
Identification of Fungicide Combinations Targeting Plasmopara viticola and Botrytis cinerea Fungicide Resistance Using Machine Learning


Abstract
:1. Introduction
Literature Review
2. Materials and Methods
2.1. Protein Structure and Ligand Structure Preparation
2.2. Molecular Docking
2.3. AutoQSAR Model Analysis
3. Results
3.1. Building of the Homology Models for Plasmopara viticola Cytochrome b
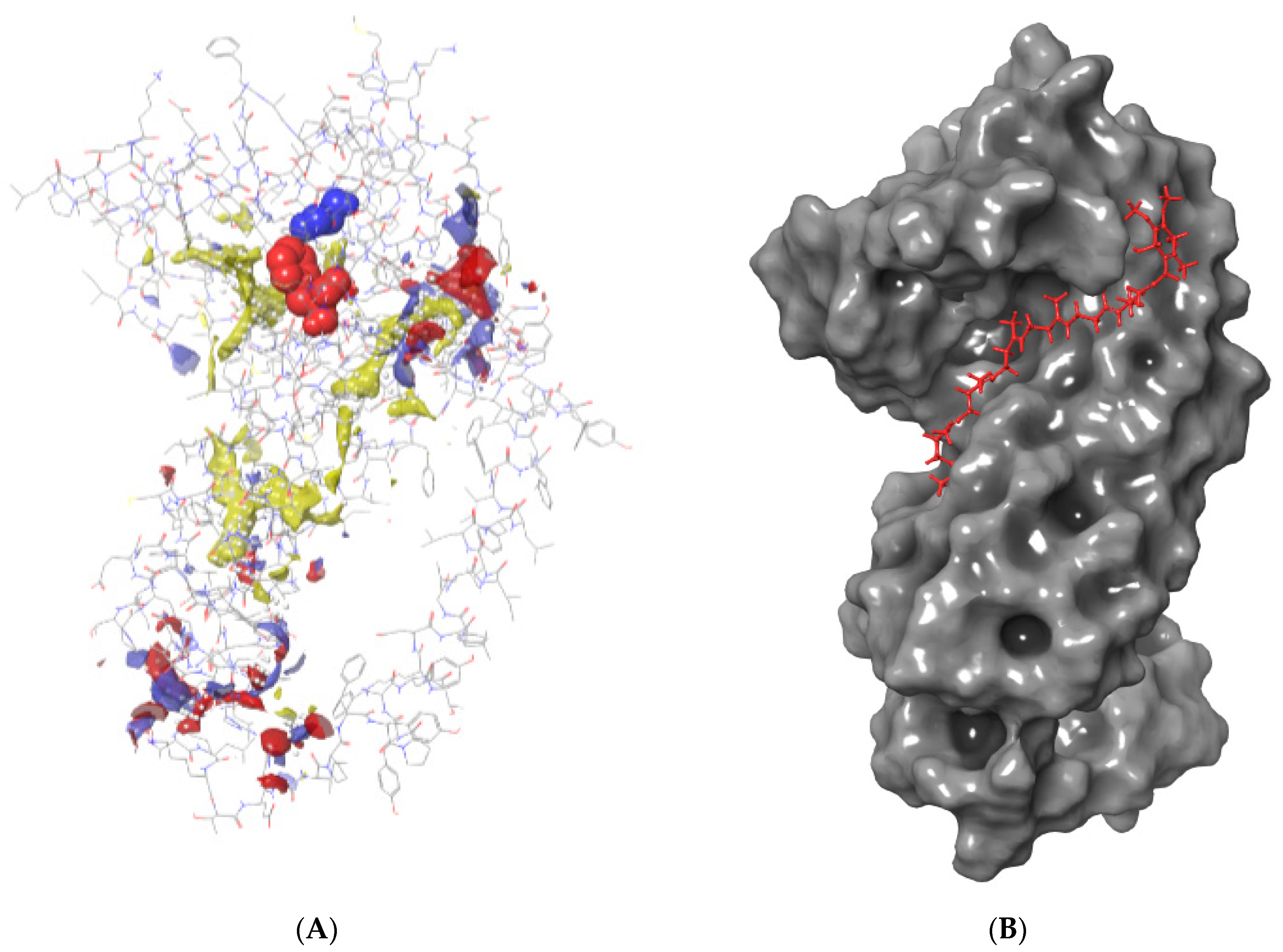
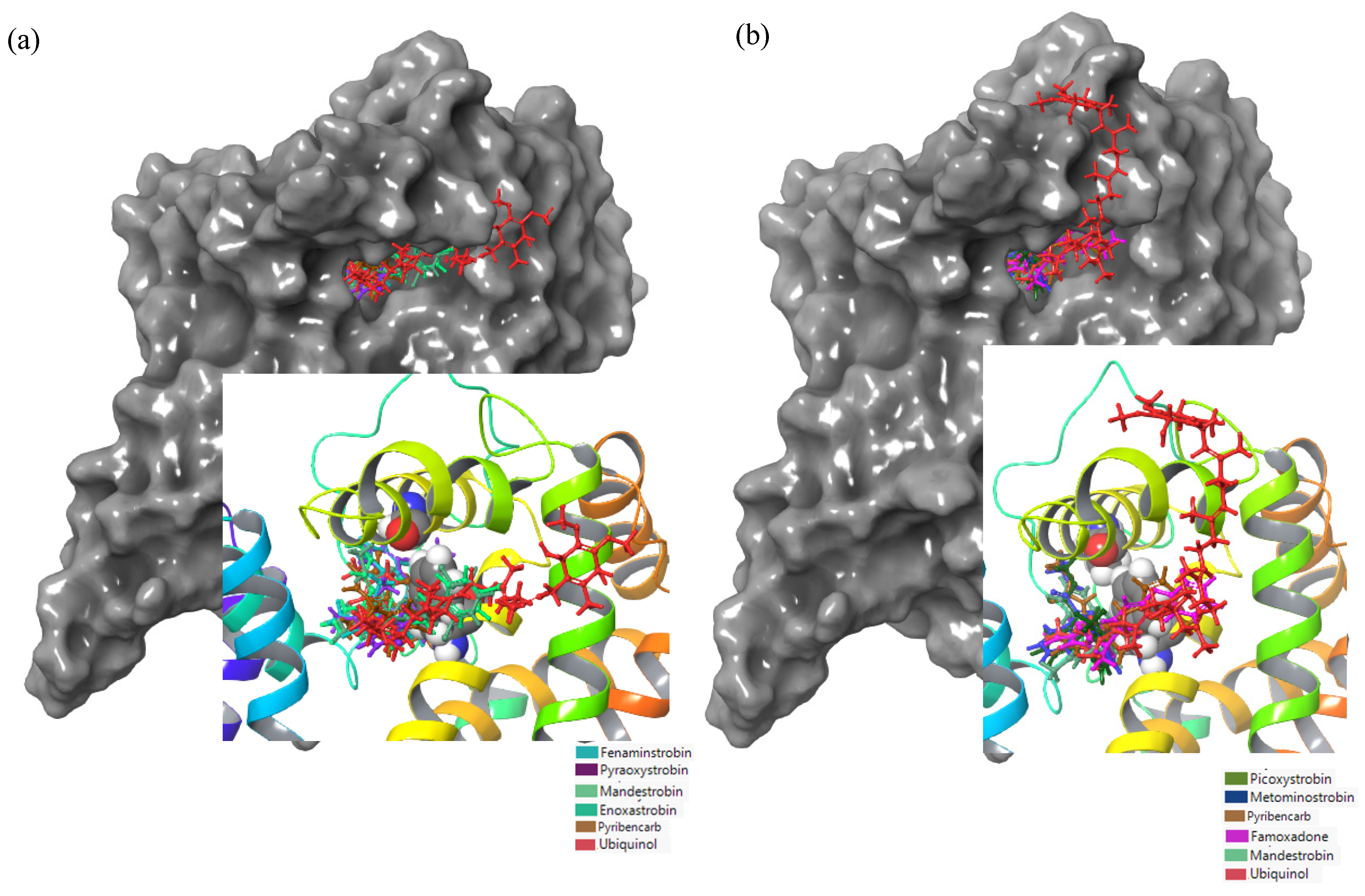
3.2. Identification of the Active Site for Plasmopara viticola
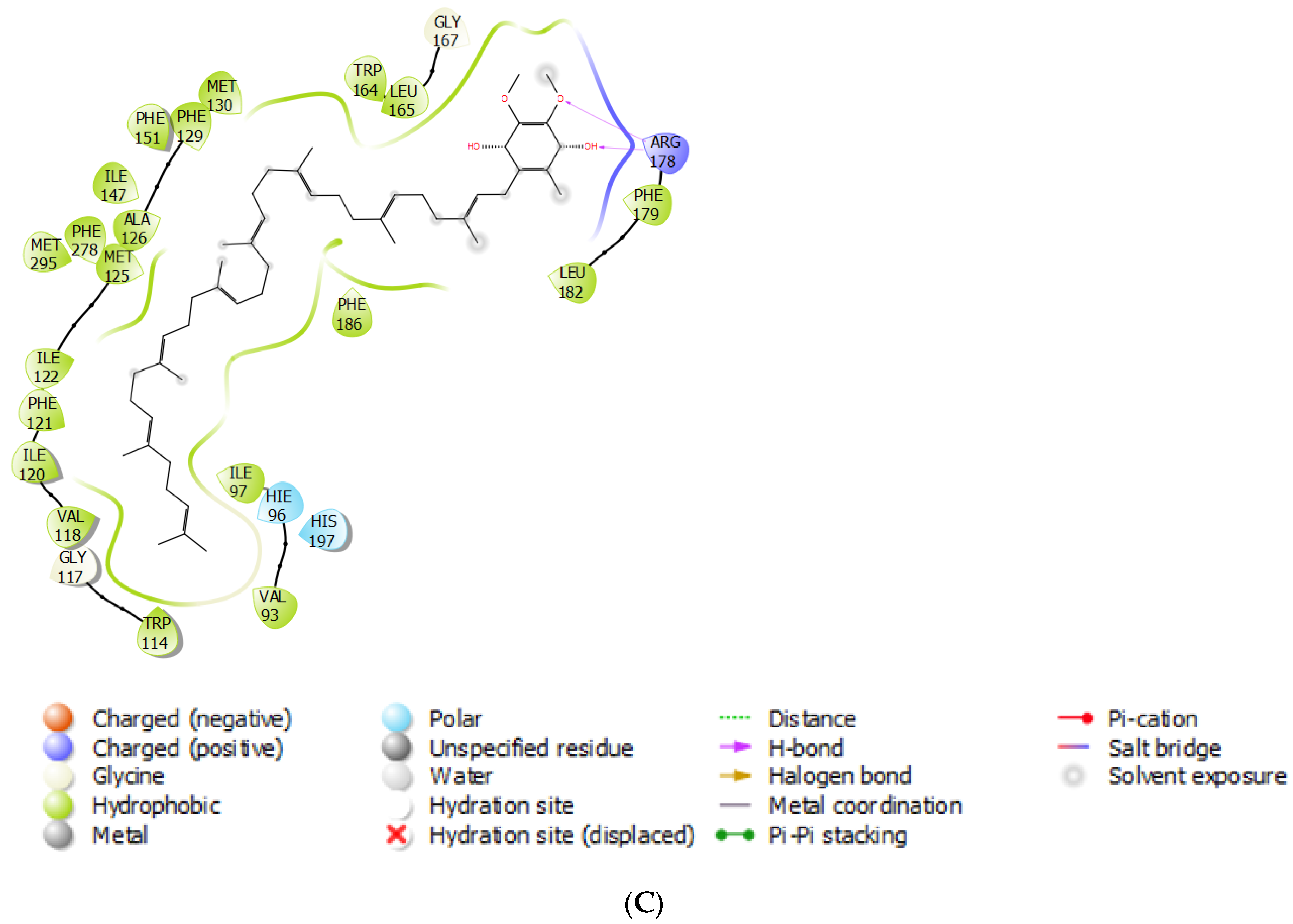
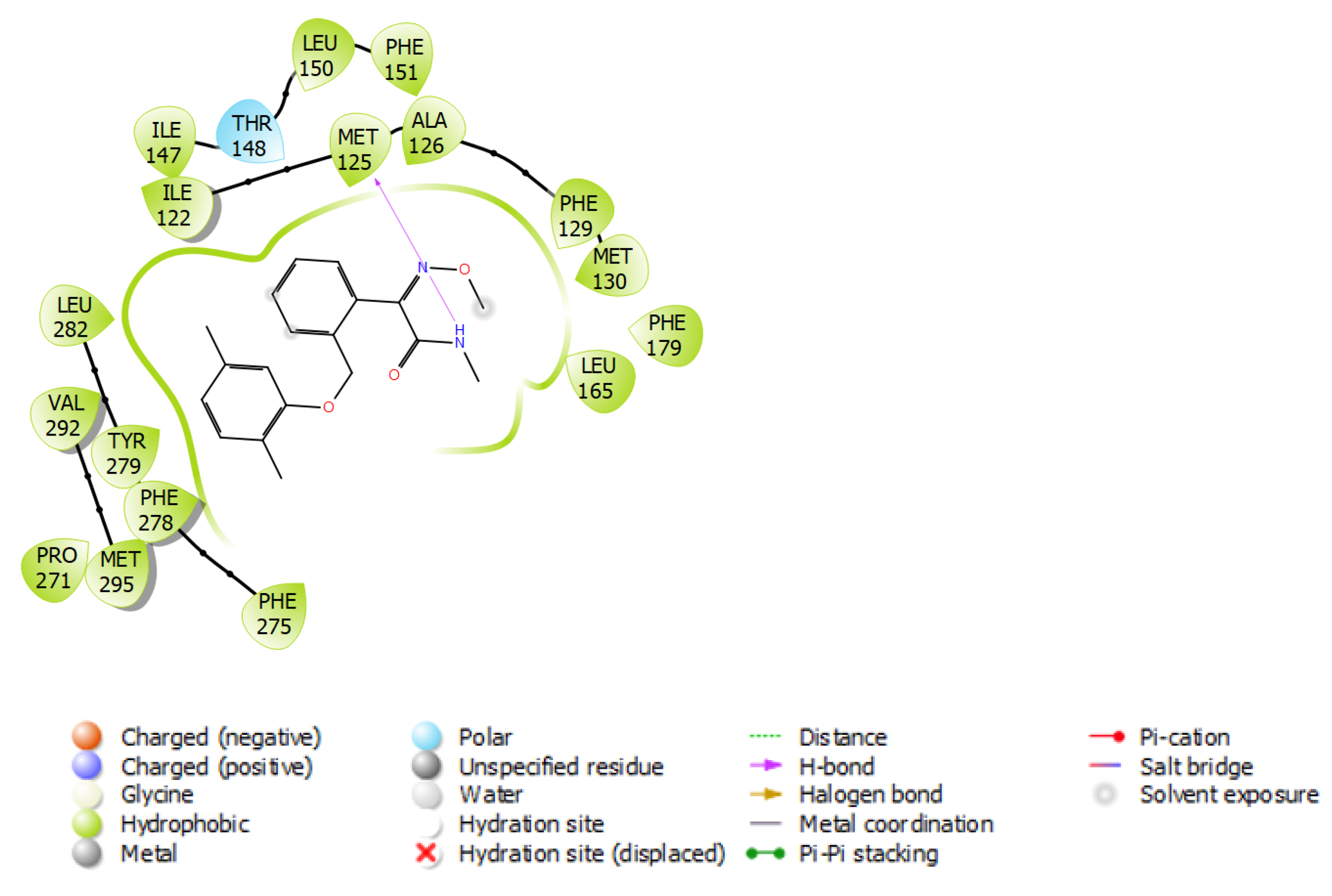
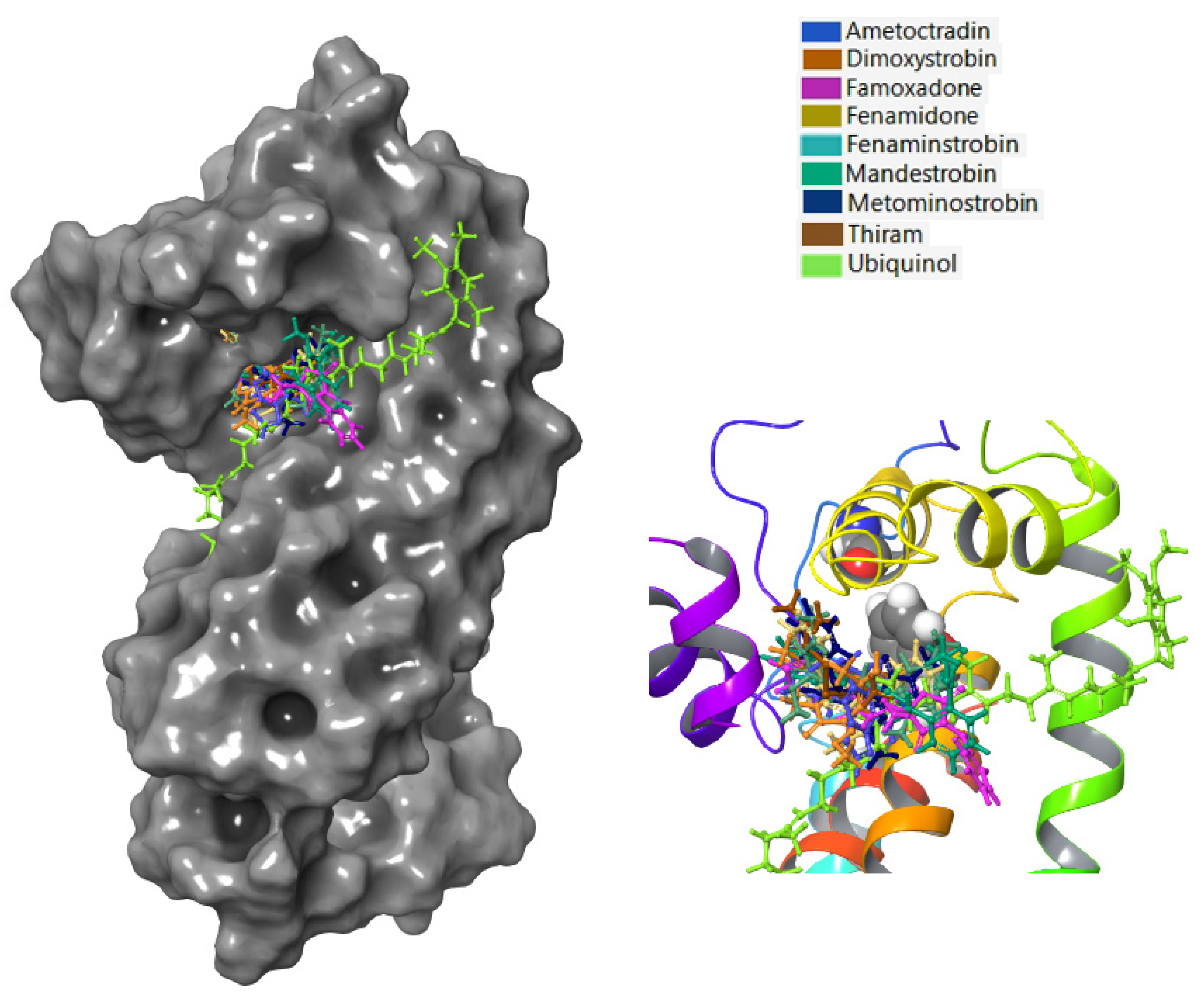
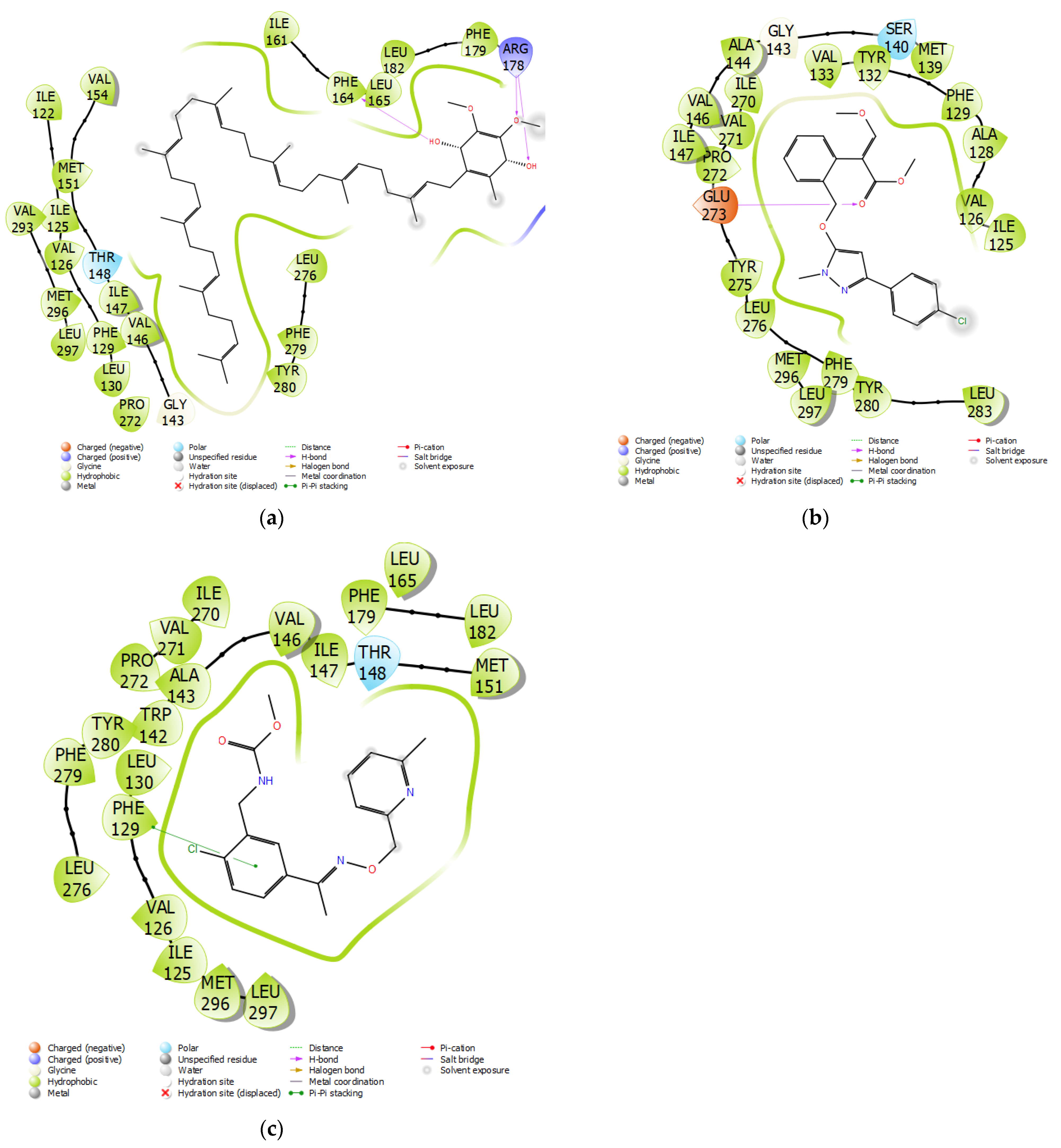
3.3. Fungicide Binding Behavior on Plasmopara viticola Cytochrome b
3.4. Mutation-Specific Observations
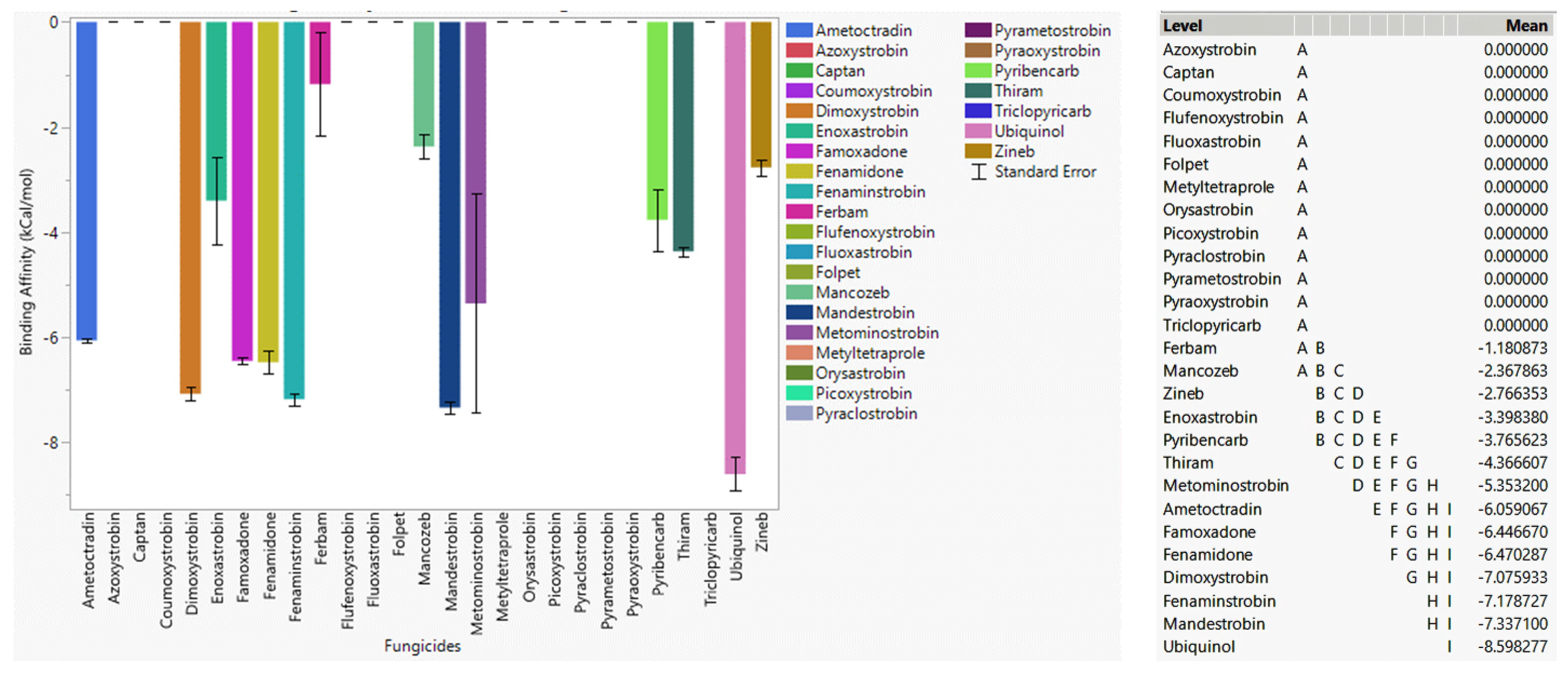
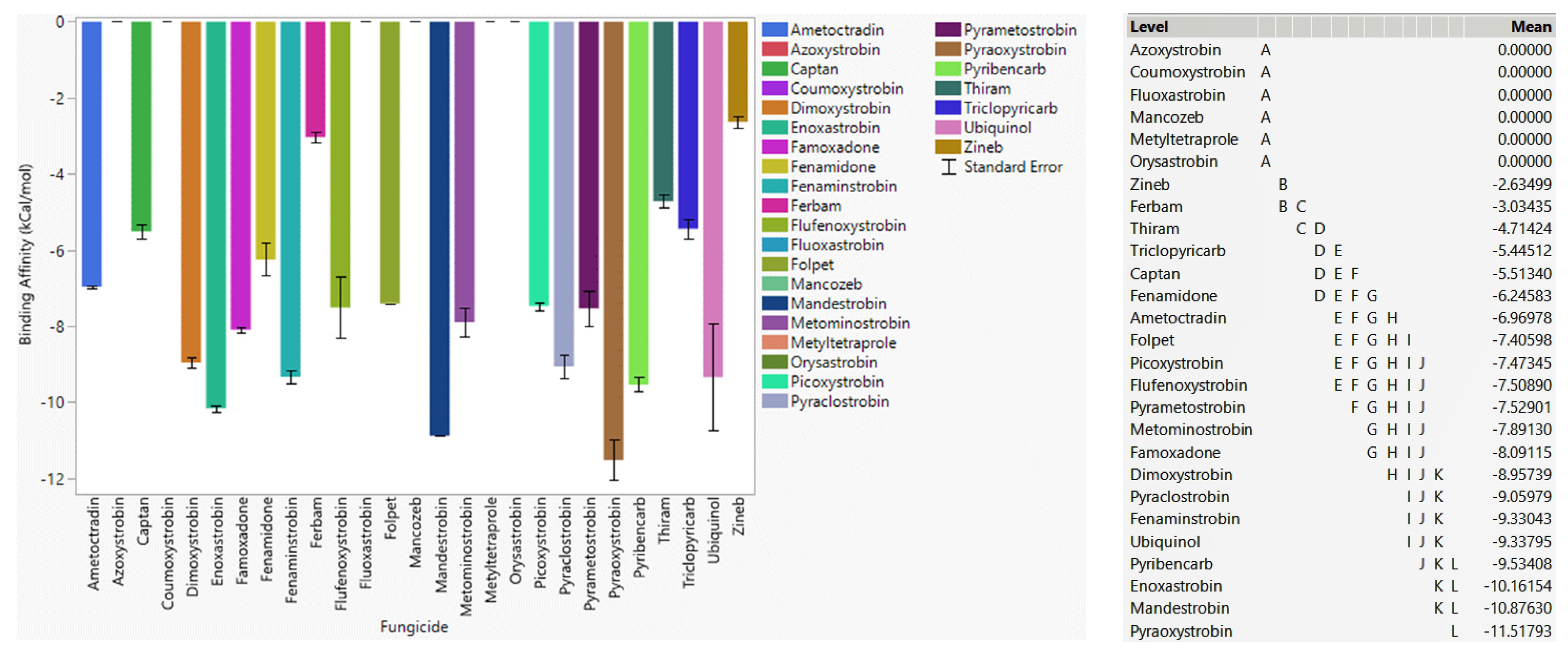
3.4.1. Fungicide Recommendations for WT
3.4.2. Fungicide Recommendations for the G143A Mutation
3.5. Fungicide Binding Behavior on Botrytis cinerea Cytochrome b
3.6. Mutation-Specific Observations
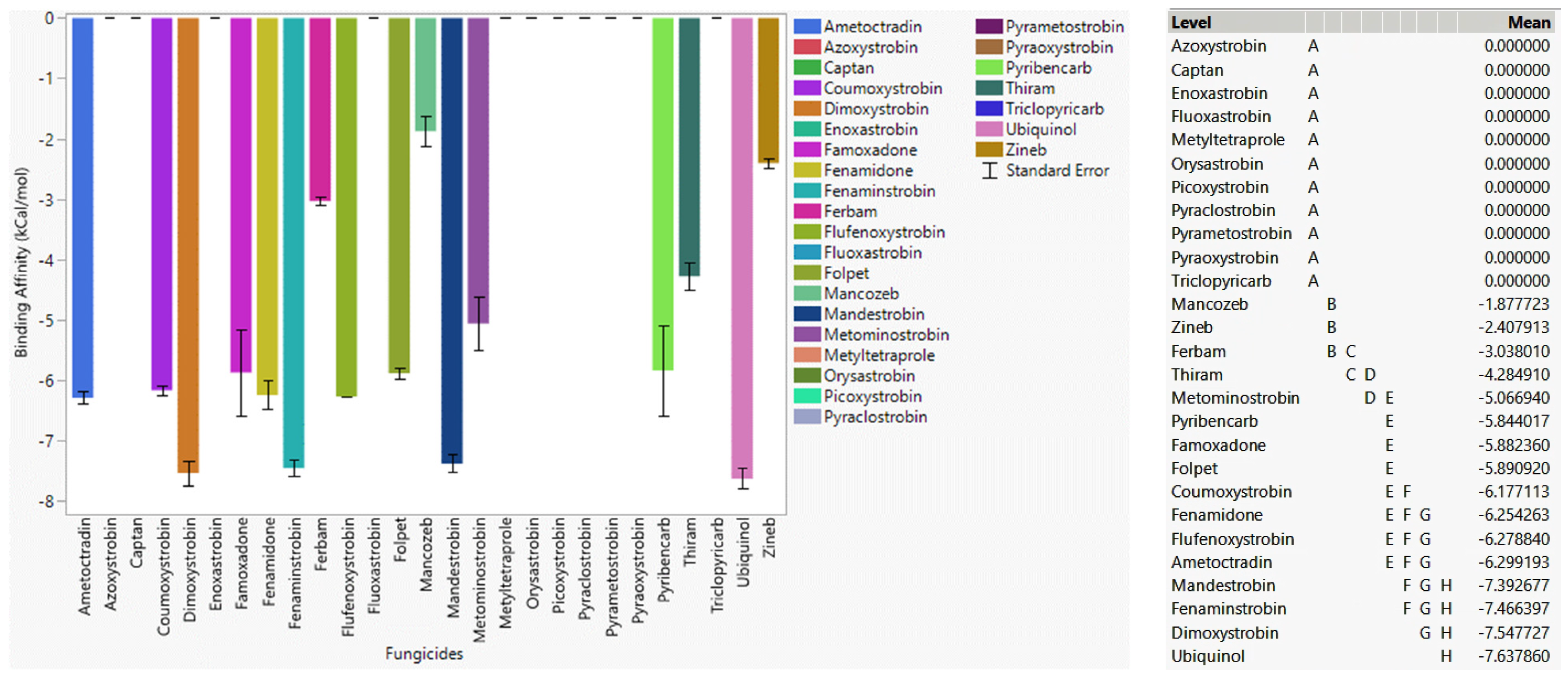
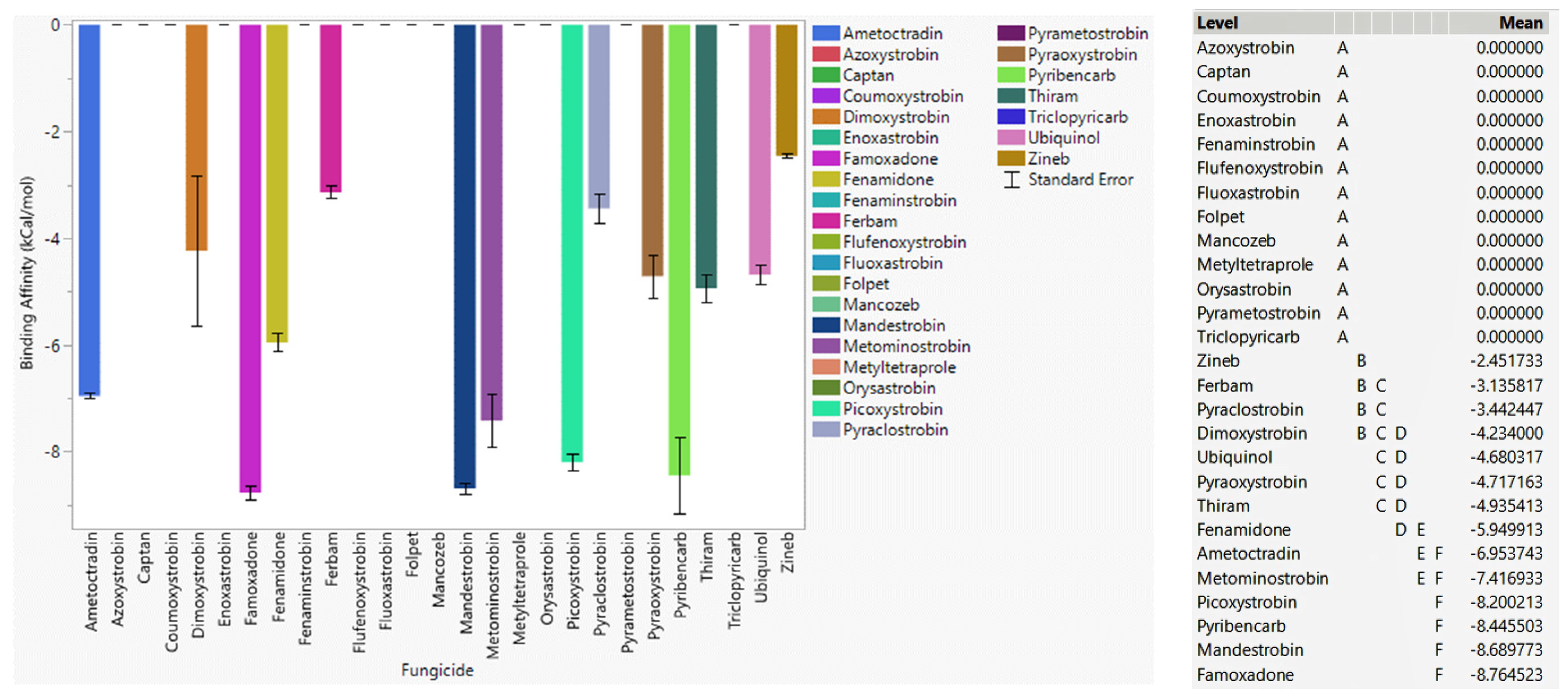
3.6.1. Fungicide Recommendations for WT
3.6.2. Fungicide Recommendations for the G143A Mutation
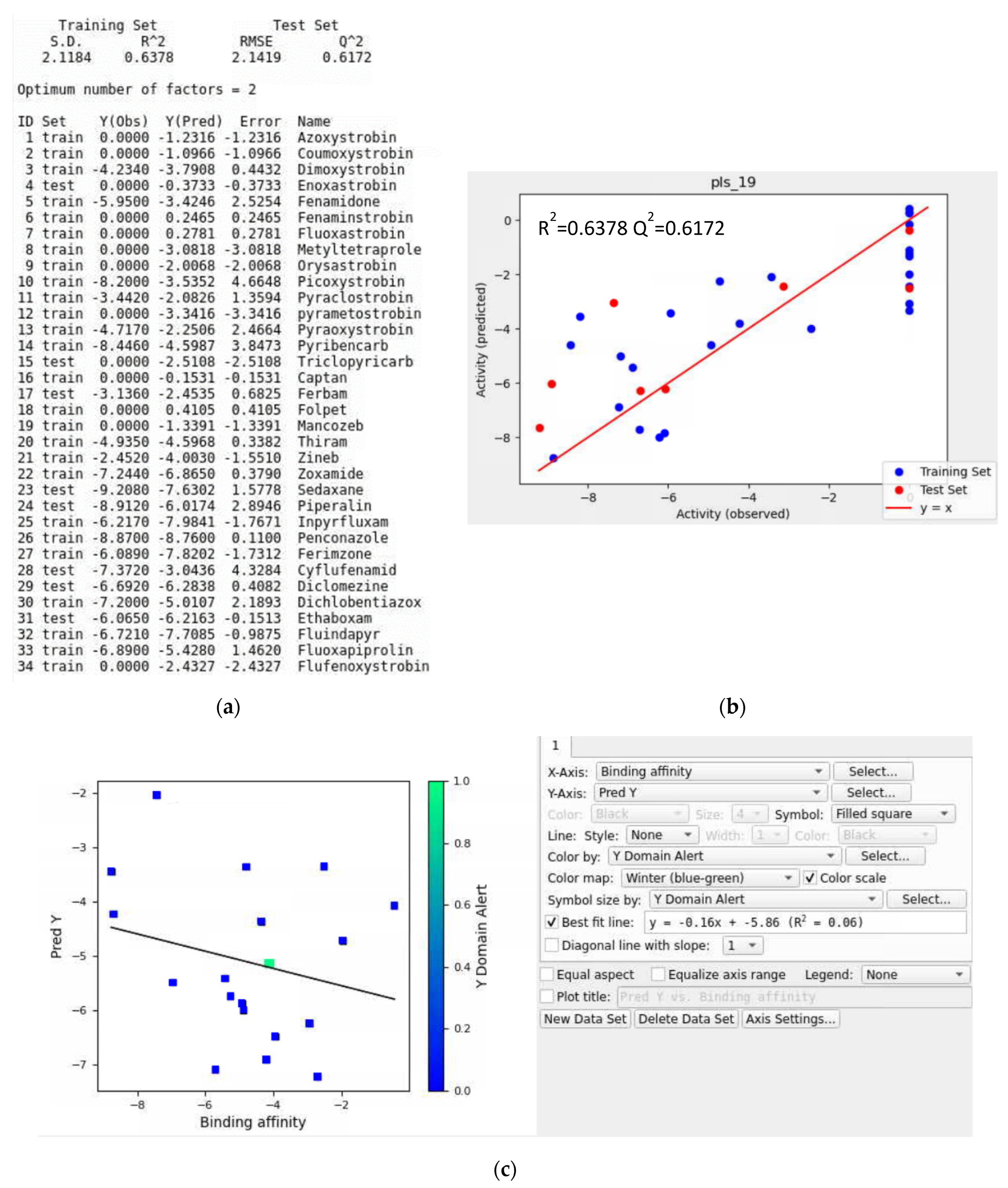
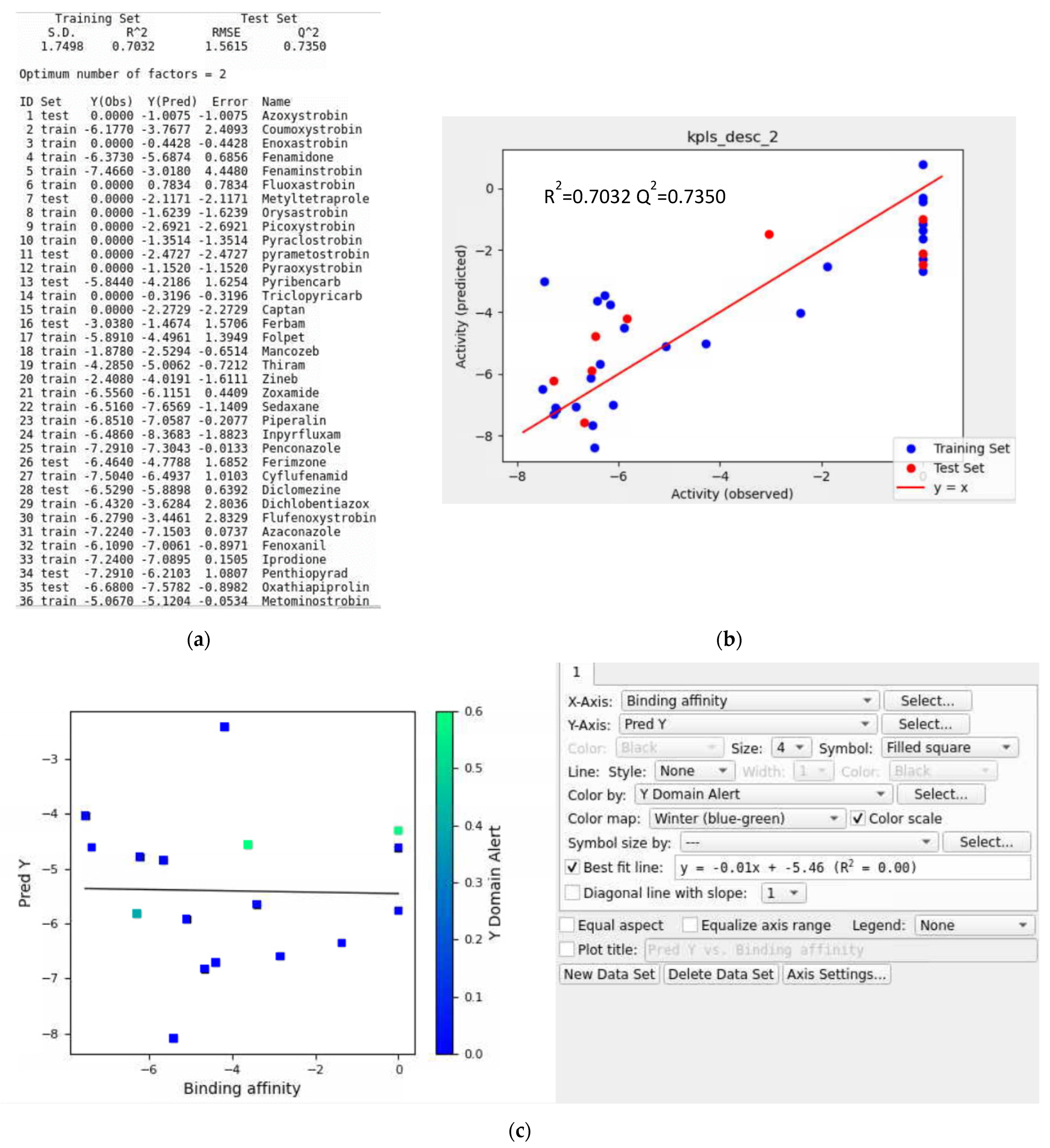
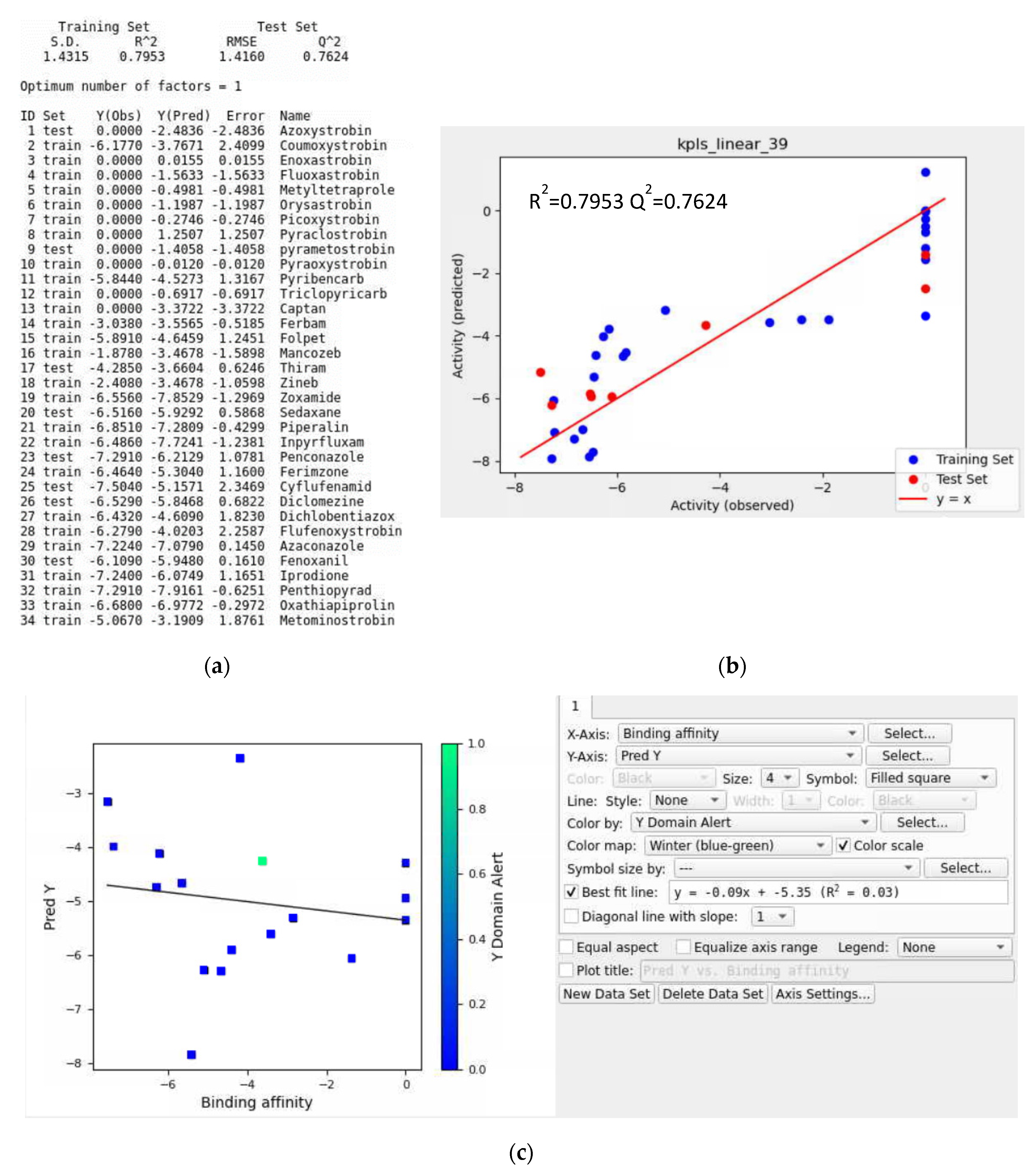
3.7. AutoQSAR Model Evaluation
Application of AutoQSR to Predict Fungicides for Botrytis cinerea
- Training Data without a Validation Set
- Iteration #1
- Iteration #2
- Iteration #3
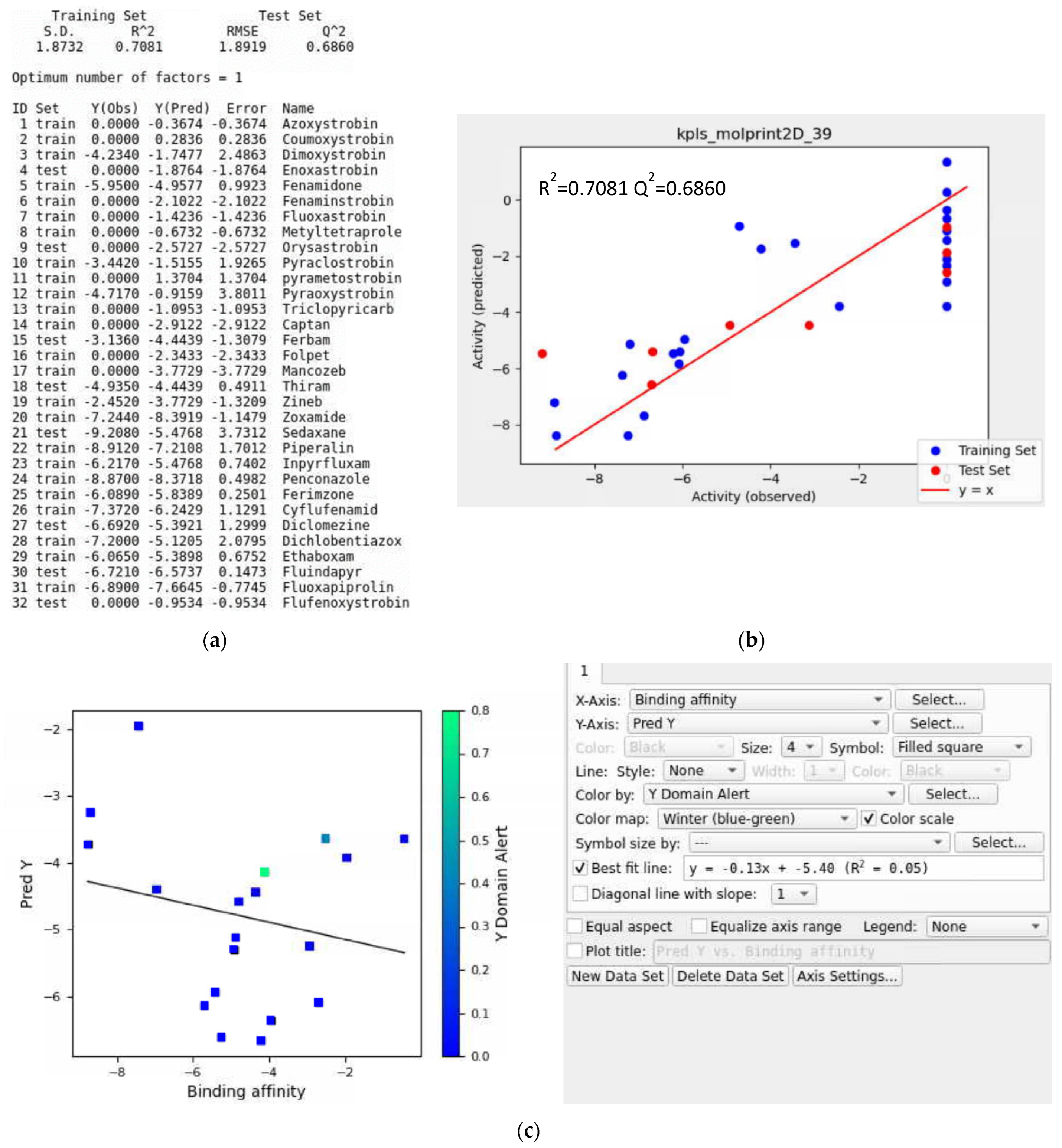
3.8. Training Data with a Validation Set
- Iteration #1
- Iteration #2
- Iteration #3
Application of AutoQSR to Predict Fungicides for Plasmopara viticola
- Training Data without a Validation Set
- Iteration #1
- Iteration #2
- Iteration #3
- Training Data with a Validation Set
- Iteration #1
- Iteration #2
- Iteration #3
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- FAO. FAOSTAT Statistical Database; Food and Agriculture Organization of the United Nations: Roma, Italy, 2020.
- Kramer, J.; Simnitt, S.; Calvin, L. Fruit and Tree Nuts Outlook: September 2021. US Dept. Agr. Econ. Res. Serv. 2021, 2021, 37. [Google Scholar]
- Murray, R.E.; Candan, A.P.; Vazquez, D.E. Manual de Poscosecha de Frutas: Manejo Integrado de Patógenos; Ediciones INTA: Buenos Aires, Argentina, 2019. [Google Scholar]
- Kassemeyer, H.-H. Fungi of grapes. In Biology of Microorganisms on Grapes, in Must and in Wine; Springer: Berlin/Heidelberg, Germany, 2017; pp. 103–132. [Google Scholar]
- Caffi, T.; Rossi, V.; Carisse, O. Evaluation of a dynamic model for primary infections caused by Plasmopara viticola on grapevine in Quebec. Plant Health Prog. 2011, 12, 22. [Google Scholar] [CrossRef]
- Agrios, G.N. Plant Pathology; Elsevier: Amsterdam, The Netherlands, 2005. [Google Scholar]
- Mezei, I.; Lukić, M.; Berbakov, L.; Pavković, B.; Radovanović, B. Grapevine Downy Mildew Warning System Based on NB-IoT and Energy Harvesting Technology. Electronics 2022, 11, 356. [Google Scholar] [CrossRef]
- Chen, W.-J.; Delmotte, F.; Cervera, S.R.; Douence, L.; Greif, C.; Corio-Costet, M.-F. At least two origins of fungicide resistance in grapevine downy mildew populations. Appl. Environ. Microbiol. 2007, 73, 5162–5172. [Google Scholar] [CrossRef] [PubMed]
- Dean, R.; Van Kan, J.A.; Pretorius, Z.A.; Hammond-Kosack, K.E.; Di Pietro, A.; Spanu, P.D.; Rudd, J.J.; Dickman, M.; Kahmann, R.; Ellis, J. The Top 10 fungal pathogens in molecular plant pathology. Mol. Plant Pathol. 2012, 13, 414–430. [Google Scholar] [CrossRef] [PubMed]
- Elmer, P.; Reglinski, T. Biosuppression of Botrytis cinerea in grapes. Plant Pathol. 2006, 55, 155–177. [Google Scholar] [CrossRef]
- Foster, A.J. Identification of fungicide targets in pathogenic fungi. Physiol. Genet. 2018, 15, 277–296. [Google Scholar]
- Bartlett, D.W.; Clough, J.M.; Godwin, J.R.; Hall, A.A.; Hamer, M.; Parr-Dobrzanski, B. The strobilurin fungicides. Pest Manag. Sci. Former. Pestic. Sci. 2002, 58, 649–662. [Google Scholar] [CrossRef]
- Hahn, M. The rising threat of fungicide resistance in plant pathogenic fungi: Botrytis as a case study. J. Chem. Biol. 2014, 7, 133–141. [Google Scholar] [CrossRef]
- Samuel, S.; Papayiannis, L.C.; Leroch, M.; Veloukas, T.; Hahn, M.; Karaoglanidis, G.S. Evaluation of the incidence of the G143A mutation and cytb intron presence in the cytochrome bc-1 gene conferring QoI resistance in Botrytis cinerea populations from several hosts. Pest Manag. Sci. 2011, 67, 1029–1036. [Google Scholar] [CrossRef]
- Hollomon, D.W. Fungicide resistance: Facing the challenge—A review. Plant Prot. Sci. 2015, 51, 170–176. [Google Scholar] [CrossRef]
- Sierotzki, H.; Kraus, N.; Assemat, P.; Stanger, C.; Cleere, S.; Windass, J.; Gisi, U. Evolution of resistance to QoI fungicides in Plasmopara viticola populations in Europe. In Proceedings of the Modern Fungicides and Antifungal Compounds IV: 14th International Reinhardsbrunn Symposium, Friedrichroda, Germany, 25–29 April 2004; pp. 73–80. [Google Scholar]
- Lijuan, X.; Shangfei, Q.; Zhengchuan, S.; Huiling, Y. Field efficacy of the combination of famoxadone and metalaxyl-M against Plasmopara viticola and the residue dynamics of the two fungicides in grape. Chin. J. Pestic. Sci. 2022, 24, 326–331. [Google Scholar]
- Kabade, S.H.; Pawar, S.B.; Edoliya, R.; Saha, S. Performance and Phytotoxicity assessment of Mancozeb 40%+ Azoxystrobin 7% OS against downy mildew of grapes in Maharashtra, India. J. Mycopathol. Res. 2022, 60, 581–585. [Google Scholar] [CrossRef]
- Liu, F.; Li, J.; Li, H.; Wang, K.; Meng, L.; Li, B.; Mu, W.; Liu, F. Evaluation on Synergistic Activity and Resistance Development of the Mixture of Iprodione and Fluopyram Against Botrytis Cinerea. Available online: https://ssrn.com/abstract=4118573 (accessed on 1 April 2023).
- Sagar, N.; Jamadar, M.; Shalini, N.; Jagginavar, S.; Pattar, P. Bioefficacy of different fungicides against Plasmopara viticola and Erysiphe necator of grapes. Pharma Innov. J. 2022, 11, 2512–2518. [Google Scholar]
- Malandrakis, A.A.; Kavroulakis, N.; Chrysikopoulos, C.V. Synergy between Cu-NPs and fungicides against Botrytis cinerea. Sci. Total Environ. 2020, 703, 135557. [Google Scholar] [CrossRef]
- Arnold, K.; Bordoli, L.; Kopp, J.; Schwede, T. The SWISS-MODEL workspace: A web-based environment for protein structure homology modelling. Bioinformatics 2006, 22, 195–201. [Google Scholar] [CrossRef]
- Maldonado, M.; Guo, F.; Letts, J.A. Atomic structures of respiratory complex III2, complex IV, and supercomplex III2-IV from vascular plants. Elife 2021, 10, e62047. [Google Scholar] [CrossRef]
- Waterhouse, A.; Bertoni, M.; Bienert, S.; Studer, G.; Tauriello, G.; Gumienny, R.; Heer, F.T.; de Beer, T.A.P.; Rempfer, C.; Bordoli, L. SWISS-MODEL: Homology modelling of protein structures and complexes. Nucleic Acids Res. 2018, 46, W296–W303. [Google Scholar] [CrossRef]
- Colovos, C.; Yeates, T.O. Verification of protein structures: Patterns of nonbonded atomic interactions. Protein Sci. 1993, 2, 1511–1519. [Google Scholar] [CrossRef]
- Pontius, J.; Richelle, J.; Wodak, S.J. Deviations from standard atomic volumes as a quality measure for protein crystal structures. J. Mol. Biol. 1996, 264, 121–136. [Google Scholar] [CrossRef]
- Irwin, J. ZINC15. docking. org: Over 1.5 billion compounds you can search and buy; 550 million lead-like you can dock. In Proceedings of the Abstracts of Papers of the American Chemical Society, Washington, DC, USA, 15–16 September 2019; Volume 257. [Google Scholar]
- Kim, S.; Chen, J.; Cheng, T.; Gindulyte, A.; He, J.; He, S.; Li, Q.; Shoemaker, B.A.; Thiessen, P.A.; Yu, B. PubChem in 2021: New data content and improved web interfaces. Nucleic Acids Res. 2021, 49, D1388–D1395. [Google Scholar] [CrossRef] [PubMed]
- Jorgensen, W.L.; Tirado-Rives, J. The OPLS [optimized potentials for liquid simulations] potential functions for proteins, energy minimizations for crystals of cyclic peptides and crambin. J. Am. Chem. Soc. 1988, 110, 1657–1666. [Google Scholar] [CrossRef] [PubMed]
- Release, S. AutoQSAR.; Schrödinger, LLC: New York, NY, USA, 2022. [Google Scholar]
- Laskowski, R.A.; Rullmann, J.A.C.; MacArthur, M.W.; Kaptein, R.; Thornton, J.M. AQUA and PROCHECK-NMR: Programs for checking the quality of protein structures solved by NMR. J. Biomol. NMR 1996, 8, 477–486. [Google Scholar] [CrossRef] [PubMed]
- Williams, C.J.; Headd, J.J.; Moriarty, N.W.; Prisant, M.G.; Videau, L.L.; Deis, L.N.; Verma, V.; Keedy, D.A.; Hintze, B.J.; Chen, V.B. MolProbity: More and better reference data for improved all-atom structure validation. Protein Sci. 2018, 27, 293–315. [Google Scholar] [CrossRef]
- Grasso, V.; Palermo, S.; Sierotzki, H.; Garibaldi, A.; Gisi, U. Cytochrome b gene structure and consequences for resistance to Qo inhibitor fungicides in plant pathogens. Pest Manag. Sci. Former. Pestic. Sci. 2006, 62, 465–472. [Google Scholar] [CrossRef]
- Mounkoro, P.; Michel, T.; Benhachemi, R.; Surpateanu, G.; Iorga, B.I.; Fisher, N.; Meunier, B. Mitochondrial complex III Qi-site inhibitor resistance mutations found in laboratory selected mutants and field isolates. Pest Manag. Sci. 2019, 75, 2107–2114. [Google Scholar] [CrossRef]
- Wong, F.P.; Wilcox, W.F. Distribution of baseline sensitivities to azoxystrobin among isolates of Plasmopara viticola. Plant Dis. 2000, 84, 275–281. [Google Scholar] [CrossRef]
- Fisher, N.; Meunier, B.; Biagini, G.A. The cytochrome bc1 complex as an antipathogenic target. Febs Lett. 2020, 594, 2935–2952. [Google Scholar] [CrossRef]
- Wenz, T.; Covian, R.; Hellwig, P.; MacMillan, F.; Meunier, B.; Trumpower, B.L.; Hunte, C. Mutational analysis of cytochrome b at the ubiquinol oxidation site of yeast complex III. J. Biol. Chem. 2007, 282, 3977–3988. [Google Scholar] [CrossRef]















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Study | Results | Limitations |
|---|---|---|
| Field efficacy of the combination of famoxadone and metalaxyl-M against Plasmopara viticola and the residue dynamics of the two fungicides in grapevine [17]. | Formulation of 30% famoxadone with metalaxyl-M SC was effective against Plasmopara viticola [17]. | Only a few fungicides were tested in the experiment. |
| Performance and phytotoxicity assessment of mancozeb 40% + azoxystrobin 7% OS against downy mildew of grapes in Maharashtra, India [18]. | Formulation of 40% mancozeb with 7% azoxystrobin was effective against Plasmopara viticola [18]. | Only a limited number of fungicides were tested over a period of two years. |
| Evaluation of synergistic activity and resistance development of the mixture of iprodione and fluopyram against Botrytis Cinerea [19]. | Formulation of 80% iprodione with 20% fluopyram was effective against Botrytis cinerea [19]. | Only two fungicides were tested in this research. |
| Bioefficacy of different fungicides against Plasmopara viticola and Erysiphe necator of grapes [20]. | Formulations of 16.6% famoxadone with 22.1% cymoxanil, 10% famoxadone with 50% mancozeb, and 4.44% fluopicolide with 66.67% fosetyl-Al were recommended for Plasmopara viticola [20]. | Experiments were limited to existing fungicide combinations on the market. |
| Synergy between Cu-NPs and fungicides against Botrytis cinerea [21]. | Formulation of copper nanoparticles with fluazinam or thiophanate was effective against Botrytis cinerea [21]. | The study was limited to a select few fungicides, and the long-term efficacy is unknown. The development of effective new nanoparticles would require many years, and the process of producing copper nanoparticles would be challenging. |
| Fungicide | Resistance 1 | Fungicide Type 2 |
|---|---|---|
| Ubiqunol | NA | NA |
| Famoxadone | HR | QoI |
| Azoxystrobin | HR/R | QoI |
| Fenamidone | HR | QoI |
| Coumoxystrobin | HR | QoI |
| Flufenoxystrobin | HR | QoI |
| Enoxastrobin | HR | QoI |
| Pyraoxystrobin | HR | QoI |
| Picoxystrobin | HR | QoI |
| Metyltetraprole | HR | QoI |
| Fenaminstrobin | HR | QoI |
| Pyribencarb | HR | QoI |
| Dimoxystrobin | HR | QoI |
| Triclopyricarb | HR | QoI |
| Metominostrobin | HR | QoI |
| Pyrametostrobin | HR | QoI |
| Mandestrobin | HR | QoI |
| Fluoxastrobin | HR | QoI |
| Pyraclostrobin | HR | QoI |
| Orysastrobin | HR | QoI |
| Folpet | LR | PHT |
| Ferbam | LR | DTC |
| Captan | LR | PHT |
| Mancozeb | LR | DTC |
| Ametoctradin | HR/R | QoI |
| Thiram | LR | DTC |
| Zineb | LR | DTC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, J.; Fernando, S.D. Identification of Fungicide Combinations Targeting Plasmopara viticola and Botrytis cinerea Fungicide Resistance Using Machine Learning. Microorganisms 2023, 11, 1341. https://doi.org/10.3390/microorganisms11051341
Zhang J, Fernando SD. Identification of Fungicide Combinations Targeting Plasmopara viticola and Botrytis cinerea Fungicide Resistance Using Machine Learning. Microorganisms. 2023; 11(5):1341. https://doi.org/10.3390/microorganisms11051341
Chicago/Turabian StyleZhang, Junrui, and Sandun D. Fernando. 2023. "Identification of Fungicide Combinations Targeting Plasmopara viticola and Botrytis cinerea Fungicide Resistance Using Machine Learning" Microorganisms 11, no. 5: 1341. https://doi.org/10.3390/microorganisms11051341