
Antibiotic Susceptibility Profiling of Human Pathogenic Staphylococcus aureus Strains Using Whole Genome Sequencing and Genome-Scale Annotation Approaches

 , ,
, ,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation of Multidrug-Resistant S. aureus
2.2. Antimicrobial Susceptibility Testing
2.3. Molecular Typing of Multi-Drug-Resistant S. aureus
2.3.1. S. aureus DNA Extraction
2.3.2. Whole Genome Sequencing, Typing, and Prediction of Resistome and Virulome
2.3.3. Pan-Genome Analysis and Phylogenomic Tree
3. Results and Discussion
3.1. Antimicrobial Susceptibility of the Isolated S. aureus
3.2. Genome Composition and Genomic Variation
3.3. Prediction of Antimicrobial Resistance Mechanism in the Studied S. aureus
3.4. Plasmid-Mediated Antimicrobial Resistance Genes
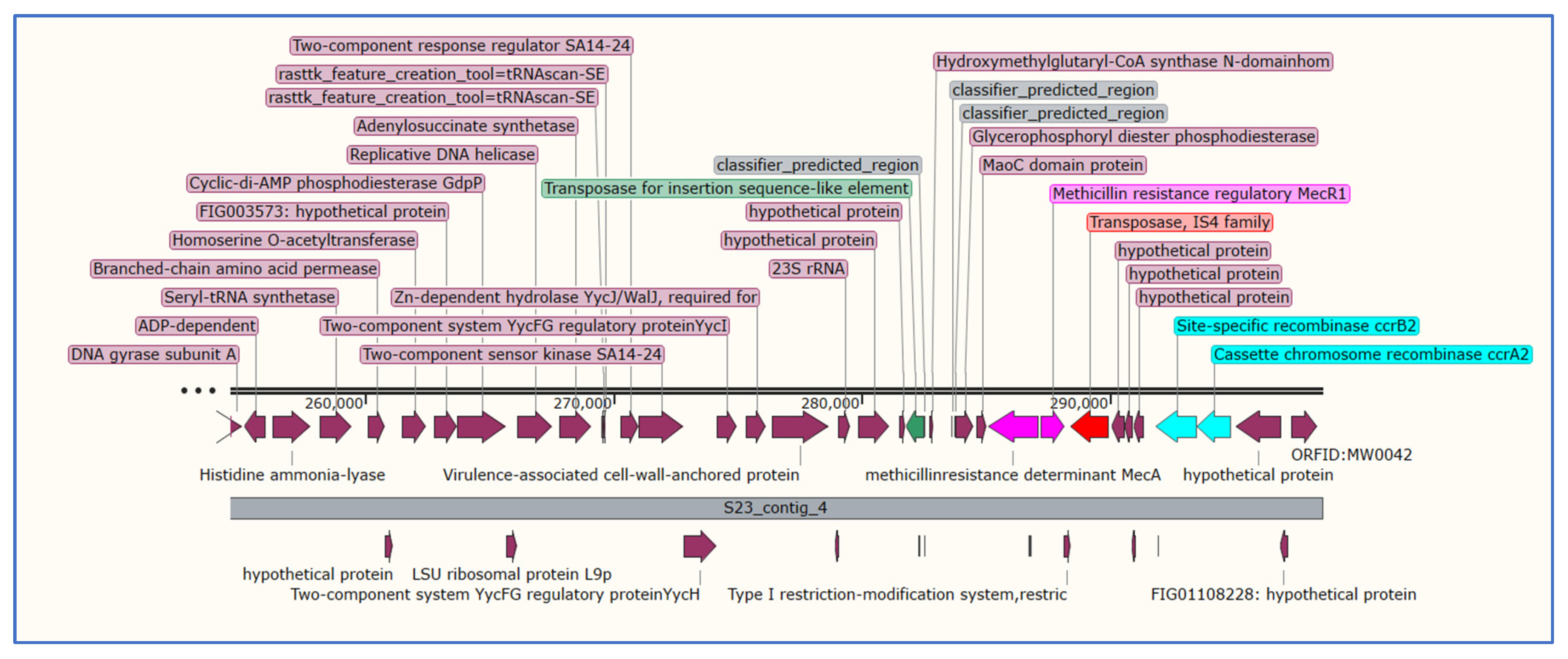
3.5. Analysis of Staphylococcal Chromosomal Cassettes mec (SCCmec)
3.6. Pan-Genome Analysis
3.7. Molecular Analysis of Virulence Factors in the Studied S. aureus
3.7.1. Detection of Panton–Valentine Leukocidin Gene
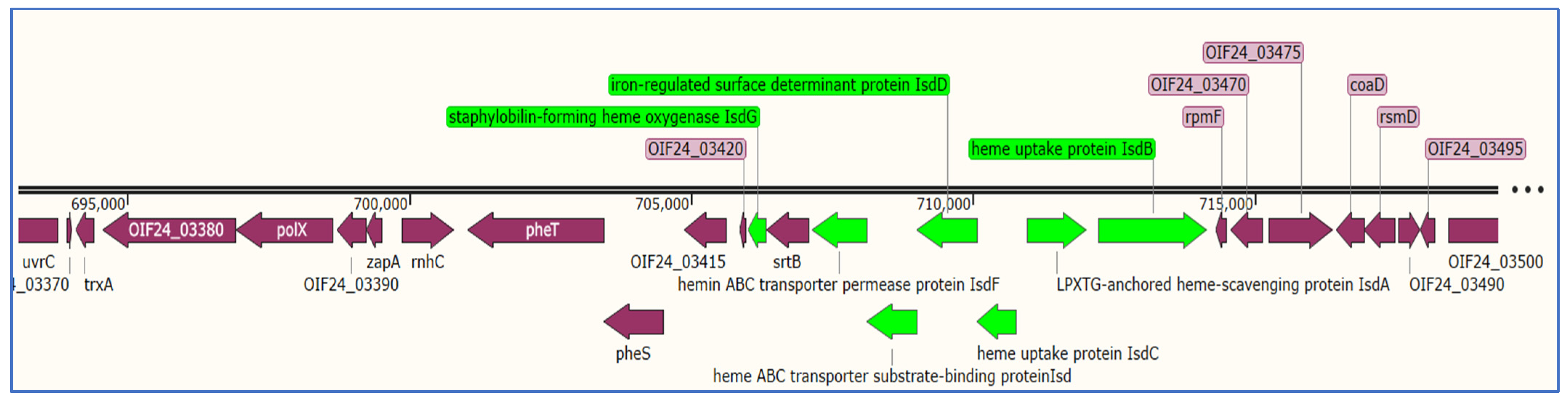
3.7.2. Detection of Genes Related to Iron Uptake System in S. aureus
3.7.3. Detection of Autolysin Genes in the Studied S. aureus Strains
3.7.4. Detection of Enterotoxin and Exotoxin in the Studied S. aureus
3.7.5. Detection of Toxic Shock Syndrome Toxin in S. aureus
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Samia, N.I.; Robicsek, A.; Heesterbeek, H.; Peterson, L.R. Methicillin-resistant Staphylococcus aureus nosocomial infection has a distinct epidemiological position and acts as a marker for overall hospital-acquired infection trends. Sci. Rep. 2022, 12, 17007. [Google Scholar] [CrossRef] [PubMed]
- Alghaithy, A.A.; Bilal, N.E.; Gedebou, M.; Weily, A.H. Nasal carriage and antibiotic resistance of Staphylococcus aureus isolates from hospital and non-hospital personnel in Abha, Saudi Arabia. Trans. R. Soc. Trop. Med. Hyg. 2000, 94, 504–547. [Google Scholar] [CrossRef]
- Bazaid, A.S.; Saeed, A.; Alrashidi, A.; Alrashidi, A.; Alshaghdali, K.; AHammam, S.; Alreshidi, T.; Alshammary, M.; Alarfaj, A.; Thallab, R.; et al. Antimicrobial Surveillance for Bacterial Uropathogens in Ha’il, Saudi Arabia: A Five-Year Multicenter Retrospective Study. Infect Drug Resist. 2021, 14, 1455–1465. [Google Scholar] [CrossRef]
- Ash, S.; Kennedy, L.E. Deep soft-tissue infections. In Schlossberg’s Clinical Infectious Disease; Oxford University Press: Oxford, UK, 2022; Volume 154. [Google Scholar]
- Bourget, M.; Pasquie, M.; Charbonneau, H.; Bonnet, E. Comparable clinical course between coagulase-negative staphylococcal and Staphylococcus aureus endocarditis. Infection 2022, 50, 483–490. [Google Scholar] [CrossRef]
- Moutaouakkil, K.; Abdellaoui, H.; Arhoune, B.; Atarraf, K.; El Fakir, S.; Yahyaoui, G.; Mahmoud, M.; Afifi, M.A.; Oumokhtar, B. Paediatric osteoarticular infections caused by Staphylococcus aureus producing panton–valentine leucocidin in morocco: Risk factors and clinical features. Afr. J. Paediatr. Surg. 2022, 19, 78. [Google Scholar] [PubMed]
- Zhang, F.; Wu, S.; Lei, T.; Wu, Q.; Zhang, J.; Huang, J.; Dai, J.; Chen, M.; Ding, Y.; Wang, J. Presence and characterization of methicillin-resistant Staphylococcus aureus co-carrying the multidrug resistance genes cfr and lsa (E) in retail food in China. Int. J. Food Microbiol. 2022, 363, 109512. [Google Scholar] [CrossRef]
- Guo, H.; Tong, Y.; Cheng, J.; Abbas, Z.; Li, Z.; Wang, J.; Zhou, Y.; Si, D.; Zhang, R. Biofilm and Small Colony Variants An Update on Staphylococcus aureus Strategies toward Drug Resistance. Int. J. Mol. Sci. 2022, 23, 1241. [Google Scholar] [CrossRef]
- Sanchini, A. Recent Developments in Phenotypic and Molecular Diagnostic Methods for Antimicrobial Resistance Detection in Staphylococcus aureus: A Narrative Review. Diagnostics 2022, 12, 208. [Google Scholar] [CrossRef]
- Naber, C.K. Staphylococcus aureus bacteremia: Epidemiology, pathophysiology, and management strategies. Clin. Infect. Infect. Dis. 2009, 48, S231–S237. [Google Scholar] [CrossRef]
- Holden, M.T.; Feil, E.J.; Lindsay, J.A.; Peacock, S.J.; Day, N.P.; Enright, M.C.; Foster, T.J.; Moore, C.E.; Hurst, L.; Atkin, R. Complete genomes of two clinical Staphylococcus aureus strains: Evidence for the rapid evolution of virulence and drug resistance. Proc. Natl. Acad. Sci. USA 2004, 101, 9786–9791. [Google Scholar] [CrossRef]
- Hiramatsu, K.; Katayama, Y.; Yuzawa, H.; Ito, T. Molecular genetics of methicillin-resistant Staphylococcus aureus. Int. J. Med. Microbiol. 2002, 292, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Moussa, I.; Shibl, A.M. Molecular characterization of methicillin-resistant Staphylococcus aureus recovered from outpatient clinics in Riyadh, Saudi Arabia. Saudi Med. J. 2009, 30, 611–617. [Google Scholar]
- Williamson, D.A.; Monecke, S.; Heffernan, H.; Ritchie, S.R.; Roberts, S.A.; Upton, A.; Thomas, M.G.; Fraser, J.D. High usage of topical fusidic acid and rapid clonal expansion of fusidic Acid–Resistant Staphylococcus aureus: A cautionary tale. Clin. Infect. Dis. 2014, 59, 1451–1454. [Google Scholar] [CrossRef]
- Udo, E.; O’brien, F.; Al-Sweih, N.; Noronha, B.; Matthew, B.; Grubb, W. Genetic lineages of community-associated methicillin-resistant Staphylococcus aureus in Kuwait hospitals. J. Clin. Microbiol. 2008, 46, 3514–3516. [Google Scholar] [CrossRef]
- Udo, E.E.; Al-Lawati, B.-H.; Al-Muharmi, Z.; Thukral, S. Genotyping of methicillin-resistant Staphylococcus aureus in the Sultan Qaboos University Hospital, Oman reveals the dominance of Panton–Valentine leucocidin-negative ST6-IV/t304 clone. New Microbes New Infect. 2014, 2, 100–105. [Google Scholar] [CrossRef] [PubMed]
- Udo, E.E.; Boswihi, S.S.; Mathew, B.; Noronha, B.; Verghese, T.; Al-Jemaz, A.; Al Saqer, F. Emergence of methicillin-resistant Staphylococcus aureus belonging to clonal complex 15 (CC15-MRSA) in Kuwait hospitals. Infect. Drug Resist. 2020, 13, 617. [Google Scholar] [CrossRef]
- Albarrag, A.; Shami, A.; Almutairi, A.; Alsudairi, S.; Aldakeel, S.; Al-Amodi, A. Prevalence and molecular genetics of methicillin-resistant Staphylococcus aureus colonization in nursing homes in Saudi Arabia. Can. J. Infect. Dis. Med. Microbiol. 2020, 2020, 2434350. [Google Scholar] [CrossRef]
- Donlan, R.M.; Costerton, J.W. Biofilms: Survival mechanisms of clinically relevant microorganisms. Clin. Microbiol. Rev. 2002, 15, 167–193. [Google Scholar] [CrossRef]
- Piechota, M.; Kot, B.; Frankowska-Maciejewska, A.; Grużewska, A.; Woźniak-Kosek, A. Biofilm Formation by Methicillin-Resistant and Methicillin-Sensitive Staphylococcus aureus Strains from Hospitalized Patients in Poland. Biomed. Res. Int. 2018, 27, 4657396. [Google Scholar]
- Alabdullatif, M.; Alzahrani, A. Expression of biofilm-associated genes in Staphylococcus aureus during storage of platelet concentrates. Transfus. Apher. Sci. 2022, 61, 103456. [Google Scholar] [CrossRef] [PubMed]
- Bui, L.M.; Conlon, B.P.; Kidd, S.P. Antibiotic tolerance and the alternative lifestyles of Staphylococcus aureus. Essays Biochem. 2017, 61, 71–79. [Google Scholar]
- Cucarella, C.; Solano, C.; Valle, J.; Amorena, B.; Lasa, I.N.I.; Penadés, J.R. Bap, a Staphylococcus aureus surface protein involved in biofilm formation. J. Bacteriol. 2001, 183, 2888–2896. [Google Scholar] [CrossRef]
- Yamamoto, T.; Nishiyama, A.; Takano, T.; Yabe, S.; Higuchi, W.; Razvina, O.; Shi, D. Community-acquired methicillin-resistant Staphylococcus aureus: Community transmission, pathogenesis, and drug resistance. J. Infect. Chemother. 2010, 16, 225–254. [Google Scholar] [CrossRef]
- Eyre, D.W.; Golubchik, T.; Gordon, N.C.; Bowden, R.; Piazza, P.; Batty, E.M.; Ip, C.L.; Wilson, D.J.; Didelot, X.; O’Connor, L. A pilot study of rapid benchtop sequencing of Staphylococcus aureus and Clostridium difficile for outbreak detection and surveillance. BMJ Open 2012, 2, e001124. [Google Scholar] [CrossRef]
- Price, J.; Claire Gordon, N.; Crook, D.; Llewelyn, M.; Paul, J. The usefulness of whole genome sequencing in the management of Staphylococcus aureus infections. Clin. Microbiol. Infect. 2013, 19, 784–789. [Google Scholar] [CrossRef]
- Humphreys, H.; Coleman, D.C. Contribution of whole-genome sequencing to understanding of the epidemiology and control of meticillin-resistant Staphylococcus aureus. J. Hosp. Infect. 2019, 102, 189–199. [Google Scholar] [CrossRef]
- Bazaid, A.S.; Barnawi, H.; Qanash, H.; Alsaif, G.; Aldarhami, A.; Gattan, H.; Alharbi, B.; Alrashidi, A.; Al-Soud, W.A.; Moussa, S.; et al. Bacterial Coinfection and Antibiotic Resistance Profiles among Hospitalised COVID-19 Patients. Microorganisms 2022, 10, 495. [Google Scholar] [CrossRef]
- Patel, J.B. Performance Standards for Antimicrobial Susceptibility Testing; Clinical and Laboratory Standards Institute: Wayne, MI, USA, 2017. [Google Scholar]
- Sabeel, S.; Salih, M.A.; Ali, M.; El-Zaki, S.-E.; Abuzeid, N.; Elgadi, Z.A.M.; Altayb, H.N.; Elegail, A.; Ibrahim, N.Y.; Elamin, B.K. Phenotypic and genotypic analysis of multidrug-resistant Mycobacterium tuberculosis isolates from Sudanese patients. Tuberc. Res. Treat. 2017, 2017, 8340746. [Google Scholar]
- Abdulhakeem, M.A.; Alreshidi, M.; Bardakci, F.; Hamadou, W.S.; De Feo, V.; Noumi, E.; Snoussi, M. Molecular Identification of Bacteria Isolated from Marketed Sparus aurata and Penaeus indicus Sea Products: Antibiotic Resistance Profiling and Evaluation of Biofilm Formation. Life 2023, 13, 548. [Google Scholar] [CrossRef]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef]
- Antipov, D.; Hartwick, N.; Shen, M.; Raiko, M.; Lapidus, A.; Pevzner, P.A. plasmidSPAdes: Assembling plasmids from whole genome sequencing data. Bioinformatics 2016, 32, 3380–3387. [Google Scholar] [CrossRef]
- Jolley, K.A.; Bray, J.E.; Maiden, M.C. Open-access bacterial population genomics: BIGSdb software, the PubMLST. org website and their applications. Wellcome Open Res. 2018, 3, 124. [Google Scholar] [CrossRef] [PubMed]
- Tatusova, T.; DiCuccio, M.; Badretdin, A.; Chetvernin, V.; Nawrocki, E.P.; Zaslavsky, L.; Lomsadze, A.; Pruitt, K.D.; Borodovsky, M.; Ostell, J. NCBI prokaryotic genome annotation pipeline. Nucleic Acids Res. 2016, 44, 6614–6624. [Google Scholar] [CrossRef]
- Bartels, M.D.; Petersen, A.; Worning, P.; Nielsen, J.B.; Larner-Svensson, H.; Johansen, H.K.; Andersen, L.P.; Jarløv, J.O.; Boye, K.; Larsen, A.R. Comparing whole-genome sequencing with Sanger sequencing for spa typing of methicillin-resistant Staphylococcus aureus. J. Clin. Microbiol. 2014, 52, 4305–4308. [Google Scholar] [CrossRef]
- Monecke, S.; Coombs, G.; Shore, A.C.; Coleman, D.C.; Akpaka, P.; Borg, M.; Chow, H.; Ip, M.; Jatzwauk, L.; Jonas, D. A field guide to pandemic, epidemic and sporadic clones of methicillin-resistant Staphylococcus aureus. PLoS ONE 2011, 6, e17936. [Google Scholar] [CrossRef] [PubMed]
- Strommenger, B.; Kettlitz, C.; Weniger, T.; Harmsen, D.; Friedrich, A.; Witte, W. Assignment of Staphylococcus isolates to groups by spa typing, SmaI macrorestriction analysis, and multilocus sequence typing. J. Clin. Microbiol. 2006, 44, 2533–2540. [Google Scholar] [CrossRef]
- Johansson, M.H.; Bortolaia, V.; Tansirichaiya, S.; Aarestrup, F.M.; Roberts, A.P.; Petersen, T.N. Detection of mobile genetic elements associated with antibiotic resistance in Salmonella enterica using a newly developed web tool: MobileElementFinder. J. Antimicrob. Chemother. 2021, 76, 101–109. [Google Scholar] [CrossRef] [PubMed]
- Zankari, E.; Hasman, H.; Cosentino, S.; Vestergaard, M.; Rasmussen, S.; Lund, O.; Aarestrup, F.M.; Larsen, M.V. Identification of acquired antimicrobial resistance genes. J. Antimicrob. Chemother. 2012, 67, 2640–2644. [Google Scholar] [CrossRef]
- Arndt, D.; Grant, J.R.; Marcu, A.; Sajed, T.; Pon, A.; Liang, Y.; Wishart, D.S. PHASTER: A better, faster version of the PHAST phage search tool. Nucleic Acids Res. 2016, 44, W16–W21. [Google Scholar] [CrossRef]
- Seemann, T. Prokka: Rapid prokaryotic genome annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef]
- Page, A.J.; Cummins, C.A.; Hunt, M.; Wong, V.K.; Reuter, S.; Holden, M.T.; Fookes, M.; Falush, D.; Keane, J.A.; Parkhill, J. Roary: Rapid large-scale prokaryote pan genome analysis. Bioinformatics 2015, 31, 3691–3693. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed]
- Hadfield, J.; Croucher, N.J.; Goater, R.J.; Abudahab, K.; Aanensen, D.M.; Harris, S.R. Phandango: An interactive viewer for bacterial population genomics. Bioinformatics 2018, 34, 292–293. [Google Scholar] [CrossRef]
- Brynildsrud, O.; Bohlin, J.; Scheffer, L.; Eldholm, V. Rapid scoring of genes in microbial pan-genome-wide association studies with Scoary. Genome Biol. 2016, 17, 238. [Google Scholar] [CrossRef] [PubMed]
- Åvall-Jääskeläinen, S.; Koort, J.; Simojoki, H.; Taponen, S. Genomic analysis of Staphylococcus aureus Isolates associated with peracute non-gangrenous or gangrenous mastitis and comparison with other mastitis-associated Staphylococcus aureus Isolates. Front. Microbiol. 2021, 12, 688819. [Google Scholar] [CrossRef] [PubMed]
- Tsergouli, K.; Karampatakis, T.; Kontopoulou, K.; Pappa, S.; Kampouridou, P.; Kallasidou, G.; Tsioka, K.; Zotou, S.; Farmaki, Ε.-E.; Kotzamanidis, C. Spa diversity and genetic characterization of t127 methicillin-resistant Staphylococcus aureus in a tertiary Greek hospital. Acta Microbiol. Et Immunol. Hung. 2022, 69, 185–192. [Google Scholar] [CrossRef]
- Al Yousef, S.A.; Taha, E.M. Methicillin-resistant Staphylococcus aureus in Saudi Arabia: Genotypes distribution review. Saudi J. Med. Med. Sci. 2016, 4, 2. [Google Scholar] [CrossRef]
- Al-Zahrani, I.A.; Azhar, E.I.; Jiman-Fatani, A.A.; Siddig, L.A.; Yasir, M.; Al-Ghamdi, A.K.; Harwood, C.R. Impact of mass migrations on the clonal variation of clinical Staphylococcus aureus strains isolated from the Western region of Saudi Arabia. J. Infect. Public Health 2019, 12, 317–322. [Google Scholar] [CrossRef]
- Moghaddam, P.Z.; Azimian, A.; Sepahy, A.A.; Iranbakhsh, A. Isolation and Genetic Characterization of Vancomycin-resistant and mecC+ Methicillin-resistant Staphylococcus aureus Strains in Clinical Samples of Bojnurd, Northeastern Iran. Jundishapur J. Microbiol. 2021, 14, e118949. [Google Scholar] [CrossRef]
- Narui, K.; Noguchi, N.; Wakasugi, K.; Sasatsu, M. Cloning and characterization of a novel chromosomal drug efflux gene in Staphylococcus aureus. Biol. Pharm. Bull. 2002, 25, 1533–1536. [Google Scholar] [CrossRef]
- Paharik, A.E.; Kotasinska, M.; Both, A.; Hoang, T.M.N.; Büttner, H.; Roy, P.; Fey, P.D.; Horswill, A.R.; Rohde, H. The metalloprotease S ep A governs processing of accumulation-associated protein and shapes intercellular adhesive surface properties in Staphylococcus epidermidis. Mol. Microbiol. 2017, 103, 860–874. [Google Scholar] [CrossRef]
- Senok, A.; Somily, A.M.; Nassar, R.; Garaween, G.; Sing, G.K.; Müller, E.; Reissig, A.; Gawlik, D.; Ehricht, R.; Monecke, S. Emergence of novel methicillin-resistant Staphylococcus aureus strains in a tertiary care facility in Riyadh, Saudi Arabia. Infect. Drug Resist. 2019, 12, 2739. [Google Scholar] [CrossRef]
- Mahdiyoun, S.M.; Kazemian, H.; Ahanjan, M.; Houri, H.; Goudarzi, M. Frequency of aminoglycoside-resistance genes in methicillin-resistant Staphylococcus aureus (MRSA) isolates from hospitalized patients. Jundishapur J. Microbiol. 2016, 9, e35052. [Google Scholar] [CrossRef] [PubMed]
- Okiki, P.; Eromosele, E.; Ade-Ojo, P.; Sobajo, O.; Idris, O.; Agbana, R. Occurrence of mecA and blaZ genes in methicillin-resistant Staphylococcus aureus associated with vaginitis among pregnant women in Ado-Ekiti, Nigeria. New Microbes New Infect. 2020, 38, 100772. [Google Scholar] [CrossRef] [PubMed]
- McCarthy, A.J.; Lindsay, J.A. The distribution of plasmids that carry virulence and resistance genes in Staphylococcus aureus is lineage associated. BMC Microbiol. 2012, 12, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Gan, T.; Shu, G.; Fu, H.; Yan, Q.; Zhang, W.; Tang, H.; Yin, L.; Zhao, L.; Lin, J. Antimicrobial resistance and genotyping of Staphylococcus aureus obtained from food animals in Sichuan Province, China. BMC Vet. Res. 2021, 17, 177. [Google Scholar] [CrossRef]
- Wang, H.; Zhuang, H.; Ji, S.; Sun, L.; Zhao, F.; Wu, D.; Shen, P.; Jiang, Y.; Yu, Y.; Chen, Y. Distribution of erm genes among MRSA isolates with resistance to clindamycin in a Chinese teaching hospital. Infect. Genet. Evol. 2021, 96, 105127. [Google Scholar] [CrossRef]
- Senok, A.; Somily, A.; Raji, A.; Gawlik, D.; Al-Shahrani, F.; Baqi, S.; Boswihi, S.; Skakni, L.; Udo, E.E.; Weber, S. Diversity of methicillin-resistant Staphylococcus aureus CC22-MRSA-IV from Saudi Arabia and the Gulf region. Int. J. Infect. Dis. 2016, 51, 31–35. [Google Scholar] [CrossRef]
- Shittu, A.O.; Udo, E.E.; Lin, J. Insights on virulence and antibiotic resistance: A review of the accessory genome of Staphylococcus aureus. Wounds 2007, 19, 237. [Google Scholar]
- Ito, T.; Kuwahara-Arai, K.; Katayama, Y.; Uehara, Y.; Han, X.; Kondo, Y.; Hiramatsu, K. Staphylococcal cassette chromosome mec (SCCmec) analysis of MRSA. In Methicillin-Resistant Staphylococcus aureus (MRSA) Protocols; Springer: Berlin/Heidelberg, Germany, 2014; pp. 131–148. [Google Scholar]
- Laham, N.A.; Mediavilla, J.R.; Chen, L.; Abdelateef, N.; Elamreen, F.A.; Ginocchio, C.C.; Pierard, D.; Becker, K.; Kreiswirth, B.N. MRSA clonal complex 22 strains harboring toxic shock syndrome toxin (TSST-1) are endemic in the primary hospital in Gaza, Palestine. PLoS ONE 2015, 10, e0120008. [Google Scholar] [CrossRef]
- Al-Saleh, A.; Shahid, M.; Farid, E.; Bindayna, K. Trends in methicillin-resistant Staphylococcus aureus in the Gulf Cooperation Council countries: Antibiotic resistance, virulence factors and emerging strains. East. Mediterr. Health J. 2022, 28, 434–443. [Google Scholar] [CrossRef]
- Zong, Z.; Lü, X. Characterization of a new SCC mec element in Staphylococcus cohnii. PLoS ONE 2010, 5, e14016. [Google Scholar] [CrossRef] [PubMed]
- Jalil, M.; Quddos, F.; Anwer, F.; Nasir, S.; Rahman, A.; Alharbi, M.; Alshammari, A.; Alshammari, H.K.; Ali, A. Comparative Pan-Genomic Analysis Revealed an Improved Multi-Locus Sequence Typing Scheme for Staphylococcus aureus. Genes 2022, 13, 2160. [Google Scholar] [CrossRef]
- Naushad, S.; Nobrega, D.B.; Naqvi, S.A.; Barkema, H.W.; De Buck, J. Genomic analysis of bovine Staphylococcus aureus isolates from milk to elucidate diversity and determine the distributions of antimicrobial and virulence genes and their association with mastitis. Msystems 2020, 5, e00063-20. [Google Scholar] [CrossRef] [PubMed]
- Hussain, K.; Bandyopadhyay, A.; Roberts, N.; Mughal, N.; Moore, L.S.; Fuller, L.C. Panton-Valentine Leukocidin producing Staphylococcus aureus: A clinical review. Clin. Exp. Dermatol. 2022, 47, 2150–2158. [Google Scholar] [CrossRef]
- Mohamadou, M.; Essama, S.R.; Ngonde Essome, M.C.; Akwah, L.; Nadeem, N.; Gonsu Kamga, H.; Sattar, S.; Javed, S. High prevalence of Panton-Valentine leukocidin positive, multidrug resistant, Methicillin-resistant Staphylococcus aureus strains circulating among clinical setups in Adamawa and Far North regions of Cameroon. PLoS ONE 2022, 17, e0265118. [Google Scholar] [CrossRef]
- Ullah, N.; Nasir, S.; Ishaq, Z.; Anwer, F.; Raza, T.; Rahman, M.; Alshammari, A.; Alharbi, M.; Bae, T.; Rahman, A. Comparative Genomic Analysis of a Panton–Valentine Leukocidin-Positive ST22 Community-Acquired Methicillin-Resistant Staphylococcus aureus from Pakistan. Antibiotics 2022, 11, 496. [Google Scholar] [CrossRef] [PubMed]
- Panton, P.; Came, M.; Valentine, F. Staphylococcal toxin. Lancet 1932, 219, 506–508. [Google Scholar] [CrossRef]
- Boakes, E.; Kearns, A.; Ganner, M.; Perry, C.; Hill, R.; Ellington, M. Distinct bacteriophages encoding Panton-Valentine leukocidin (PVL) among international methicillin-resistant Staphylococcus aureus clones harboring PVL. J. Clin. Microbiol. 2011, 49, 684–692. [Google Scholar] [CrossRef]
- El-Deeb, W.; Cave, R.; Fayez, M.; Alhumam, N.; Quadri, S.; Mkrtchyan, H.V. Methicillin Resistant Staphylococci Isolated from Goats and Their Farm Environments in Saudi Arabia Genotypically Linked to Known Human Clinical Isolates: A Pilot Study. Microbiol. Spectr. 2022, 10, e00387-322. [Google Scholar] [CrossRef]
- Hassan, E.A.; Abdel-Rahim, M.H.; Hassan, T.H.M.; A Azoz, N.M.; Mohamed, M.E. Some virulence genes of Staphylococcus aureus isolated from infected vascular accesses in hemodialysis patients at Assiut University Hospitals. Bull. Pharm. Sci. Assiut 2022, 45, 371–387. [Google Scholar] [CrossRef]
- Alghizzi, M.; Shami, A. The prevalence of Staphylococcus aureus and methicillin resistant Staphylococcus aureus in milk and dairy products in Riyadh, Saudi Arabia. Saudi J. Biol. Sci. 2021, 28, 7098–7104. [Google Scholar] [CrossRef] [PubMed]
- Valenciano-Bellido, S.; Caaveiro, J.M.M.; Morante, K.; Sushko, T.; Nakakido, M.; Nagatoishi, S.; Tsumoto, K. Structure and role of the linker domain of the iron surface-determinant protein IsdH in heme transportation in Staphylococcus aureus. J. Biol. Chem. 2022, 298, 101995. [Google Scholar] [CrossRef]
- Sharp, K.H.; Schneider, S.; Cockayne, A.; Paoli, M. Crystal structure of the heme-IsdC complex, the central conduit of the Isd iron/heme uptake system in Staphylococcus aureus. J. Biol. Chem. 2007, 282, 10625–10631. [Google Scholar] [CrossRef] [PubMed]
- Pishchany, G.; Sheldon, J.R.; Dickson, C.F.; Alam, M.T.; Read, T.D.; Gell, D.A.; Heinrichs, D.E.; Skaar, E.P. IsdB-dependent hemoglobin binding is required for acquisition of heme by Staphylococcus aureus. J. Infect. Dis. 2014, 209, 1764–1772. [Google Scholar] [CrossRef]
- Hemmadi, V.; Biswas, M. An overview of moonlighting proteins in Staphylococcus aureus infection. Arch. Microbiol. 2021, 203, 481–498. [Google Scholar] [CrossRef]
- Porayath, C.; Suresh, M.K.; Biswas, R.; Nair, B.G.; Mishra, N.; Pal, S. Autolysin mediated adherence of Staphylococcus aureus with Fibronectin, Gelatin and Heparin. Int. J. Biol. Macromol. 2018, 110, 179–184. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.; Ma, S.X.; St. John, A.; Torres, V.J. The Major Autolysin Atl Regulates the Virulence of Staphylococcus aureus by Controlling the Sorting of LukAB. Infect. Immun. 2022, 90, e00056-22. [Google Scholar]
- de Souza, G.M.; de Jesus, M.F.A.; de Souza Ferreira, M.V.; de Oliveira Vieira, K.C.; Nakagaki, W.R.; Lordelo, E.P.; Cataneli, V.P.; Winkelstroter, L.K. Virulence, biofilm formation ability and antimicrobial resistance of Staphylococcus aureus isolated from cell phones of university students. ABCS Health Sci. 2022, 47, e022203. [Google Scholar] [CrossRef]
- Ahmed, O.B. Prevalence of Exfoliative and Toxic Shock Syndrome Toxin Genes in Methicillin-resistant Staphylococcus aureus Strains Isolated from Clinical Specimens in Makkah, Saudi Arabia. Br. J. Med. Med. Res. 2016, 11, 1. [Google Scholar] [CrossRef]
- Hamdan-Partida, A.; González-García, S.; Martínez-Ruíz, F.J.; Zavala-Sánchez, M.Á.; Bustos-Hamdan, A.; Bustos-Martínez, J. Molecular Characterization of Staphylococcus aureus Strains Isolated from Mobile Phones. Microorganisms 2022, 10, 669. [Google Scholar] [CrossRef]
- Ahmed, O.B. Prevalence of Exfoliative and Toxic Shock Syndrome Genes in Methicillin-Resistant Staphylococcus aureus Strains Isolated from Clinical Specimens. Curr. Overv. Dis. Health Res. 2022, 4, 120–128. [Google Scholar]
- Opal, S.; Johnson-Winegar, A.; Cross, A. Staphylococcal scalded skin syndrome in two immunocompetent adults caused by exfoliatin B-producing Staphylococcus aureus. J. Clin. Microbiol. 1988, 26, 1283–1286. [Google Scholar] [CrossRef] [PubMed]
- Udo, E.E.; Boswihi, S.S.; Al-Sweih, N. High prevalence of toxic shock syndrome toxin–producing epidemic methicillin-resistant Staphylococcus aureus 15 (EMRSA-15) strains in Kuwait hospitals. New Microbes New Infect. 2016, 12, 24–30. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Gender | Location | Sample |
|---|---|---|---|
| S1 | Male | Ward | Wound |
| S8 | Female | Ward | Wound |
| S9 | Male | ICU | Throat swab |
| S14 | Male | PICU | Eye swab |
| S20 | Male | ICU | Sputum |
| S21 | Female | Ward | Wound |
| S22 | Male | Ward | Pleural Fluid |
| S23 | Male | Ward | Wound |
| Strain | Resistance Profile |
|---|---|
| S1 | Cefoxitin; Cefotaxime; Ampicillin; Penicillin G; Oxacillin; Trimethoprim; Erythromycin; Ciprofloxacin; Tetracycline |
| S8 | Gentamicin; Cefoxitin; Cefotaxime; Ampicillin; Penicillin G |
| S9 | Gentamicin; Cefoxitin; Cefotaxime; Ampicillin; Penicillin G; Oxacillin |
| S14 | Cefotaxime; Ceftaroline; Ampicillin |
| S20 | Cefoxitin; Cefotaxime; Ceftaroline; Ampicillin; Penicillin G; Teicoplanin; Vancomycin; Clindamycin; Erythromycin; Linezolid; Tetracycline |
| S21 | Cefoxitin; Cefotaxime; Ceftaroline; Ampicillin; Penicillin G; Oxacillin; Daptomycin; Trimethoprim; Vancomycin |
| S22 | Cefoxitin; Cefotaxime; Ampicillin; Penicillin G; Teicoplanin; Vancomycin; Clindamycin; Erythromycin; Ciprofloxacin; Levofloxacin; Moxifloxacin; Tetracycline |
| S23 | Cefoxitin; Cefotaxime; Ampicillin; Penicillin G; Oxacillin; Clindamycin; Erythromycin |
| Antibiotics Tested | S1 | S8 | S9 | S14 | S20 | S21 | S22 | S23 | ARI |
|---|---|---|---|---|---|---|---|---|---|
| Gentamicin | S | R | R | S | S | S | S | S | 2/8 = 0.25 |
| Cefoxitin | R | R | R | S | R | R | R | R | 7/8 = 0.875 |
| Cefotaxime | R | R | R | R | R | R | R | R | 8/8 = 1 |
| Ceftaroline | S | S | S | R | R | R | I | S | 3/8 = 0.375 |
| Ampicillin | R | R | R | R | R | R | R | R | 8/8 = 1 |
| Penicillin G | R | R | R | S | R | R | R | R | 7/8 = 0.875 |
| Oxacillin | R | S | R | S | S | R | S | R | 4/8 = 0.5 |
| Daptomycin | S | S | S | S | S | R | S | S | 1/8 = 0.125 |
| Trimethoprim | R | S | S | S | S | R | S | S | 2/8 = 0.25 |
| Teicoplanin | S | S | S | S | R | S | R | S | 2/8 = 0.25 |
| Vancomycin | S | S | S | S | R | R | R | S | 3/8 = 0.375 |
| Clindamycin | S | S | S | S | R | S | R | R | 3/8 = 0.375 |
| Erythromycin | R | S | S | R | R | S | R | R | 5/8 = 0.625 |
| Linezolid | S | S | S | S | R | S | S | S | 1/8 = 0.125 |
| Nitrofurantoin | S | S | S | S | S | S | S | S | 0/8 = 0 |
| Ciprofloxacin | R | S | S | S | S | S | R | S | 2/8 = 0.25 |
| Levofloxacin | S | S | S | S | S | S | R | S | 1/8 = 0.125 |
| Moxifloxacin | S | S | S | S | S | S | R | S | 1/8 = 0.125 |
| Rifampin | S | S | S | S | S | S | S | S | 0/8 = 0 |
| Tetracycline | R | S | S | S | R | S | R | S | 3/8 = 0.375 |
| Tigecycline | S | S | S | S | S | S | S | S | 0/8 = 0 |
| MARI | 0.428 | 0.238 | 0.285 | 0.190 | 0.523 | 0.428 | 0.571 | 0.333 |
| Isolate | ST | Spa Type | MLST CCs | Genome Length | No of Contigs | Coverage | N50 | GC | CDS | tRNA | rRNA |
|---|---|---|---|---|---|---|---|---|---|---|---|
| S1 | 1 | t127 | CC01 | 2,806,634 | 21 | 438 | 336,511 | 32.7% | 2659 | 54 | 4 |
| S8 | 97 | t189 | CC97 | 2,758,541 | 32 | 463 | 300,844 | 32.7% | 2632 | 56 | 3 |
| S9 | 97 | t2297 | CC97 | 2,853,553 | 48 | 403 | 286,668 | 32.7% | 2777 | 55 | 3 |
| S14 | 121 | t314 | CC121 | 2,778,595 | 61 | 331 | 113,582 | 32.7% | 2634 | 58 | 4 |
| S20 | 1 | t5388 | CC01 | 2,785,405 | 14 | 331 | 547,174 | 32.7% | 2645 | 55 | 3 |
| S21 | 22 | t845 | C22 | 2,742,174 | 44 | 471 | 173,813 | 32.7% | 2607 | 53 | 5 |
| S22 | 291 | t3649 | CC291 | 2,755,471 | 34 | 526 | 211,298 | 32.7% | 2632 | 53 | 3 |
| S23 | 6 | t304 | CC6 | 2,750,772 | 26 | 485 | 298,524 | 32.7% | 2601 | 57 | 4 |
| Biosample/Accession number: S1 (SAMN31278742/JAOXRK000000000); S8 (SAMN31278743/JAOXRJ000000000); S9 (SAMN31278744/JAOXRI000000000); S14 (SAMN31278745/JAOXRH000000000); S20 (SAMN31278746/JAOXRG000000000); S21 (SAMN31278747/JAOXRF000000000); S22 (SAMN31278748/JAOXRE000000000); S23 (SAMN31278749/JAOXRD000000000). | |||||||||||
| Gene | Resistance Mechanism | S1 | S8 | S9 | S14 | S20 | S21 | S22 | S23 |
|---|---|---|---|---|---|---|---|---|---|
| mecA | Antibiotic target replacement | x | x | x | |||||
| mecC | Antibiotic target replacement | x | x | ||||||
| arlR | Antibiotic efflux | x | x | x | x | x | x | x | x |
| arlS | Antibiotic efflux | x | x | x | x | ||||
| S. aureus norA | Antibiotic efflux | x | x | x | x | x | x | x | x |
| mgrA | Antibiotic efflux | x | x | x | x | x | x | x | x |
| dfrC | Antibiotic target replacement | x | |||||||
| fusC | Antibiotic target protection | x | x | x | x | ||||
| S. aureus FosB | Antibiotic inactivation | x | |||||||
| mepR | Antibiotic efflux | x | x | x | x | x | x | x | x |
| APH(3′)-IIIa | Antibiotic inactivation | x | |||||||
| AAC(6′)-APH(2″) | Antibiotic inactivation | x | x | ||||||
| norC | Antibiotic efflux | x | x | x | x | x | x | x | x |
| S. aureus LmrS | Antibiotic efflux | x | x | x | x | x | x | x | x |
| sepA | Antibiotic efflux | x | x | x | x | x | x | x | x |
| sdrM | Antibiotic efflux | x | x | x | x | x | x | x | x |
| tet(45) | Antibiotic efflux | x | |||||||
| tetK | Antibiotic efflux | x | |||||||
| PC1 beta-lactamase (blaZ) | Antibiotic inactivation | x | x | x | x | x | x | x | |
| SAT-4 | Antibiotic inactivation | x | |||||||
| ErmC | Antibiotic target alteration | x | x |
| Strain | Plasmid | Contig | Coverage | Plasmid Identity | Reference | Genes | ||
|---|---|---|---|---|---|---|---|---|
| Name | Position in Contig | Identity | ||||||
| S1 | pE5 | 1 | 1573 | 99% | M17990.1 | erm(C) | 1542–2276 | 100% |
| rep10 | 251–727 | 100% | ||||||
| S8 | pUSA300HOUMS | 1 | 697 | 99% | CP000732.1 | rep20 | 19,264–20,253 | 100% |
| blaZ | 10,806–9961 | 100% | ||||||
| ISSau6 | 10,806–9961 | 100% | ||||||
| S9 | pUSA300HOUMS | 3 | 426 | 99% | CP000732.1 | rep20 | 2896–3885 | 100% |
| blaZ | 13,996–14,841 | 100% | ||||||
| ISSau6 | 13,996–14,841 | 100% | ||||||
| S14 | pUSA02 | 2 | 1477 | 99% | CP000257.1 | tetK | 1425–46 | 100% |
| rep7a | 3379–4323 | 100% | ||||||
| pE5 | 3 | 5.73 | 99% | M17990.1 | rep10 | 512–988 | 100% | |
| erm(C) | 1803–2537 | 100% | ||||||
| S20 | pER07993.3A.1 | 1 | 648 | 99% | CP049391.1 | rep5a | 1669–2529 | 100% |
| blaZ | 8989–9834 | 100% | ||||||
| S21 | - | - | - | - | - | - | - | - |
| S22 | pLDNT_611 | 1 | 897 | 99% | CP080252.1 | rep5a | 1066–206 | 100% |
| rep16 | 2357–3100 | 100% | ||||||
| blaZ | 13,545–14,390 | 100% | ||||||
| S23 | pE5 | 2 | 2296 | 99% | M17990.1 | rep10 | 1746–2222 | 100% |
| erm(C) | 197–931 | 100% | ||||||
| Isolate | SCCmec Type | SCCmec Gene Cassettes | Identity (%) | Ref. Coverage | Contig No | Contig Position |
|---|---|---|---|---|---|---|
| S1 | N/A | ccrB1:1:COL:CP000046 | 92.25 | 1626/1625 | S1_contig_2 | 63,208–64,832 |
| ccrA1:1:COL:CP000046 | 94.37 | 1350/1350 | S1_contig_2 | 64,854–66,203 | ||
| S8 | N/A | ccrC1-allele-8:1:AB462393 | 99.94 | 1677/1677 | S8_contig_3 | 1522–3198 |
| mec-class-C2:3:AB478780 | 99.91 | 2290/2398 | S8_contig_18 | 2493–4782 | ||
| S9 | N/A | ccrC1-allele-8:1:AB462393 | 98.45 | 1677/1677 | S9_contig_20 | 1462–3138 |
| mec-class-C2:3:AB478780 | 100 | 2198/2398 | S9_contig_23 | 2400–4597 | ||
| ccrC1-allele-2:1:AB512767 | 100 | 1680/1680 | S9_contig_1 | 7153–8832 | ||
| S14 | N/A | mec-class-C2:5:AB505629 | 99.86 | 4408/4408 | S14_contig_44 | 295–4702 |
| ccrC1-allele-8:1:AB462393 | 99.94 | 1677/1677 | S14_contig_14 | 55,301–56,977 | ||
| S20 | No SCCmec | |||||
| S21 | IVa(2B) | mecA:5:CP000046 | 100 | 2007/2007 | S21_contig_18 | 1366–3372 |
| IS1272:2:AB033763 | 99.68 | 1550/1585 | S21_contig_34 | 1–1550 | ||
| subtype-IVa(2B):1:CA05:AB063172 | 100 | 1491/1491 | S21_contig_20 | 27,585–29,075 | ||
| dmecR1:1:AB033763 | 100 | 987/987 | S21_contig_18 | 280–1266 | ||
| ccrA2:7:81108:AB096217 | 100 | 1350/1350 | S21_contig_20 | 33,027–34,376 | ||
| ccrB2:9:JCSC4469:AB097677 | 99.94 | 1650/1650 | S21_contig_20 | 34,377–36,026 | ||
| S22 | No SCCmec | |||||
| S23 | IVa(2B) | subtype-a(2B):1:CA05:AB063172 | 99.93 | 1491/1491 | S23_contig_16 | 28,081–29,571 |
| mecA:12:AB505628 | 100 | 2010/2010 | S23_contig_4 | 285,112–287,121 | ||
| dmecR1:1:AB033763 | 100 | 987/987 | S23_contig_4 | 287,218–288,204 | ||
| IS1272:3:AM292304 | 100 | 1843/1843 | S23_contig_4 | 288,193–290,035 | ||
| ccrB2:9:JCSC4469:AB097677 | 99.94 | 1650/1650 | S23_contig_4 | 291,877–293,526 | ||
| ccrA2:7:81108:AB096217 | 100 | 1350/1350 | S23_contig_4 | 293,527–294,876 | ||
| Core Genes | (99% ≤ strains ≤ 100%) | 2027 |
| Soft-core genes | (95% ≤ strains < 99%) | 0 |
| Shell genes | (15% ≤ strains < 95%) | 992 |
| Cloud genes | (0% ≤ strains < 15%) | 818 |
| Total genes | (0% ≤ strains ≤ 100%) | 3837 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Snoussi, M.; Noumi, E.; Bouali, N.; Bazaid, A.S.; Alreshidi, M.M.; Altayb, H.N.; Chaieb, K. Antibiotic Susceptibility Profiling of Human Pathogenic Staphylococcus aureus Strains Using Whole Genome Sequencing and Genome-Scale Annotation Approaches. Microorganisms 2023, 11, 1124. https://doi.org/10.3390/microorganisms11051124
Snoussi M, Noumi E, Bouali N, Bazaid AS, Alreshidi MM, Altayb HN, Chaieb K. Antibiotic Susceptibility Profiling of Human Pathogenic Staphylococcus aureus Strains Using Whole Genome Sequencing and Genome-Scale Annotation Approaches. Microorganisms. 2023; 11(5):1124. https://doi.org/10.3390/microorganisms11051124
Chicago/Turabian StyleSnoussi, Mejdi, Emira Noumi, Nouha Bouali, Abdulrahman S. Bazaid, Mousa M. Alreshidi, Hisham N. Altayb, and Kamel Chaieb. 2023. "Antibiotic Susceptibility Profiling of Human Pathogenic Staphylococcus aureus Strains Using Whole Genome Sequencing and Genome-Scale Annotation Approaches" Microorganisms 11, no. 5: 1124. https://doi.org/10.3390/microorganisms11051124