gravity-sedimentation. Acta Agric. Scand. 1985, 35, 175–187. [Google Scholar] [CrossRef]
- Gregorich, E.G.; Voroney, R.P.; Kachanoski, R.G. Ultrasonic dispersion of aggregates: Distribution of organic matter in size fractions. Can. J. Soil Sci. 1988, 68, 395–403. [Google Scholar]
- Neumann, D.; Heuer, A.; Hemkemeyer, M.; Martens, R.; Tebbe, C.C. Response of microbial communities to long-term fertilization depends on their microhabitat. FEMS Microbiol. Ecol. 2013, 86, 71–84. [Google Scholar] [CrossRef]
- Hemkemeyer, M.; Christensen, B.T.; Martens, R.; Tebbe, C.C. Soil particle size fractions harbour distinct microbial communities and differ in potential for microbial mineralisation of organic pollutants. Soil Biol. Biochem. 2015, 90, 255–265. [Google Scholar] [CrossRef]
- Hemkemeyer, M.; Dohrmann, A.B.; Christensen, B.T.; Tebbe, C.C. Bacterial preferences for specific soil particle size fractions revealed by community analyses. Front. Microbiol. 2018, 9, 149. [Google Scholar] [CrossRef]
- Kandeler, E.; Tscherko, D.; Bruce, K.D.; Stemmer, M.; Hobbs, P.J.; Bardgett, R.D.; Amelung, W. Structure and function of the soil microbial community in microhabitats of a heavy metal polluted soil. Biol. Fertil. Soils 2000, 32, 390–400. [Google Scholar] [CrossRef]
- Gerzabek, M.H.; Haberhauer, G.; Kandeler, E.; Sessitsch, A.; Kirchmann, H. Response of organic matter pools and enzyme activities in particle size fractions to organic amendments in a long-term field experiment. In Developments in Soil Science; Elsevier: Amsterdam, The Netherlands, 2002; Volume 28, pp. 329–344. [Google Scholar]
- Semenov, V.M.; Lebedeva, T.N.; Pautova, N.B. Particulate organic matter in noncultivated and arable soils. Eurasian Soil Sci. 2019, 52, 396–404. [Google Scholar] [CrossRef]
- Trasar-Cepeda, C.; Leirós, M.C.; Gil-Sotres, F. Hydrolytic enzyme activities in agricultural and forest soils. Some implications for their use as indicators of soil quality. Soil Quality. Soil Biol. Biochem. 2008, 40, 2146–2155. [Google Scholar] [CrossRef]
- Six, J.; Bossuyt, H.; Degryze, S.; Denef, K. A history of research on the link between (micro) aggregates, soil biota, and soil organic matter dynamics. Soil Tillage Res. 2004, 79, 7–31. [Google Scholar] [CrossRef]



| Enzymes Tested | Forest | Arable |
|---|---|---|
| β-glucosidase | 137.70 ± 36.57 a | 85.61 ± 18.80 b |
| Cellobiohydrolase | 21.97 ± 2.17 a | 18.09 ± 1.99 b |
| Chitinase | 103.13 ± 88.49 a | 44.49 ± 8.88 a |
| Xylanase | 23.62 ± 10.85 a | 16.18 ± 1.23 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yudina, A.; Ovchinnikova, O.; Cheptsov, V.; Fomin, D. Localization of C Cycle Enzymes in Arable and Forest Phaeozems within Levels of Soil Microstructure. Microorganisms 2023, 11, 1343. https://doi.org/10.3390/microorganisms11051343
Yudina A, Ovchinnikova O, Cheptsov V, Fomin D. Localization of C Cycle Enzymes in Arable and Forest Phaeozems within Levels of Soil Microstructure. Microorganisms. 2023; 11(5):1343. https://doi.org/10.3390/microorganisms11051343
Chicago/Turabian StyleYudina, Anna, Olga Ovchinnikova, Vladimir Cheptsov, and Dmitry Fomin. 2023. "Localization of C Cycle Enzymes in Arable and Forest Phaeozems within Levels of Soil Microstructure" Microorganisms 11, no. 5: 1343. https://doi.org/10.3390/microorganisms11051343