
A Review of Modern Methods for the Detection of Foodborne Pathogens
Department of Food Science and Human Nutrition, College of Agriculture and Veterinary Medicine, Qassim University, Buraydah 51452, Saudi Arabia
Microorganisms 2023, 11(5), 1111; https://doi.org/10.3390/microorganisms11051111
Submission received: 1 February 2023
/
Revised: 9 April 2023
/
Accepted: 10 April 2023
/
Published: 24 April 2023
(This article belongs to the Special Issue Latest Review Papers in Food Microbiology 2023)
Abstract
:Despite the recent advances in food preservation techniques and food safety, significant disease outbreaks linked to foodborne pathogens such as bacteria, fungi, and viruses still occur worldwide indicating that these pathogens still constitute significant risks to public health. Although extensive reviews of methods for foodborne pathogens detection exist, most are skewed towards bacteria despite the increasing relevance of other pathogens such as viruses. Therefore, this review of foodborne pathogen detection methods is holistic, focusing on pathogenic bacteria, fungi, and viruses. This review has shown that culture-based methods allied with new approaches are beneficial for the detection of foodborne pathogens. The current application of immunoassay methods, especially for bacterial and fungal toxins detection in foods, are reviewed. The use and benefits of nucleic acid-based PCR methods and next-generation sequencing-based methods for bacterial, fungal, and viral pathogens’ detection and their toxins in foods are also reviewed. This review has, therefore, shown that different modern methods exist for the detection of current and emerging foodborne bacterial, fungal, and viral pathogens. It provides further evidence that the full utilization of these tools can lead to early detection and control of foodborne diseases, enhancing public health and reducing the frequency of disease outbreaks.
Keywords:
foodborne; pathogens; bacteria; fungi; viruses; culture-based; PCR; immunoassays; NGS; illnesses1. Introduction
Microorganisms are important in food production, safety and spoilage. Microbial interactions with food can lead to food unfit for human consumption [1,2]. This is because some of these foodborne microorganisms not only cause food spoilage but are also pathogenic in nature, presenting risks to food safety and foodborne illnesses to food handlers, consumers, and society in general [3].
Food-borne illnesses and diseases are major threats to human health and lives with over 200 foodborne diseases identified [4]. Common foodborne pathogens include bacteria such as Bacillus cereus, Clostridium botulinum, Campylobacter sp., C. perfringens, some Escherichia coli serogroups, Listeria monocytogenes, Salmonella spp., Shigella spp., Staphylococcus aureus Vibrio spp. etc. [5,6,7]. Consequently, detection methods were developed for these pathogens to ensure compliance with fixed legal and regulatory food safety thresholds based on either pathogen presence or absence and counts. Foodborne fungal species such as Penicillium, Claviceps, Aspergillus and Fusarium species which can produce mycotoxins are also a threat to public health [6]. Key foodborne pathogenic viruses include Norovirus, Hepatitis A and E viruses, Rotavirus, Enterovirus, Adenovirus, Sapovirus, Achivirus, and Astrovirus [7,8].
In recent times, there have been significant advances in food preservation methods designed to extend the shelf-life of food and eliminate spoilage and or pathogenic microbial groups. Some of the commonly used methods include storage at low temperatures (chilling and freezing), application of chemical preservatives (e.g., essential oils and bacteriocins [9,10,11,12], and vacuum and modified atmosphere packaging [13,14]. While some of these methods eliminate food pathogens, research has shown that when some of these pathogens are exposed to environmental stress or food preservation-related stress, they may survive the stress and exist as metabolically injured cells with impaired metabolic activities. In this case, the pathogens may or may not recover from this damage (irreversible or reversible). They may also survive as dormant cells, for example as viable but not culturable cells (VNBC) or as persister cells [15]. VNBC cells are cells that are no longer culturable using known growth media but possess low and detectable metabolic activities while persister cells are surviving cells tolerant to but not resistant to antibiotics, with low metabolic activities, undetectable with viability assays. However, persister cells may become culturable when exposed to a specific stimulus [15].
The regular occurrence of foodborne diseases shows that despite the significant advances in food manufacturing and safety, foodborne pathogens still constitute significant public health risks [15]. Consequently, a variety of methods, culture, and culture-independent, have been developed to detect these foodborne pathogens in food and protect public health. There have been numerous reviews of foodborne pathogens’ detection using culture-dependent and independent methods such as the use of culture media, Polymerase Chain Reactions (PCR) (multiplex PCR, Real-Time PCR, Reverse Transcriptase PCR (RT-PCR)), DNA microarray, nucleic acid sequence-based amplification (NASBA), and Next Generation Sequencing (NGS) immunological and nanotechnology-based methods [15,16,17].
However, most of these reviews are skewed towards bacteria (with a passing mention of other foodborne pathogenic groups). This is because bacteria are the most encountered foodborne pathogens of public health significance. However, other pathogenic groups such as fungi and viruses are also important as they also are known to cause foodborne diseases outbreak. Therefore, this review not only focuses on bacteria but also equally on fungi and viruses with emphasis on the use of some specific molecular methods such as PCR (End-point PCR, Multiplex PCR and RT PCR etc.) and NGS to advance our current knowledge of the detection and identification methods of these different foodborne pathogens. The latest data on the occurrence of foodborne diseases are reported with some examples of recent outbreaks of foodborne diseases being presented in the next section to demonstrate that foodborne pathogens continue to pose a significant risk to public health.
2. Outbreaks of Foodborne Illnesses
2.1. Bacteria
Foodborne illnesses caused by bacteria are a worldwide occurrence. For example, between 128,000 and 325,000 hospitalizations and 3000 to 5000 deaths, and at least 76 million illnesses per year are estimated to be associated with foodborne illnesses in the USA, costing the economy up to $83 billion per year [4,18,19]. In the EU, specifically in 2020, there were 20,017 human cases and 3086 foodborne outbreaks of foodborne diseases, with Salmonella (in eggs and egg products) being the most frequently implicated bacterial causative agent for foodborne outbreaks [20]. Apart from Salmonella, and Campylobacter, Yersinia, Shiga toxin-producing Escherichia coli (STEC) and Listeria monocytogenes were the other most common causative agents of foodborne illnesses in 2020 in the EU [20]. L. monocytogenes-related illnesses were one of the illnesses that resulted in the highest human fatalities in the EU during this time frame [20]. Worldwide, it has been estimated that there are about 3 million cases of diarrhoea related to foodborne microbial pathogens annually [19]. According to the Australian Department of Health, it is estimated that 4.1 million foodborne diseases or cases occur in Australia [21]. A review of foodborne pathogens and associated illnesses have been drawn up by authors [5].
2.2. Fungi
In contrast to bacteria and viruses, outbreaks of illnesses associated with fungal foodborne pathogens are rare given that only about 300 of the estimated 1.5 million fungal species are known to cause illnesses in humans [22]. Nevertheless, there are some notable examples of the outbreak of illnesses, typically due to fungal secondary metabolites such as toxins and sometimes occurring only in vulnerable populations such as people with suppressed immunity, with transplants or undergoing immunosuppressive treatments. However, on rare occasions, some immunocompetent people can also succumb to foodborne fungal illnesses.
For example, in 2013, there was an outbreak of gastroenteritis (vomiting, nausea and diarrhoea) in over 200 people who consumed yoghurts contaminated with Mucor circinelloides in the USA [23]. An outbreak of food poisoning by Rhizopus microsporus was reported in 7 hospital patients in Hong Kong from contaminated pre-packaged ready-to-eat meals or cornstarch used to produce allopurinol tablets [24]. A review of the literature [25] has shown some key filamentous fungi such as Aspergillus, Fusarium, and Mucor when consumed with contaminated food or inhaled can cause localized infections in the sinuses, lungs, and other areas in immunocompetent people. Invasive pulmonary disease caused by some of these fungal pathogens typically occurs in immunocompromised patients (with inhalation as a primary source). However, gastrointestinal routes could be important as well, hence the need for such patients to avoid foods likely to be contaminated with fungi [25]. An extensive survey of foodborne fungal agents was drawn up by authors [26].
2.3. Viruses
Viral outbreaks of food poisoning are less frequently reported compared to bacterial outbreaks. Concerning viruses, Noroviruses and Hepatitis A viruses are one of the most common foodborne viral pathogens known to cause illnesses in humans. Noroviruses, in particular, have been linked to up to 21 million cases of acute gastroenteritis annually in the United States of America [27]. In 2020 in the EU, Norovirus (in crustaceans, shellfish, molluscs, and their products) was a major cause of reported food outbreaks [20]. Enteric viruses are thought to account for up to 13% and 45% of outbreaks of foodborne illnesses in the EU and the US respectively [28] with sporadic reports of these outbreaks in the public domain. A list of foodborne illnesses associated with viruses has been reported by [8].
A recent review of reported virus-linked foodborne illness outbreaks has identified that the gastroenteritis outbreak at the 2018 Winter Olympics in South Korea (194 cases), the consumption of frozen raspberry-linked outbreak in 2017 in Canada (700 cases) and the outbreak in 2017 Royal Caribbean cruise line were all linked to Noroviruses [27]. Other reviews of the literature have shown that in 2014, viral agents accounted for 20% of all reported outbreaks in the EU [20]. Specific examples include the 162 Hepatitis A virus cases in the US in 2011 (consumption of contaminated pomegranate seeds) and over 1100 cases in China in 2012 (Norovirus in frozen strawberries). In 2020, West Nile virus-based illnesses were one the major foodborne pathogens that resulted in the highest number of human fatalities in the EU [20].
3. Methods for Detecting Foodborne Pathogens
The detection of pathogens in food matrices with high reproducibility and sensitivity is of utmost importance to guarantee food safety. However, it is still a difficult challenge due to factors such as interference by other and non-target microbiota, low numbers of target groups, and difficulties in microbial extraction from food matrices. There are different methods used for detecting and identifying foodborne pathogens. These range from the use of culture-based methods, immunological assays, nucleic acid-based methods (Polymerase Chain Reactions (PCR) and Next Generation Sequencing (NGS) methods. This review provides a brief explanation of the principles underpinning each class of method (excluding biosensors), where relevant, followed by a review of the application of some of the frequently used approaches for pathogen detection in the current literature. This paper reviews the use of culture-based methods, immunological assays, PCR and Next Generation Sequencing-based approaches.
However, the success of any of these approaches is dependent on the use of the appropriate aseptic sampling and sample storage protocols. The sampling method is dependent on the type of food being sampled, target microbial groups, and the microbial detection methods. For example, sample collection and analysis for any method involving the isolation and use of microbial cultures should follow standard protocols certified and developed by official organizations such as FDA, FSIS/USDA, ISO, and AOAC [29]. For other methods, standardized protocols exist for sample collection, analysis, and data interpretation based on the kit, reagent, and equipment manufacturer’s prescription.
3.1. Culture-Based Methods
Culture-based methods remain the reference methods for detecting foodborne pathogens despite the existence of other more modern methods [30,31,32,33]. These methods are premised on the ability of bacteria and fungi to grow on culture media, forming visible colonies which can then be subject to other downstream assays as required. They remain the first choice and may be required by law for the detection of microorganisms in food testing laboratories [15]. They can be used to generate culture-based qualitative and quantitative data on the presence of food pathogens. Culture-based methods are more successful when the growth requirements of target microorganisms are known with culture media being used to enrich, selectively isolate, or discriminate between target microorganisms and other groups [6]. In addition, cultures can be subject to tests such as colony characteristics, Gram staining reaction, biochemical characterization, and MALDI TOF MS and PCR-sequencing for identification purposes [6].
Despite its ease of use, relatively cheap cost, and the fact that it allows the isolation of the microorganisms for use in downstream applications, culture-based methods have significant limitations (Table 1). These methods have low sensitivity as not all microorganisms are culturable and some culturable bacteria (such as S. typhi and E. coli) can exist as VBNC, as a result of stress and other factors [15,34]. These can lead to underestimation or non-detection of foodborne pathogens creating a potential risk to food safety. Culturing microorganisms is also a slow and laborious process requiring a series of steps and may require the use of adjunct methods (e.g. biochemical, serological, nucleic acid-based methods for conclusive identification) and can take up to a week for bacteria [16,35] or longer for fungi. They also may not be suitable for the rapid detection of microorganisms for on-the-spot or real-time foodborne pathogen detection [36,37].
3.1.1. Culture-Based Detection of Foodborne Bacteria
Some of the key bacterial groups commonly associated with foodborne illnesses are E. coli, L. monocytogenes, Yersinia, Salmonella, Campylobacter, Clostridium, Enterobacteria, and Bacillus species. These bacterial groups cause foodborne illnesses/diseases by direct bacterial growth and or the secretion of toxins. These bacterial groups can be cultured using the appropriate culture media except when they are in the VBNC phase. They are usually ingested through the consumption of contaminated foods such as dairy, poultry, and beef products (including Ready to Eat (RTE) foods) [7]. Therefore, the use of cultural media alone is unreliable. Moreover, they are laborious and have limited sensitivity as they can only be used to detect a subset (culturable bacteria) with a significant population of bacteria not being culturable. Indeed, nucleic acid-based and other molecular methods with greater sensitivity and ability to detect both culturable and non-culturable bacteria should be more widely used. Instead, culture-based approaches coupled with other methods such as PCR, Immunoassays, NGS, Biosensors, and MALDI Tof MS are increasingly being used for the detection and identification of foodborne pathogens. This reduces the time needed for bacterial detection/identification from a week using traditional approaches to less than 3 days. For example, MALDI TOF MS is an accurate, sensitive, and relatively faster identification method for fastidious bacteria than PCR and NGS-based approaches [41,42]. It is also a comparatively cheaper method to use compared to NGS [43]. It involves the generation of the peptide mass fingerprint (PMF) of isolated bacterial cultures (and comparing that with the PMF in a selected database for identity typing [43].
In recent times MALDI TOF MS has been successfully used to identify foodborne pathogenic bacteria, especially those belonging to the key foodborne bacterial pathogenic groups. With respect to Salmonella, Campylobacter, and Enterobacteria, S. typhimurium from minced beef [44], Campylobacter jejuni, E. coli, and Enterobacter cloacae from Matazeez, a Middle Eastern lamb and vegetable-based stew [45] have been identified using MALDI TOF MS. MALDI TOF MS has also been used to identify E. coli, S. typhimurium , Klebsiella, Enterobacter and other food pathogens from fish, meat and dairy products from stores and supermarkets in Egypt [46]. Similarly, MALDI TOF was used to identify foodborne pathogens such as E. coli, S. typhimurium , S. aureus, C. jejuni, Klebsiella, and L. monocytogenes in fresh vegetables in Beijing, China [47], and salads, burgers, and tortillas in Saudi Arabia [45] and raw milk from public markets in Turkey [48]. With respect to Clostridium, C. difficile and C. perfringens from baby food and baby food supplements [49] have been identified with MALDI TOF MS, These reports, therefore, show that despite its limitations culture-based methods coupled with MALDI TOF MS-based approaches are currently being used for the isolation and subsequent identification by MALDI TOF MS of foodborne pathogenic bacteria.
3.1.2. Culture-Based Detection of Foodborne Fungi
Fungi and their metabolites (mycotoxin) can be ingested with contaminated food. In culture-dependent detection, fungi may take up to 7 days of incubation to be visible on culture plates. Moreover, for fungal pathogens, culture-based approaches are now widely used in conjunction with methods such as PCR, Immunoassays, and MALDI TOF MS, NGS for fungal (including pathogenic fungi) detection and identification [50,51,52].
Using MALDI TOF MS as an example, the method has been used to identify detected key fungal groups such as Rhizopus, Aspergillus, Fusarium, and Mucor [45]. Potential food pathogens such as A. niger, A. flavus, and other fungal groups such as Alternaria alternata, P. digitatum, and Candida albicans have been detected and identified in foods such as salads, burgers, tortilla, cheese, kabsa, and jareesh from restaurants in Al-Qassim regions in Saudi Arabia using MALDI TOF MS [45]. Other fungal groups such as Mucor [50], foodborne yeasts [53], and fungi in fermented foods [54] have also been detected and identified using MALDI TOF MS. However, this approach is comparatively less used for fungi than for bacteria primarily because of the limited availability of curated fungal spectra database and adaptation of the method for fungi [55].
3.1.3. Culture-Based Detection of Foodborne Viruses
Noroviruses and Hepatitis A viruses are the most common cause of foodborne illnesses/diseases [20]. Noroviruses alongside other viruses such as Rotaviruses and Astroviruses typically cause gastroenteritis while foodborne Hepatitis viruses cause either hepatitis A or E with the enteroviruses causing hand and foot diseases, meningitis/encephalitis, and heart disorders. They can contaminate food such as seafood (shellfish, oysters, and molluscs), fish, milk, fruits, and vegetables with transmission through the faecal-oral route or inhaled as droplets or through contact with contaminated items [8,56]. The only exception is the Hepatitis E virus which is associated with raw and undercooked meat and liver [8]. Detailed information on the different foodborne viruses, their route of entry, and the type of food they have been detected in have been reviewed [8].
Detection of viruses in food can be carried out by propagating the extracts from test food samples in cell cultures [27] and the observation of the formation of viral-induced cytopathic effects. Viral quantification can then be achieved through tissue culture, infectious dose 50 (TCID 50), plaque assay, and the most probable number method [57]. These methods have been applied in multiple studies to detect viruses of significant public health risks in food items. However, the use of these culture-based methods for the detection of viruses is limited by a variety of factors. These factors include the typically low concentration of viruses in food, thereby requiring the use of very sensitive methods for enrichment before detection [58]. It also takes days to culture viruses, making this approach unsuitable for rapid testing. In addition, some viruses do not show cytopathic effects and efficient cell culture systems do not exist for some of the foodborne viruses such as Noroviruses [27,57,59]. This is why molecular methods such as RT-PCR which are comparably less laborious, highly sensitive, and accurate are more widely used for viral detection. However, there are still some reports of culture-based approaches still being used for the detection of some viruses.
For example, Avian Influenza viruses (H7N9 and H5N8), a substantial risk to public health have been detected in wild birds, poultry, and ducks [60,61,62]. The method of detection typically involves homogenizing samples and growing aliquots of the homogenate in embryonated chicken eggs (cell culture) for 2–5 days followed by hemagglutination, with positive hemagglutination demonstrating the presence of the virus [63]. Sometimes, culture methods are integrated with other tools such as plaque assay methods and integrated cell culture RT-qPCR (virus propagated in cell culture and its amplification assessed with RT-qPCR) to aid viral detection. A good example is the Human adenovirus (HAdv) which has been successfully detected in fresh produce such as lettuce onions and strawberries using plaque assay and integrated cell culture RT-qPCR approaches [64].
3.2. Immunological Assays
These assays include serotyping, immunofluorescence, use of lateral flow devices (LFD) and enzyme-linked immune sorbent assay (ELISA) approaches, with ELISA, being a very accurate immunological method for detecting foodborne pathogens and their toxins. These approaches work on the principle that there is an affinity between microbial antigens and antibodies and that this affinity can be exploited for the rapid and accurate detection of foodborne pathogens (Table 1) [37]. The key advantages of these assays are that they are easier to carry out, faster than culture-based methods, can detect toxins, and can be highly specific. However, the contamination of the reaction matrix can lead to the generation of false positives (Table 1) [65]. ELISA and lateral flow devices (LFD) are some of the most widely used immunoassays for detecting foodborne microorganisms and their toxins in recent times with their principle and mode of action well reviewed [16,65,66] ELISA can sometimes be used in conjunction with other methods (including PCR), to increase their specificity and efficiency. For example, a PCR-ELISA approach has led to a 100-fold increase in the detection of Fusarium verticillioides in contaminated corn samples compared to detection using conventional PCR [67].
3.2.1. Detection of Foodborne Bacteria and Bacterial Toxins Using Immunoassays
ELISA has been used to detect key bacterial groups such as Salmonella, Campylobacter, E. coli, and Listeria. In particular, ELISA had been used to detect Salmonella sp. in meat and dairy products, pasta and eggs [68,69], Campylobacter in food samples [70] E. coli O157: H7 in cabbage [71] and beef [72], Vibrio parahaemolyticus in seafood [73] and Listeria in milk [74]. The method has also been used to detect staphylococcal enterotoxins [75], botulinum toxins in spiked food (meat and milk) [76], Shiga toxins [77], and B. cereus enterotoxins [78] in different food matrixes.
3.2.2. Detection of Foodborne Fungi and Fungal Toxins Using Immunoassays
ELISA kits (PCR-ELISA) have been developed for the detection of pathogenic F. verticillioides [67] and other species. In addition, it is commonly used to detect mycotoxins. ELISA has been used to detect aflatoxin B1 from A. flavus in stored maize [79], low levels of aflatoxin in dried stockfish [80], peanuts [81], and soy milk [82]. Similarly, LFDs have been used to detect mycotoxins such as aflatoxin, deoxynivalenol, and fumonisins in maize and barley [66].
3.2.3. Detection of Foodborne Viruses Using Immunoassays
Immunoassays are more commonly used to detect pathogenic viruses in clinical and environmental samples for point-of-care and disease diagnostic and public health surveillance reasons [38,83] than those from food sources. Historically, ELISA immunoassay kits are available for the detection of adenoviruses, group A Rotaviruses, and Astroviruses [84] and have been used for detecting viruses in food items [85]. However, they are no longer a method of choice because of the advent of nucleic acid-based methods which are more rapid and sensitive approaches.
3.3. Nucleic-Acid Based Methods (Polymerase Chain Reaction (PCR) and Its Variants (PCR, RT-PCR etc.)
PCR and its variants are nucleic acid-based methods that are used to detect specific DNA or RNA sequences of pathogenic microorganisms. Specific primers are designed for target pathogens allowing them to be exponentially amplified in food samples. PCR-based methods are faster, more sensitive, highly reproducible, and versatile approaches compared to most culture-based and immunoassay methods [65] making it a method of choice for the detection of foodborne pathogens. They are applied to nucleic acids extracted from food samples or microbial isolates isolated from food samples. The principles, advantages, and limitations of PCR-based approaches and their variants are well-reviewed (Table 1) [16,65,86,87].
There are many variants of this nucleic acid-based approach (Figure 1). These include the conventional PCRs (nested, touchdown, hot-start PCR, etc.), which are commonly applied for the detection of foodborne pathogens based on the use of primers targeting the DNA of these pathogens. Nested PCR is a modification of standard PCR and involves the use of 2 different primer sets in two PCR reaction runs. The second primer set is used to amplify a secondary target with the amplicon generated with the first primer set [86]. Nested PCR is, therefore, designed to pick up low levels of target pathogens [88]. Touchdown PCR prevents the amplification of non-specific sequences. This is achieved by starting the early PCR steps at high annealing temperatures followed by incremental decreases in subsequent cycles thereby allowing the primer in use to bind to the target sequence at the highest temperature that prevents the amplification of non-specific sequences [86]. Another variant, multiplex PCR can be used to detect multiple pathogens at the same time as it involves the use of different primers in the same reaction [86]. Reverse Transcriptase PCR (RT-PCR) allows for the detection of RNA (metabolically active microorganisms) by creating a complementary DNA (cDNA) from the RNA transcript after which the cDNA is amplified. Real-time or quantitative PCR (qPCR), is used to quantify pathogen load on food items based on the use of fluorescent dyes or probes [88] using a specially designed thermal cycler. It differs from conventional PCR in that amplification of target DNA is monitored in real-time instead of at the endpoint as obtained in conventional PCR. RT-PCR and qPCR can be combined (RT-qPCR) for qualitative and quantitative purposes. RNA is reverse transcribed into cDNA and the cDNA is subject to Real-Time PCR for quantitative detection of RNA [86]. Other forms of PCR such as droplet digital PCR (ddPCR) allow for quantification without the use of a standard curve [89,90]. In DDPCR, target samples are fractionated into thousands of droplets (each droplet being one-nanolitre reverse micelles (water in oil)) which are subject to a fluorescent probe-based PCR assay [89,90]. Loop-mediated isothermal amplification (LAMP) is another DNA amplification method that yields more DNA copies (about a billion copies) than normal PCR ( about a million copies) within an hour [91]. It involves the use of up to three primer pairs to target up to eight specific sites on DNA strands of interest in amplification reactions conducted at stable temperatures [91].
3.3.1. Detection of Foodborne Bacteria Using PCR-Based Methods
Conventional PCR and its variants (which are DNA based) involve the use of primers (specific or degenerate) that can be applied to detect a target or group of microorganisms in food items. However, these approaches (endpoint PCR, nested PCR, multiplex PCR, etc.) cannot indicate whether the target organism(s) is viable or not. These approaches have been used to successfully detect key bacterial pathogens such as Salmonella spp, Campylobacter sp., E. coli O157:H7, S. aureus, B. cereus, L. monocytogenes, and V. parahaemolyticus in ready-to-eat Korean foods [93], Minas Frescal cheese [94], milk [95], beef [96], spiked chicken egg, pork, salad [97], and fish [98].
Real-Time or quantitative PCR is more widely used than (PCR or multiplex PCR) to detect and quantify the abundance of key foodborne bacterial pathogens in extracts from food samples or purified isolates from food samples. Salmonella, E. coli, Campylobacter, and L. monocytogenes in cheese, chicken, beef burgers, turkey, pork, egg, chicken, and fish have been detected and quantified using Real-Time PCR [34,99,100,101]. Using the same method, C. perfringens, E. coli, and S. aureus were detected and quantified in fresh and ready-to-eat vegetables [102]. Other pathogens such as V. vulnificus, V. parahaemolyticus, and V. cholerae [103,104,105] have been detected in seafood, shrimp, and mussels using Real-Time PCR.
In contrast, Reverse Transcriptase PCR (RT-PCR)-based methods are not as widely used for detecting foodborne bacterial pathogens as Real Time or quantitative PCR. This is because, mRNA despite being a better indicator of the viability of bacteria, is rapidly degraded in the food matrix generating false negative results [106]. The process is also labour-intensive [106] and both of these factors account for why the approach is less used for detecting foodborne bacteria than other methods [15]. Nevertheless, this approach has been used to detect bacterial pathogens in different food samples such as S. typhimurium in artificially spiked jalapeno and serrano peppers [107] and S. enterica in spiked egg broth and milk [108].
Viable PCR (vPCR), another variant of PCR that involves the use of intercalating dyes such as ethidium monoazide (EMA) and propidium monoazide (PMA) for sample pre-treatment followed by PCR amplification allowing for the detection of living foodborne pathogens [109]. vPCR could be more readily applied for the detection of pathogens than RT PCR. These intercalating dyes can penetrate the membrane of damaged or dead bacterial cells, irreversibly binding to their DNA molecules thereby preventing their amplification by PCR primers. Therefore, any amplicon observed from the subsequent PCR will be from live bacterial cells [109]. It has been used to detect live Salmonella sp. in spiked pork meat and RTE salad [110] and Helicobacter pylori in retail pork meat [111] demonstrating the usefulness of this approach in assessing food safety [110]. Combining vPCR with Real-Time PCR has allowed for the accurate detection and quantification of viable Campylobacter sp. in frozen and chilled poultry meat [112] and Listeria sp. in spiked meat, salad, cheese, and milk [113]. Other forms of nucleic acid-based amplification methods such as LAMP (Loop-Mediated Isothermal Amplification) have been used to detect V. vulnificus (Yang et al., 2021), V. parahaemolyticus [103] in foods such as seafood.
3.3.2. Detection of Foodborne Fungi Using PCR-Based Methods
A review of the literature showed limited reports (compared to bacteria) of the detection of pathogenic fungi on food materials using PCR-based methods. However, there are multiple reports of the detection of fungal pathogens from clinical and environmental samples using nucleic acid and PCR-based methods [114,115,116,117]. Different PCR or nucleic acid-based methods such as endpoint PCR, real-time PCR, nested PCR, quantitative (RT)-PCR, loop-mediated isothermal amplification (LAMP) and multiplex PCR have been applied for the detection of pathogenic or mycotoxigenic fungi [118]. These methods have been applied directly on extracts from food samples, swabs of food surfaces, and isolates from food samples to detect and characterize foodborne fungal pathogens.
Multiplex PCR has been developed and successfully used to detect aflatoxigenic Aspergillus isolates from “meju”, a traditional fermented soybean food starter from Korea [119]. In this instance, specially designed primers were used to successfully discriminate between aflatoxin and non-aflatoxin-producing fungi, with this being validated by TLC and HPLC results of filtrates from test cultures [119]. Multiplex PCR has also been used to simultaneously detect Aspergillus, Penicillium, and Fusarium in contaminated maize grain powder [120].
Patulin (toxin) producing Penicillium expansum has also been successfully detected on artificially contaminated apples using RT-PCR [121]. Loop-mediated isothermal amplification (LAMP) assays have been developed for the rapid detection of ochratoxin-producing Penicillium nordicum in dry-cured meat products [122]. PCR combined with other methods such as denaturing gradient gel electrophoresis (DGGE) and sequencing can now be used to detect ochratoxin-producing Aspergillus niger in wine [123]. DGGE is a method that is used to separate DNA fragments (PCR amplicons) based on their melting characteristics in polyacrylamide gels. This generates a gel-based fingerprint of the key microbial groups amplified and further downstream processes can be used to characterize the identity of these groups if required [123].
3.3.3. Detection of Foodborne Viruses Using PCR-Based Methods
Molecular methods such as PCR, Multiplex PCR, Real Time-PCR (RT PCR), digital RT-PCR, and Quantitative RT PCR can also be used to detect viral pathogens in food. RT-PCR, especially, is a commonly used method probably because a significant number of viruses are RNA viruses and one of the best approaches for the detection of foodborne viral pathogens [57]. This is because it allows for the quantification of viral particles when combined with quantitative real-time PCR, which is very sensitive and specific with high throughput [27]. These can be applied directly to food samples or coupled with cell culture assay (isolation of pathogens using cell culture followed by detection using PCR) [57]. Either way, viruses must be extracted and concentrated from food materials first, before the application of PCR. The limitations of the technique are that the extraction processes may be inefficient resulting in low recovery or outright loss of viral particles [27] and the amplification process may also be affected by sample inhibitors. RT PCR or real-time quantitative PCR (RT qPCR) is also unable to distinguish between infectious and non-infectious particles [59]. One way to overcome the limitations of the detection concerning identifying infectious particles is the use of intercalating dyes such as propidium monoazide (PMA) or ethidium monoazide (EMA) for sample pre-treatment before RT PCR or RT qPCR to inhibit the amplification from non-infectious particles. This has been successfully used to distinguish infectious Hepatitis A viruses and Rotavirus from non-infectious particles in laboratory-based analysis [124].
There are many examples of the use of RT-PCR for viral detection in food. For example, the prevalence of Hepatitis A and Norovirus was investigated in harvested mussels in Italy. RT-PCR was used to show the high prevalence of norovirus conclusively demonstrating the public health risk associated with mussel consumption [125]. RT-PCR has similarly been used to detect zoonotic Hepatitis E viruses in spiked pork liver sausages [126] and in raw and liver sausages sold in retail shops in Germany [127]. In Brazil, this approach was used to assess the prevalence of Adenovirus, Hepatitis E virus and Rotavirus in beef, pork, and chicken meat cuts from a market in Brazil with results showing that Rotavirus was the most significant viral pathogen in chicken cuts in these samples [128].
RT-PCR has also been used along with new methods such as microfluidics to detect viruses in different food items or used to enhance the viral detection efficiency of other molecular methods. Raw fruits such as soft berries can harbor pathogens such as Norovirus and Hepatitis A virus which are causative agents of gastroenteritis. However, their concentrations and infectious doses tend to be low in contaminated food samples. Therefore, microfluidics was successfully applied to enhance the detection of these two viruses in soft berries using digital RT-PCR and in this instance, it lowered PCR inhibition while boosting viral detection efficiencies [129]. Similarly, microfluidic digital PCR was successfully used to detect Norovirus and Hepatitis A viruses in spiked/contaminated lettuce, with the observed viral recoveries significantly higher than those obtained with other molecular methods such as RT-PCR [130].
3.4. Next-Generation Sequencing (NGS) Methods
Next-generation sequencing (NGS) approaches coupled with bioinformatics are powerful approaches that have greatly benefitted food safety. NGS is primarily used in many ways, firstly to determine the whole genome sequence of an isolate (whole genome sequencing or WGS) and secondly, in metagenomics to determine the sequences of many of the microorganisms present in a sample. In the latter application, particular microbial groups (bacteria, fungi, or viruses) can be targeted using 16S rRNA, ITS, or any other biological markers [131]. The different workflows of NGS applied to food microbiology are shown in Figure 2. There are excellent reviews of Next Generation Sequencing methods principles, types, advantages, and disadvantages (Table 1) [39,40,132].
3.4.1. Detection and Identification of Foodborne Bacteria Using NGS
NGS has many applications in food safety. NGS has been applied to successfully screen fresh produce for the presence of human pathogens such as Salmonella a, allowing for their detection [134]. The limit of detection (LoD) of Salmonella and MS2, a Norovirus surrogate was determined using multiple NGS approaches (16S amplicon sequencing, RNA-seq using ScriptSeq, and NEBNext (New England BioLabs ) kits. ScriptSeq approach was the most sensitive method with a Salmonella LoD of 104 CFU reaction−1 (Colony Forming Units) and the phage MS2 LoD of 105 PFU reaction−1 (Plaque Forming Units) [134]. A review of the literature has shown that L. monocytogenes strains have been identified in foodborne disease outbreaks using WGS [135]. Shiga toxin-producing E. coli have been detected in spiked spinach using shotgun metagenomics with an LoD of ~10 CFU/100 g) [136].
NGS has also been applied after the detection of foodborne bacterial pathogens by other methods such as RT PCR, for confirmatory purposes. Pathogens such as S. sonnei, L. monocytogenes, C. jejuni, S. enterica subsp. enterica serovar enteritidis, S. aureus, E. coli, and Y. enterocolitica have been detected using multiplex PCR-based NGS (Illumina sequencing) from food items such as meat (pork and chicken), milk, seafood, and vegetables [137]. Similarly, NGS (amplicon-based Illumina sequencing) has been successfully applied to amplicons from PCR and RT-PCR of samples from RTE salads, detecting opportunistic pathogens such as Aeromonas hydrophyla and Rahnella aquatilis [138]. NGS in form of whole genome sequencing is especially useful for the surveillance of foodborne pathogens [139].
3.4.2. Detection of Foodborne Fungi Using NGS
3.4.3. Detection of Foodborne Viruses Using NGS
Presently, NGS methods are used as part of a suite of methods for the detection and sequencing of foodborne viruses. This means that non-NGS methods such as RT PCR are first used to detect positive samples before these samples are sequenced with NGS for confirmatory purposes. For example, whole genome sequencing was used to phylogenetically characterize the Avian influenza virus obtained from the chicken and duck meat products brought in by international travelers [63] and samples from Tongzhou poultry meat markets (China) [142]. NGS (metagenomics) was also applied to frozen strawberries in a Norovirus outbreak in Germany in 2012 (alongside other methods such as RT qPCR and dPCR) to conclusively detect and type the genotype of the Norovirus involved in the outbreak [143]. Similarly, NGS (metagenomics) was used to successfully detect Norovirus and or Hepatitis A viruses in RT-PCR-positive samples from virus-spiked celery samples [144], oysters from fish producers in Japan [145] and in RT qPCR-positive lettuce, parsley, and strawberry samples from organic farms in Spain [146]. There have also been instances where the amplicon-based NGS approach has been used to detect and type viral pathogens in food samples. For example, a high diversity of Noroviruses and Hepatitis A viruses was detected in organically grown fresh lettuce, parsley, and strawberry in RT qPCR positive samples using nested PCR targeting NoV and HPV genes with Illumina adapters incorporated into the nested primers [146].
4. Conclusions and Future Directions
A review of four approaches, culture-based, immunoassay, nucleic-acid-based (PCR), and NGS-based methodologies was conducted. Here, culture-based methods when allied with other techniques such as MALDI TOF MS, and nucleic acid-based methods such as PCR (Real-Time PCR for Bacteria and RT-PCR for fungi) are effective at rapidly detecting and identifying different foodborne pathogens, especially those belonging to bacterial and fungal groups. Nucleic acid-based methods such as Real-Time PCR and vPCR combined with sequencing approaches are more widely used than immunoassay and NGS-based approaches for pathogen detection. NGS-based approaches (such as metagenomics) offer unparalleled insights into the genotype, diversity, and activities of foodborne pathogens with their potential for pathogen surveillance, tracing, screening (in a food chain), and identification. These approaches would be extremely useful in the earlier detection, and management of foodborne outbreaks thereby enhancing public safety and health. Current and future directions should be focused on ensuring that existing tools are applied for foodborne pathogen control through training of food surveillance and safety officers on the use of these methods. In addition, more research should be carried out on the development of more effective approaches (such as the use of bacteriophages for the control and elimination of foodborne pathogens.
Funding
This research received no external funding.
Data Availability Statement
No new data were created or analyzed in this study. Data sharing is not applicable to this article.
Acknowledgments
The researcher would like to thank the Deanship of Scientific Research, Qassim University for funding publication of this project.
Conflicts of Interest
The author declares no conflict of interest.
References
- Saucier, L. Microbial spoilage, quality and safety within the context of meat sustainability. Meat Sci. 2016, 120, 78–84. [Google Scholar] [CrossRef] [PubMed]
- Iulietto, M.F.; Sechi, P.; Borgogni, E.; Cenci-Goga, B.T. Meat spoilage: A critical review of a neglected alteration due to ropy slime producing bacteria. Ital. J. Anim. Sci. 2015, 14, 4011. [Google Scholar] [CrossRef]
- Quthama, A.-Z.; Diguță, C.F.; Dopcea, G.; Matei, F. Conventional Versus Modern Techniques Used for the Detection of Pathogens in Food Matrices: A Review. Sci. Bull. Ser. F. Biotechnol. 2022, 26, 79–88. [Google Scholar]
- Mead, P.S.; Slutsker, L.; Dietz, V.; McCaig, L.F.; Bresee, J.S.; Shapiro, C.; Griffin, P.M.; Tauxe, R.V. Food-related illness and death in the United States. Emerg. Infect. Dis. 1999, 5, 607. [Google Scholar] [CrossRef]
- Bintsis, T. Foodborne pathogens. AIMS Microbiol. 2017, 3, 529. [Google Scholar] [CrossRef]
- Martinović, T.; Andjelković, U.; Gajdošik, M.Š.; Rešetar, D.; Josić, D. Foodborne pathogens and their toxins. J. Proteom. 2016, 147, 226–235. [Google Scholar] [CrossRef]
- Lianou, A.; Panagou, E.Z.; Nychas, G.-J.E. Meat safety—I foodborne pathogens and other biological issues. In Lawrie’s Meat Science; Elsevier: Cambridge, UK, 2023; pp. 549–590. [Google Scholar]
- Pexara, A.; Govaris, A. Foodborne viruses and innovative non-thermal food-processing technologies. Foods 2020, 9, 1520. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Y.; Chen, M.; Warner, R.D.; Fang, Z. Incorporating nisin and grape seed extract in chitosan-gelatine edible coating and its effect on cold storage of fresh pork. Food Control 2020, 110, 107018. [Google Scholar] [CrossRef]
- Ribeiro-Santos, R.; Andrade, M.; de Melo, N.R.; Sanches-Silva, A. Use of essential oils in active food packaging: Recent advances and future trends. Trends Food Sci. Technol. 2017, 61, 132–140. [Google Scholar] [CrossRef]
- Batiha, G.E.-S.; Hussein, D.E.; Algammal, A.M.; George, T.T.; Jeandet, P.; Al-Snafi, A.E.; Tiwari, A.; Pagnossa, J.P.; Lima, C.M.; Thorat, N.D. Application of natural antimicrobials in food preservation: Recent views. Food Control 2021, 126, 108066. [Google Scholar] [CrossRef]
- Falleh, H.; Jemaa, M.B.; Saada, M.; Ksouri, R. Essential oils: A promising eco-friendly food preservative. Food Chem. 2020, 330, 127268. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Guo, H.; Zhang, H.; Ren, F.; Zhang, M.; Ge, S.; Luo, H.; Zhao, L. Dynamics of Bacterial Communities of Lamb Meat Packaged in Air and Vacuum Pouch during Chilled Storage. Food Sci. Anim. Resour. 2019, 39, 209. [Google Scholar] [CrossRef] [PubMed]
- Kaur, M.; Shang, H.; Tamplin, M.; Ross, T.; Bowman, J.P. Culture-dependent and culture-independent assessment of spoilage community growth on VP lamb meat from packaging to past end of shelf-life. Food Microbiol. 2017, 68, 71–80. [Google Scholar] [CrossRef] [PubMed]
- Foddai, A.C.; Grant, I.R. Methods for detection of viable foodborne pathogens: Current state-of-art and future prospects. Appl. Microbiol. Biotechnol. 2020, 104, 4281–4288. [Google Scholar] [CrossRef] [PubMed]
- Law, J.W.-F.; Ab Mutalib, N.-S.; Chan, K.-G.; Lee, L.-H. Rapid methods for the detection of foodborne bacterial pathogens: Principles, applications, advantages and limitations. Front. Microbiol. 2015, 5, 770. [Google Scholar] [CrossRef] [PubMed]
- Saravanan, A.; Kumar, P.S.; Hemavathy, R.; Jeevanantham, S.; Kamalesh, R.; Sneha, S.; Yaashikaa, P. Methods of detection of food-borne pathogens: A review. Environ. Chem. Lett. 2021, 19, 189–207. [Google Scholar] [CrossRef]
- Nyachuba, D.G. Foodborne illness: Is it on the rise? Nutr. Rev. 2010, 68, 257–269. [Google Scholar] [CrossRef] [PubMed]
- Ribot, E.M.; Hise, K.B. Future challenges for tracking foodborne diseases. EMBO Rep. 2016, 17, 1499–1505. [Google Scholar] [CrossRef]
- Authority, E.F.S. The European Union One Health 2020 Zoonoses Report. EFSA J. 2021, 19, e06971. [Google Scholar]
- Kirk, M.; Glass, K.; Ford, L.; Brown, K.; Hall, G. Foodborne illness in Australia: Annual incidence circa 2010. Canberra Aust. Gov. Dep. Health 2014, 20, 1857–1864. [Google Scholar]
- Hawksworth, D.L. The magnitude of fungal diversity: The 1· 5 million species estimate revisited. Mycol. Res. 2001, 105, 1422–1432. [Google Scholar] [CrossRef]
- Lee, S.C.; Billmyre, R.B.; Li, A.; Carson, S.; Sykes, S.M.; Huh, E.Y.; Mieczkowski, P.; Ko, D.C.; Cuomo, C.A.; Heitman, J. Analysis of a food-borne fungal pathogen outbreak: Virulence and genome of a Mucor circinelloides isolate from yogurt. MBio 2014, 5, e01390-14. [Google Scholar] [CrossRef]
- Cheng, V.C.; Chan, J.F.; Ngan, A.H.; To, K.K.; Leung, S.; Tsoi, H.; Yam, W.; Tai, J.W.; Wong, S.S.; Tse, H. Outbreak of intestinal infection due to Rhizopus microsporus. J. Clin. Microbiol. 2009, 47, 2834–2843. [Google Scholar] [CrossRef] [PubMed]
- Paterson, R.R.M.; Lima, N. Filamentous fungal human pathogens from food emphasising Aspergillus, Fusarium and Mucor. Microorganisms 2017, 5, 44. [Google Scholar] [CrossRef] [PubMed]
- Benedict, K.; Chiller, T.M.; Mody, R.K. Invasive fungal infections acquired from contaminated food or nutritional supplements: A review of the literature. Foodborne Pathog. Dis. 2016, 13, 343–349. [Google Scholar] [CrossRef]
- Miranda, R.C.; Schaffner, D.W. Virus risk in the food supply chain. Curr. Opin. Food Sci. 2019, 30, 43–48. [Google Scholar] [CrossRef]
- Yeargin, T.; Gibson, K. Key characteristics of foods with an elevated risk for viral enteropathogen contamination. J. Appl. Microbiol. 2019, 126, 996–1010. [Google Scholar] [CrossRef]
- Da Silva, N.; Taniwaki, M.H.; Junqueira, V.C.; Silveira, N.; Okazaki, M.M.; Gomes, R.A.R. Microbiological Examination Methods of Food and Water: A Laboratory Manual; CRC Press: Boca Raton, FL, USA, 2018. [Google Scholar]
- Patil-Joshi, A.; Rangaswamy, B.; Apte-Deshpande, A. based PCR method development, validation and application for microbial detection. J. Genet. Eng. Biotechnol. 2021, 19, 1–10. [Google Scholar] [CrossRef]
- Park, D.G.; Ha, E.S.; Kang, B.; Choi, I.; Kwak, J.E.; Choi, J.; Park, J.; Lee, W.; Kim, S.H.; Kim, S.H.; et al. Development and evaluation of a next-generation Sequencing panel for the multiple detection and identification of pathogens in fermented foods. J. Microbiol. Biotechnol. 2023, 33, 83–95. [Google Scholar] [CrossRef]
- Altayb, H.N.; Badri, R.M.; Chaieb, K.; Moglad, E. Detection and characterization of the most common foodborne pathogens by using multiplex PCR procedure. Saudi J. Biol. Sci. 2023, 1–14. [Google Scholar] [CrossRef]
- Chen, K.; Ma, B.; Li, J.; Chen, E.; Xu, Y.; Yu, X.; Sun, C.; Zhang, M. A rapid and sensitive europium nanoparticle-based lateral flow immunoassay combined with recombinase polymerase amplification for simultaneous detection of three food-borne pathogens. Int. J. Environ. Res. Public Health 2021, 18, 4574. [Google Scholar] [CrossRef] [PubMed]
- Gill, A. The importance of bacterial culture to food microbiology in the age of genomics. Front. Microbiol. 2017, 8, 777. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Lin, C.-W.; Wang, J.; Oh, D.H. Advances in rapid detection methods for foodborne pathogens. J. Microbiol. Biotechnol. 2014, 24, 297–312. [Google Scholar] [CrossRef] [PubMed]
- Tietjen, M.; Fung, D.Y. Salmonellae and food safety. Crit. Rev. Microbiol. 1995, 21, 53–83. [Google Scholar] [CrossRef]
- Oluwaseun, A.C.; Phazang, P.; Sarin, N.B. Biosensors: A fast-growing technology for pathogen detection in agriculture and food sector. In Biosensing Technologies for the Detection of Pathogens—A Prospective Way for Rapid Analysis, 1st ed.; INTECH: Jakarta Selatan, Indonesia, 2018; pp. 37–52. [Google Scholar]
- Wu, J.-L.; Tseng, W.-P.; Lin, C.-H.; Lee, T.-F.; Chung, M.-Y.; Huang, C.-H.; Chen, S.-Y.; Hsueh, P.-R.; Chen, S.-C. Four point-of-care lateral flow immunoassays for diagnosis of COVID-19 and for assessing dynamics of antibody responses to SARS-CoV-2. J. Infect. 2020, 81, 435–442. [Google Scholar] [CrossRef] [PubMed]
- Kumar, K.R.; Cowley, M.J.; Davis, R.L. Next-generation sequencing and emerging technologies. In Seminars in Thrombosis and Hemostasis; Thieme Medical Publishers: Stuttgart, Germany, 2019; pp. 661–673. [Google Scholar]
- McCombie, W.R.; McPherson, J.D.; Mardis, E.R. Next-generation sequencing technologies. Cold Spring Harb. Perspect. Med. 2019, 9, a036798. [Google Scholar] [CrossRef]
- Biswas, S.; Rolain, J.-M. Use of MALDI-TOF mass spectrometry for identification of bacteria that are difficult to culture. J. Microbiol. Methods 2013, 92, 14–24. [Google Scholar] [CrossRef]
- Rychert, J. Benefits and Limitations of MALDI-TOF Mass Spectrometry for the Identification of Microorganisms. J. Infect. Epidemiol. 2019, 2, 4. [Google Scholar] [CrossRef]
- Singhal, N.; Kumar, M.; Kanaujia, P.K.; Virdi, J.S. MALDI-TOF mass spectrometry: An emerging technology for microbial identification and diagnosis. Front. Microbiol. 2015, 6, 791. [Google Scholar] [CrossRef]
- Shafini, A.; Son, R.; Mahyudin, N.; Rukayadi, Y.; Zainazor, T.T. Prevalence of Salmonella spp. in chicken and beef from retail outlets in Malaysia. Int. Food Res. J. 2017, 24, 437. [Google Scholar]
- Elbehiry, A.; Marzouk, E.; Hamada, M.; Al-Dubaib, M.; Alyamani, E.; Moussa, I.M.; AlRowaidhan, A.; Hemeg, H.A. Application of MALDI-TOF MS fingerprinting as a quick tool for identification and clustering of foodborne pathogens isolated from food products. New Microbiol. 2017, 40, 269–278. [Google Scholar] [PubMed]
- Khater, D.F.; Lela, R.A.; El-Diasty, M.; Moustafa, S.A.; Wareth, G. Detection of harmful foodborne pathogens in food samples at the points of sale by MALDT-TOF MS in Egypt. BMC Res. Notes 2021, 14, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Liu, G.; Huang, X.; Li, L.; Lin, H.; Xu, D. MALDI-TOF MS-Based identification of bacteria and a survey of fresh vegetables with pathogenic bacteria in Beijing, China. Food Biosci. 2021, 41, 100746. [Google Scholar] [CrossRef]
- Cebeci, T. A Survey of Raw Milk For Microbiological Quality and Typing of Foodborne Pathogens by MALDI-TOF MS. Adnan Menderes Üniversitesi Ziraat Fakültesi Derg. 2019, 16, 185–191. [Google Scholar] [CrossRef]
- Sulaiman, I.M.; Miranda, N.; Simpson, S. MALDI-TOF mass spectrometry and 16S rRNA gene sequence analysis for the identification of foodborne clostridium spp. J. AOAC Int. 2021, 104, 1381–1388. [Google Scholar] [CrossRef]
- Quéro, L.; Girard, V.; Pawtowski, A.; Tréguer, S.; Weill, A.; Arend, S.; Cellière, B.; Polsinelli, S.; Monnin, V.; van Belkum, A. Development and application of MALDI-TOF MS for identification of food spoilage fungi. Food Microbiol. 2019, 81, 76–88. [Google Scholar] [CrossRef]
- Pattabhiramaiah, M.; Mallikarjunaiah, S. High-Throughput Sequencing for Detection of Foodborne Pathogens in Food Safety. In Sequencing Technologies in Microbial Food Safety and Quality; CRC Press: Boca Raton, FL, USA, 2021; pp. 393–424. [Google Scholar]
- Hasan, N.; Zanuddin, N. Molecular identification of isolated fungi from banana, mango and pineapple spoiled fruits. In Proceedings of the AIP Conference Proceedings, Maharashtra, India, 5–6 July 2018; p. 020074. [Google Scholar]
- Quintilla, R.; Kolecka, A.; Casaregola, S.; Daniel, H.M.; Houbraken, J.; Kostrzewa, M.; Boekhout, T.; Groenewald, M. MALDI-TOF MS as a tool to identify foodborne yeasts and yeast-like fungi. Int. J. Food Microbiol. 2018, 266, 109–118. [Google Scholar] [CrossRef]
- Ahmadsah, L.S.; Kim, E.; Jung, Y.-S.; Kim, H.-Y. Identification of LAB and fungi in Laru, a fermentation starter, by PCR-DGGE, SDS-PAGE, and MALDI-TOF MS. J. Microbiol. Biotechnol. 2018, 28, 32–39. [Google Scholar] [CrossRef]
- Bader, O. Fungal species identification by MALDI-ToF mass spectrometry. Hum. Fungal Pathog. Identif. Methods Protoc. 2017, 1508, 323–337. [Google Scholar]
- Su, W.; Liang, D.; Tan, M. Nucleic acid-based detection for foodborne virus utilizing microfluidic systems. Trends Food Sci. Technol. 2021, 113, 97–109. [Google Scholar] [CrossRef]
- Bosch, A.; Sánchez, G.; Abbaszadegan, M.; Carducci, A.; Guix, S.; Le Guyader, F.S.; Netshikweta, R.; Pintó, R.M.; Van Der Poel, W.H.; Rutjes, S. Analytical methods for virus detection in water and food. Food Anal. Methods 2011, 4, 4–12. [Google Scholar] [CrossRef]
- Chhabra, P.; Vinjé, J. Molecular detection methods of foodborne viruses. In Viruses in Foods; Springer: London, UK, 2016; pp. 303–333. [Google Scholar]
- Sánchez, G.; Bosch, A. Survival of enteric viruses in the environment and food. Viruses Foods 2016, 367–392. [Google Scholar] [CrossRef]
- Dai, M.; Yan, N.; Huang, Y.; Zhao, L.; Liao, M. Survivability of highly pathogenic avian influenza virus on raw chicken meat in different environmental conditions. Lancet Microbe 2022, 3, e92. [Google Scholar] [CrossRef]
- Zhang, J.; Li, X.; Wang, X.; Ye, H.; Li, B.; Chen, Y.; Chen, J.; Zhang, T.; Qiu, Z.; Li, H. Genomic evolution, transmission dynamics, and pathogenicity of avian influenza A (H5N8) viruses emerging in China, 2020. Virus Evol. 2021, 7, veab046. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Mitake, H.; Kiso, M.; Ito, M.; Iwatsuki-Hirimoto, K.; Yamayoshi, S.; Lopes, T.J.; Feng, H.; Sumiyoshi, R.; Shibata, A. Characterization of H7N9 avian influenza viruses isolated from duck meat products. Transbound. Emerg. Dis. 2020, 67, 792–798. [Google Scholar] [CrossRef]
- Shibata, A.; Okamatsu, M.; Sumiyoshi, R.; Matsuno, K.; Wang, Z.-J.; Kida, H.; Osaka, H.; Sakoda, Y. Repeated detection of H7N9 avian influenza viruses in raw poultry meat illegally brought to Japan by international flight passengers. Virology 2018, 524, 10–17. [Google Scholar] [CrossRef]
- Marti, E.; Barardi, C.R.M. Detection of human adenoviruses in organic fresh produce using molecular and cell culture-based methods. Int. J. Food Microbiol. 2016, 230, 40–44. [Google Scholar] [CrossRef]
- Priyanka, B.; Patil, R.K.; Dwarakanath, S. A review on detection methods used for foodborne pathogens. Indian J. Med. Res. 2016, 144, 327. [Google Scholar] [CrossRef]
- Agriopoulou, S.; Stamatelopoulou, E.; Varzakas, T. Advances in analysis and detection of major mycotoxins in foods. Foods 2020, 9, 518. [Google Scholar] [CrossRef]
- Omori, A.M.; Ono, E.Y.S.; Bordini, J.G.; Hirozawa, M.T.; Fungaro, M.H.P.; Ono, M.A. Detection of Fusarium verticillioides by PCR-ELISA based on FUM21 gene. Food Microbiol. 2018, 73, 160–167. [Google Scholar] [CrossRef]
- Di Febo, T.; Schirone, M.; Visciano, P.; Portanti, O.; Armillotta, G.; Persiani, T.; Di Giannatale, E.; Tittarelli, M.; Luciani, M. Development of a capture ELISA for rapid detection of Salmonella enterica in food samples. Food Anal. Methods 2019, 12, 322–330. [Google Scholar] [CrossRef]
- Wang, S.; Zhang, Y.; An, W.; Wei, Y.; Liu, N.; Chen, Y.; Shuang, S. Magnetic relaxation switch immunosensor for the rapid detection of the foodborne pathogen Salmonella enterica in milk samples. Food Control 2015, 55, 43–48. [Google Scholar] [CrossRef]
- Khan, J.A.; Rathore, R.S.; Abulreesh, H.H.; Qais, F.A.; Ahmad, I. Cultural and immunological methods for the detection of Campylobacter jejuni: A review. Indian J. Biotechnol. Pharm. Res. 2018, 6, 4–10. [Google Scholar]
- Pang, B.; Zhao, C.; Li, L.; Song, X.; Xu, K.; Wang, J.; Liu, Y.; Fu, K.; Bao, H.; Song, D. Development of a low-cost paper-based ELISA method for rapid Escherichia coli O157: H7 detection. Anal. Biochem. 2018, 542, 58–62. [Google Scholar] [CrossRef]
- Zhao, Y.; Zeng, D.; Yan, C.; Chen, W.; Ren, J.; Jiang, Y.; Jiang, L.; Xue, F.; Ji, D.; Tang, F. Rapid and accurate detection of Escherichia coli O157: H7 in beef using microfluidic wax-printed paper-based ELISA. Analyst 2020, 145, 3106–3115. [Google Scholar] [CrossRef]
- Kumar, B.K.; Raghunath, P.; Devegowda, D.; Deekshit, V.K.; Venugopal, M.N.; Karunasagar, I.; Karunasagar, I. Development of monoclonal antibody based sandwich ELISA for the rapid detection of pathogenic Vibrio parahaemolyticus in seafood. Int. J. Food Microbiol. 2011, 145, 244–249. [Google Scholar] [CrossRef]
- Tu, Z.; Chen, Q.; Li, Y.; Xiong, Y.; Xu, Y.; Hu, N.; Tao, Y. Identification and characterization of species-specific nanobodies for the detection of Listeria monocytogenes in milk. Anal. Biochem. 2016, 493, 1–7. [Google Scholar] [CrossRef]
- Nagaraj, S.; Ramlal, S.; Kingston, J.; Batra, H.V. Development of IgY based sandwich ELISA for the detection of staphylococcal enterotoxin G (SEG), an egc toxin. Int. J. Food Microbiol. 2016, 237, 136–141. [Google Scholar] [CrossRef]
- Stanker, L.H.; Scotcher, M.C.; Cheng, L.; Ching, K.; McGarvey, J.; Hodge, D.; Hnasko, R. A monoclonal antibody based capture ELISA for botulinum neurotoxin serotype B: Toxin detection in food. Toxins 2013, 5, 2212–2226. [Google Scholar] [CrossRef]
- Valderrama, W.B.; Dudley, E.G.; Doores, S.; Cutter, C.N. Commercially available rapid methods for detection of selected food-borne pathogens. Crit. Rev. Food Sci. Nutr. 2016, 56, 1519–1531. [Google Scholar] [CrossRef]
- Rajkovic, A.; Jovanovic, J.; Monteiro, S.; Decleer, M.; Andjelkovic, M.; Foubert, A.; Beloglazova, N.; Tsilla, V.; Sas, B.; Madder, A. Detection of toxins involved in foodborne diseases caused by Gram-positive bacteria. Compr. Rev. Food Sci. Food Saf. 2020, 19, 1605–1657. [Google Scholar] [CrossRef] [PubMed]
- Hassan, F.F.; Al-Jibouri, M.H.; Hashim, A.K.J. Isolation and Identification of Fungal Propagation in Stored Maize and detection of aflatoxin B1 Using TLC and ELISA Technique. Iraqi J. Sci. 2014, 55, 634–642. [Google Scholar]
- Ounleye, A.O.; Olaiya, G.A. Isolation, Identification and Mycotoxin Production of Some Mycoflora of Dried Stockfish (Gadus morhua). Acad. J. Sci 2015, 4, 345–363. [Google Scholar]
- Oplatowska-Stachowiak, M.; Sajic, N.; Xu, Y.; Haughey, S.A.; Mooney, M.H.; Gong, Y.Y.; Verheijen, R.; Elliott, C.T. Fast and sensitive aflatoxin B1 and total aflatoxins ELISAs for analysis of peanuts, maize and feed ingredients. Food Control 2016, 63, 239–245. [Google Scholar] [CrossRef]
- Beley, M.A.J.; Teves, F.G.; Reina, M.; Madamba, S. Isolation of fungal species and detection of aflatoxin from soy milk products using ELISA method. Int. Res. J. Biol. Sci 2013, 2, 45–48. [Google Scholar]
- Kim, H.; Chung, D.-R.; Kang, M. A new point-of-care test for the diagnosis of infectious diseases based on multiplex lateral flow immunoassays. Analyst 2019, 144, 2460–2466. [Google Scholar] [CrossRef]
- Koopmans, M.; Duizer, E. Foodborne viruses: An emerging problem. Int. J. Food Microbiol. 2004, 90, 23–41. [Google Scholar] [CrossRef]
- Appleton, H. Control of food-borne viruses. Br. Med. Bull. 2000, 56, 172–183. [Google Scholar] [CrossRef]
- Singh, J.; Birbian, N.; Sinha, S.; Goswami, A. A critical review on PCR, its types and applications. Int. J. Adv. Res. Biol. Sci. 2014, 1, 65–80. [Google Scholar]
- Muhamad Rizal, N.S.; Neoh, H.-m.; Ramli, R.; A/LK Periyasamy, P.R.; Hanafiah, A.; Abdul Samat, M.N.; Tan, T.L.; Wong, K.K.; Nathan, S.; Chieng, S. Advantages and limitations of 16S rRNA next-generation sequencing for pathogen identification in the diagnostic microbiology laboratory: Perspectives from a middle-income country. Diagnostics 2020, 10, 816. [Google Scholar] [CrossRef]
- Mancini, V.; Murolo, S.; Romanazzi, G. Diagnostic methods for detecting fungal pathogens on vegetable seeds. Plant Pathol. 2016, 65, 691–703. [Google Scholar] [CrossRef]
- Olmedillas-López, S.; Olivera-Salazar, R.; García-Arranz, M.; García-Olmo, D. Current and emerging applications of droplet digital PCR in oncology: An updated review. Mol. Diagn. Ther. 2021, 21, 493–510. [Google Scholar] [CrossRef] [PubMed]
- Yang, R.; Paparini, A.; Monis, P.; Ryan, U. Comparison of next-generation droplet digital PCR (ddPCR) with quantitative PCR (qPCR) for enumeration of Cryptosporidium oocysts in faecal samples. Int. J. Parasitol. 2014, 44, 1105–1113. [Google Scholar] [CrossRef]
- Soroka, M.; Wasowicz, B.; Rymaszewska, A. Loop-Mediated Isothermal Amplification (LAMP): The Better Sibling of PCR? Cells 2021, 10, 1931. [Google Scholar] [CrossRef] [PubMed]
- Bosch, A.; Pintó, R.M.; Guix, S. Foodborne viruses. Curr. Opin. Food Sci. 2016, 8, 110–119. [Google Scholar] [CrossRef]
- Lee, N.; Kwon, K.Y.; Oh, S.K.; Chang, H.-J.; Chun, H.S.; Choi, S.-W. A multiplex PCR assay for simultaneous detection of Escherichia coli O157: H7, Bacillus cereus, Vibrio parahaemolyticus, Salmonella spp., Listeria monocytogenes, and Staphylococcus aureus in Korean ready-to-eat food. Foodborne Pathog. Dis. 2014, 11, 574–580. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, R. Escherichia coli O157: H7 identification in Minas Frescalcheesecommercialized in Goiânia. GO. Braz. 2014, 45, 7–10. [Google Scholar]
- Moezi, P.; Kargar, M.; Doosti, A.; Khoshneviszadeh, M. Multiplex touchdown PCR assay to enhance specificity and sensitivity for concurrent detection of four foodborne pathogens in raw milk. J. Appl. Microbiol. 2019, 127, 262–273. [Google Scholar] [CrossRef]
- Samad, A.; Abbas, F.; Ahmad, Z.; Tanveer, Z.; Ahmad, I.; Patching, S.G.; Nawaz, N.; Asmat, M.T.; Raziq, A.; Naeem, M. Multiplex polymerase chain reaction detection of Shiga toxin genes and antibiotic sensitivity of Escherichia coli O157: H7 isolated from beef meat in Quetta, Pakistan. J. Food Saf. 2018, 38, e12540. [Google Scholar] [CrossRef]
- Vinayaka, A.C.; Ngo, T.A.; Kant, K.; Engelsmann, P.; Dave, V.P.; Shahbazi, M.-A.; Wolff, A.; Bang, D.D. Rapid detection of Salmonella enterica in food samples by a novel approach with combination of sample concentration and direct PCR. Biosens. Bioelectron. 2019, 129, 224–230. [Google Scholar] [CrossRef]
- Pires, N.M.; Dong, T.; Yang, Z.; da Silva, L.F. Recent methods and biosensors for foodborne pathogen detection in fish: Progress and future prospects to sustainable aquaculture systems. Crit. Rev. Food Sci. Nutr. 2021, 61, 1852–1876. [Google Scholar] [CrossRef] [PubMed]
- Bai, X.; Wang, Z.; Li, W.; Xiao, F.; Huang, J.; Xu, Q.; Xu, H. Rapid and accurate detection for Listeria monocytogenes in milk using ampicillin-mediated magnetic separation coupled with quantitative real-time PCR. Microchem. J. 2022, 183, 108063. [Google Scholar] [CrossRef]
- Chin, N.A.; Salihah, N.T.; Shivanand, P.; Ahmed, M.U. Recent trends and developments of PCR-based methods for the detection of food-borne Salmonella bacteria and Norovirus. J. Food Sci. Technol. 2021, 59, 4570–4582. [Google Scholar] [CrossRef]
- Heo, E.J.; Kim, H.-Y.; Suh, S.H.; MOON, J.S. Comparison of DNA Extraction Methods for the Quantification of Listeria monocytogenes in Dairy Products by Real-Time Quantitative PCR. J. Food Prot. 2022, 85, 1531–1537. [Google Scholar] [CrossRef]
- Azimirad, M.; Nadalian, B.; Alavifard, H.; Panirani, S.N.; Bonab, S.M.V.; Azimirad, F.; Gholami, F.; Jabbari, P.; Yadegar, A.; Busani, L. Microbiological survey and occurrence of bacterial foodborne pathogens in raw and ready-to-eat green leafy vegetables marketed in Tehran, Iran. Int. J. Hyg. Environ. Health 2021, 237, 113824. [Google Scholar] [CrossRef] [PubMed]
- Cao, X.; Zhao, L.; Zhang, J.; Chen, X.; Shi, L.; Fang, X.; Xie, H.; Chang, Y.; Wang, L. Detection of viable but nonculturable Vibrio parahaemolyticus in shrimp samples using improved real-time PCR and real-time LAMP methods. Food Control 2019, 103, 145–152. [Google Scholar] [CrossRef]
- Bonnin-Jusserand, M.; Copin, S.; Le Bris, C.; Brauge, T.; Gay, M.; Brisabois, A.; Grard, T.; Midelet-Bourdin, G. Vibrio species involved in seafood-borne outbreaks (Vibrio cholerae, V. parahaemolyticus and V. vulnificus): Review of microbiological versus recent molecular detection methods in seafood products. Crit. Rev. Food Sci. Nutr. 2019, 59, 597–610. [Google Scholar] [CrossRef]
- Yang, X.; Zhang, X.; Wang, Y.; Shen, H.; Jiang, G.; Dong, J.; Zhao, P.; Gao, S. A real-time recombinase polymerase amplification method for rapid detection of Vibrio vulnificus in seafood. Front. Microbiol. 2020, 11, 586981. [Google Scholar] [CrossRef]
- Xiao, L.; Zhang, L.; Wang, H.H. Critical issues in detecting viable Listeria monocytogenes cells by real-time reverse transcriptase PCR. J. Food Prot. 2012, 75, 512–517. [Google Scholar] [CrossRef]
- Miller, N.D.; Draughon, F.A.; D’Souza, D.H. Real-time reverse-transcriptase–polymerase chain reaction for Salmonella enterica detection from jalapeno and serrano peppers. Foodborne Pathog. Dis. 2010, 7, 367–373. [Google Scholar] [CrossRef]
- Zhou, M.; Yang, J.; Zhou, X.; Liu, B.; Liu, D.; Yuan, C.; He, Y.; Pan, L.; Shi, X. Development of a sigDE-based real-time reverse-transcriptase PCR for the detection of viable Salmonella enterica. Foodborne Pathog. Dis. 2014, 11, 537–544. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Lan, X.; Zhu, L.; Ru, P.; Xu, W.; Liu, H. PCR Mediated Nucleic Acid Molecular Recognition Technology for Detection of Viable and Dead Foodborne Pathogens. Foods 2022, 11, 2675. [Google Scholar] [CrossRef] [PubMed]
- Dinh Thanh, M.; Agustí, G.; Mader, A.; Appel, B.; Codony, F. Improved sample treatment protocol for accurate detection of live Salmonella spp. in food samples by viability PCR. PLoS ONE 2017, 12, e0189302. [Google Scholar] [CrossRef]
- De Cooman, L.; Flahou, B.; Houf, K.; Smet, A.; Ducatelle, R.; Pasmans, F.; Haesebrouck, F. Survival of Helicobacter suis bacteria in retail pig meat. Int. J. Food Microbiol. 2013, 166, 164–167. [Google Scholar] [CrossRef] [PubMed]
- Castro, A.G.; Dorneles, E.M.; Santos, E.L.; Alves, T.M.; Silva, G.R.; Figueiredo, T.C.; Assis, D.C.; Lage, A.P.; Cançado, S.V. Viability of Campylobacter spp. in frozen and chilled broiler carcasses according to real-time PCR with propidium monoazide pretreatment. Poult. Sci. 2018, 97, 1706–1711. [Google Scholar] [CrossRef] [PubMed]
- Agustí, G.; Fittipaldi, M.; Codony, F. Optimization of a viability PCR method for the detection of Listeria monocytogenes in food samples. Curr. Microbiol. 2018, 75, 779–785. [Google Scholar] [CrossRef] [PubMed]
- Sexton, D.J.; Kordalewska, M.; Bentz, M.L.; Welsh, R.M.; Perlin, D.S.; Litvintseva, A.P. Direct detection of emergent fungal pathogen Candida auris in clinical skin swabs by SYBR green-based quantitative PCR assay. J. Clin. Microbiol. 2018, 56, e01337-18. [Google Scholar] [CrossRef]
- Wagner, K.; Springer, B.; Pires, V.; Keller, P.M. Molecular detection of fungal pathogens in clinical specimens by 18S rDNA high-throughput screening in comparison to ITS PCR and culture. Sci. Rep. 2018, 8, 6964. [Google Scholar] [CrossRef]
- Luchi, N.; Ioos, R.; Santini, A. Fast and reliable molecular methods to detect fungal pathogens in woody plants. Appl. Microbiol. Biotechnol. 2020, 104, 2453–2468. [Google Scholar] [CrossRef]
- Vergidis, P.; Moore, C.B.; Novak-Frazer, L.; Rautemaa-Richardson, R.; Walker, A.; Denning, D.W.; Richardson, M.D. High-volume culture and quantitative real-time PCR for the detection of Aspergillus in sputum. Clin. Microbiol. Infect. 2020, 26, 935–940. [Google Scholar] [CrossRef]
- Rahman, H.U.; Yue, X.; Yu, Q.; Zhang, W.; Zhang, Q.; Li, P. Current PCR-based methods for the detection of mycotoxigenic fungi in complex food and feed matrices. World Mycotoxin J. 2020, 13, 139–150. [Google Scholar] [CrossRef]
- Kim, D.M.; Chung, S.H.; Chun, H.S. Multiplex PCR assay for the detection of aflatoxigenic and non-aflatoxigenic fungi in meju, a Korean fermented soybean food starter. Food Microbiol. 2011, 28, 1402–1408. [Google Scholar] [CrossRef]
- Rahman, H.U.; Yue, X.; Ren, X.; Zhang, W.; Zhang, Q.; Li, P. Multiplex PCR assay to detect Aspergillus, Penicillium and Fusarium species simultaneously. Food Addit. Contam. Part A 2020, 37, 1939–1950. [Google Scholar] [CrossRef] [PubMed]
- Tannous, J.; Atoui, A.; El Khoury, A.; Kantar, S.; Chdid, N.; Oswald, I.P.; Puel, O.; Lteif, R. Development of a real-time PCR assay for Penicillium expansum quantification and patulin estimation in apples. Food Microbiol. 2015, 50, 28–37. [Google Scholar] [CrossRef] [PubMed]
- Ferrara, M.; Perrone, G.; Gallo, A.; Epifani, F.; Visconti, A.; Susca, A. Development of loop-mediated isothermal amplification (LAMP) assay for the rapid detection of Penicillium nordicum in dry-cured meat products. Int. J. Food Microbiol. 2015, 202, 42–47. [Google Scholar] [CrossRef]
- Laforgue, R.; Guérin, L.; Pernelle, J.; Monnet, C.; Dupont, J.; Bouix, M. Evaluation of PCR-DGGE methodology to monitor fungal communities on grapes. J. Appl. Microbiol. 2009, 107, 1208–1218. [Google Scholar] [CrossRef]
- Coudray-Meunier, C.; Fraisse, A.; Martin-Latil, S.; Guillier, L.; Perelle, S. Discrimination of infectious hepatitis A virus and rotavirus by combining dyes and surfactants with RT-qPCR. BMC Microbiol. 2013, 13, 216. [Google Scholar] [CrossRef]
- La Bella, G.; Martella, V.; Basanisi, M.; Nobili, G.; Terio, V.; La Salandra, G. Food-borne viruses in shellfish: Investigation on norovirus and HAV presence in Apulia (SE Italy). Food Environ. Virol. 2017, 9, 179–186. [Google Scholar] [CrossRef] [PubMed]
- Althof, N.; Trojnar, E.; Böhm, T.; Burkhardt, S.; Carl, A.; Contzen, M.; Kilwinski, J.; Mergemeier, S.; Moor, D.; Mäde, D. Interlaboratory validation of a method for hepatitis E virus RNA detection in meat and meat products. Food Environ. Virol. 2019, 11, 1–8. [Google Scholar] [CrossRef]
- Szabo, K.; Trojnar, E.; Anheyer-Behmenburg, H.; Binder, A.; Schotte, U.; Ellerbroek, L.; Klein, G.; Johne, R. Detection of hepatitis E virus RNA in raw sausages and liver sausages from retail in Germany using an optimized method. Int. J. Food Microbiol. 2015, 215, 149–156. [Google Scholar] [CrossRef]
- Soares, V.M.; Dos Santos, E.A.R.; Tadielo, L.E.; Cerqueira-Cézar, C.K.; Sampaio, A.N.d.C.E.; Eisen, A.K.A.; de Oliveira, K.G.; Padilha, M.B.; de Moraes Guerra, M.E.; Gasparetto, R. Detection of adenovirus, rotavirus, and hepatitis E virus in meat cuts marketed in Uruguaiana, Rio Grande do Sul, Brazil. One Health 2022, 14, 100377. [Google Scholar] [CrossRef]
- Fraisse, A.; Coudray-Meunier, C.; Martin-Latil, S.; Hennechart-Collette, C.; Delannoy, S.; Fach, P.; Perelle, S. Digital RT-PCR method for hepatitis A virus and norovirus quantification in soft berries. Int. J. Food Microbiol. 2017, 243, 36–45. [Google Scholar] [CrossRef] [PubMed]
- Coudray-Meunier, C.; Fraisse, A.; Martin-Latil, S.; Guillier, L.; Delannoy, S.; Fach, P.; Perelle, S. A comparative study of digital RT-PCR and RT-qPCR for quantification of Hepatitis A virus and Norovirus in lettuce and water samples. Int. J. Food Microbiol. 2015, 201, 17–26. [Google Scholar] [CrossRef] [PubMed]
- Jagadeesan, B.; Gerner-Smidt, P.; Allard, M.W.; Leuillet, S.; Winkler, A.; Xiao, Y.; Chaffron, S.; Van Der Vossen, J.; Tang, S.; Katase, M. The use of next generation sequencing for improving food safety: Translation into practice. Food Microbiol. 2019, 79, 96–115. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Li, Y.; Li, S.; Hu, N.; He, Y.; Pong, R.; Lin, D.; Lu, L.; Law, M. Comparison of next-generation sequencing systems. J. Biomed. Biotechnol. 2012, 2012, 251364. [Google Scholar] [CrossRef] [PubMed]
- Desdouits, M.; de Graaf, M.; Strubbia, S.; Oude Munnink, B.B.; Kroneman, A.; Le Guyader, F.S.; Koopmans, M.P. Novel opportunities for NGS-based one health surveillance of foodborne viruses. One Health Outlook 2020, 2, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Lewis, E.; Hudson, J.; Cook, N.; Barnes, J.; Haynes, E. Next-generation sequencing as a screening tool for foodborne pathogens in fresh produce. J. Microbiol. Methods 2020, 171, 105840. [Google Scholar] [CrossRef]
- Law, J.W.-F.; Ab Mutalib, N.-S.; Chan, K.-G.; Lee, L.-H. An insight into the isolation, enumeration, and molecular detection of Listeria monocytogenes in food. Front. Microbiol. 2015, 6, 1227. [Google Scholar] [CrossRef]
- Leonard, S.R.; Mammel, M.K.; Lacher, D.W.; Elkins, C.A. Application of metagenomic sequencing to food safety: Detection of Shiga toxin-producing Escherichia coli on fresh bagged spinach. Appl. Environ. Microbiol. 2015, 81, 8183–8191. [Google Scholar] [CrossRef]
- Ferrario, C.; Lugli, G.A.; Ossiprandi, M.C.; Turroni, F.; Milani, C.; Duranti, S.; Mancabelli, L.; Mangifesta, M.; Alessandri, G.; van Sinderen, D. Next generation sequencing-based multigene panel for high throughput detection of food-borne pathogens. Int. J. Food Microbiol. 2017, 256, 20–29. [Google Scholar] [CrossRef]
- Mira Miralles, M.; Maestre-Carballa, L.; Lluesma-Gomez, M.; Martinez-Garcia, M. High-throughput 16S rRNA sequencing to assess potentially active bacteria and foodborne pathogens: A case example in ready-to-eat food. Foods 2019, 8, 480. [Google Scholar] [CrossRef] [PubMed]
- Moran-Gilad, J. Whole genome sequencing (WGS) for food-borne pathogen surveillance and control–taking the pulse. Eurosurveillance 2017, 22, 30547. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.; Chen, Y.; Han, S.; Lv, L.; Li, L. Next-Generation Sequencing Applications for the Study of Fungal Pathogens. Microorganisms 2022, 10, 1882. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, A.E.; Rossoff, J.; Hollemon, D.; Hong, D.K.; Muller, W.J.; Chaudhury, S. Cell-free DNA next-generation sequencing successfully detects infectious pathogens in pediatric oncology and hematopoietic stem cell transplant patients at risk for invasive fungal disease. Pediatr. Blood Cancer 2019, 66, e27734. [Google Scholar] [CrossRef]
- Cui, S.; Zou, L.; Liu, Y.; Zhao, J.; Lu, G.; Shi, W.; Pan, Y.; Peng, X.; Zhang, D.; Yang, P. Detection by environmental surveillance and genomic characterization of H5N8 highly pathogenic avian influenza virus from a poultry meat market in Beijing, China, 2021–2022. Vet. Res. Commun. 2022, 46, 1369–1375. [Google Scholar] [CrossRef]
- Bartsch, C.; Höper, D.; Mäde, D.; Johne, R. Analysis of frozen strawberries involved in a large norovirus gastroenteritis outbreak using next generation sequencing and digital PCR. Food Microbiol. 2018, 76, 390–395. [Google Scholar] [CrossRef]
- Yang, Z.; Mammel, M.; Papafragkou, E.; Hida, K.; Elkins, C.A.; Kulka, M. Application of next generation sequencing toward sensitive detection of enteric viruses isolated from celery samples as an example of produce. Int. J. Food Microbiol. 2017, 261, 73–81. [Google Scholar] [CrossRef]
- Imamura, S.; Kanezashi, H.; Goshima, T.; Haruna, M.; Okada, T.; Inagaki, N.; Uema, M.; Noda, M.; Akimoto, K. Next-generation sequencing analysis of the diversity of human noroviruses in Japanese oysters. Foodborne Pathog. Dis. 2017, 14, 465–471. [Google Scholar] [CrossRef]
- Itarte, M.; Martínez-Puchol, S.; Forés, E.; Hundesa, A.; Timoneda, N.; Bofill-Mas, S.; Girones, R.; Rusiñol, M. NGS techniques reveal a high diversity of RNA viral pathogens and papillomaviruses in fresh produce and irrigation water. Foods 2021, 10, 1820. [Google Scholar] [CrossRef]
Figure 1.
Nucleic acid-based methods for detecting and identifying foodborne pathogens. Adapted from [92].
Figure 1.
Nucleic acid-based methods for detecting and identifying foodborne pathogens. Adapted from [92].

Figure 2.
Next-Generation Sequencing approach for detecting and identifying foodborne pathogens. Adapted from [32,133].


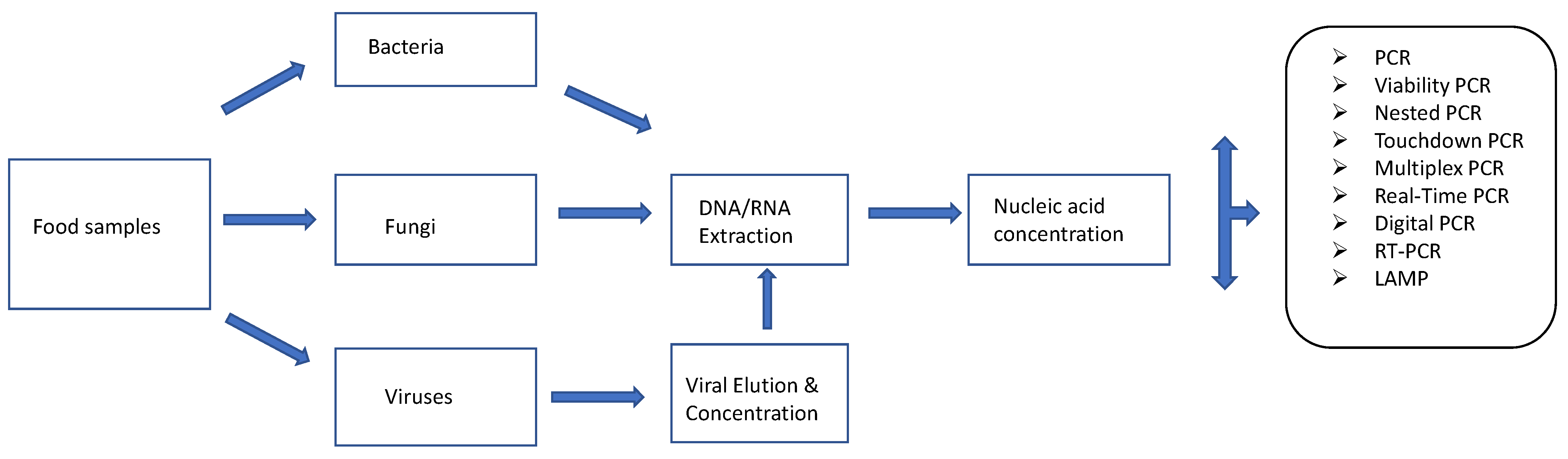{kind=link}
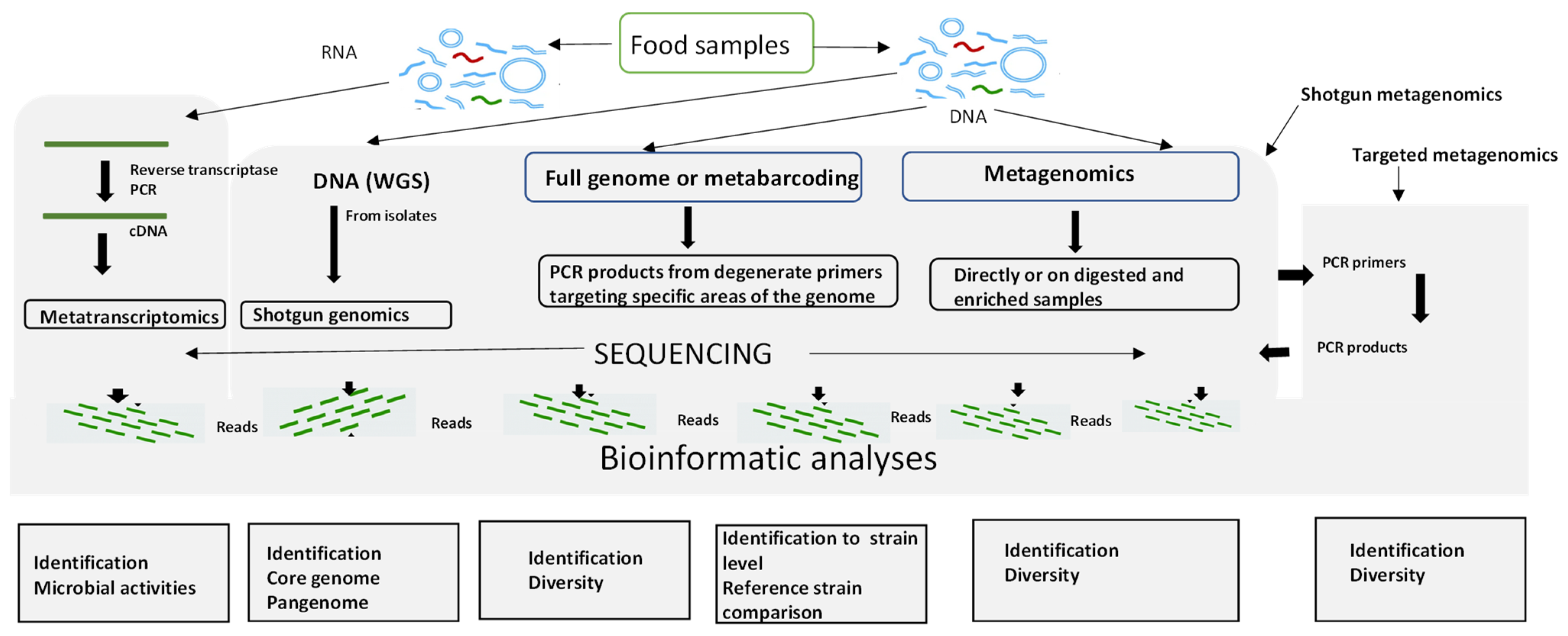{kind=link}
Table 1.
Methods for foodborne pathogen detection.
| Method | Target Groups | Principle | Advantages | Limitations |
|---|---|---|---|---|
| Culture Based Methods | Bacteria, fungi, and viruses | Use of culture media containing the growth requirements of target microorganisms |
|
|
| Immunological assays (ELISA and LFD) | Bacteria, fungi, and viruses | The affinity between microbial antigens and monoclonal or polyclonal antibodies is exploited for the detection of microorganisms |
|
|
| Conventional PCR (e.g., nested, touchdown, multiplex, etc.) | Bacteria, fungi, and viruses | Based on the use of primers targeting specific regions of microbial DNA. Target when present is exponentially amplified |
|
|
| Viability PCR/qPCR | Bacteria | Special dyes used in the test render non-viable bacterial DNA non-amplifiable |
|
|
| Reverse Transcriptase (RT) PCR/qPCR | Bacteria, fungi, and viruses | mRNA transcripts level declines quickly after cell death. Detected mRNA should be from viable cells and is therefore targeted (RT-PCR). Transcripts are amplified and quantified using dyes or probes |
|
|
| Next Generation Sequencing Approaches | Bacteria, fungi, and viruses | Whole genome or markers such as ITS and 16S rRNA are targeted and sequenced (random shotgun sequencing, gene/marker specific sequencing, etc.) |
|
|
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Aladhadh, M. A Review of Modern Methods for the Detection of Foodborne Pathogens. Microorganisms 2023, 11, 1111. https://doi.org/10.3390/microorganisms11051111
AMA Style
Aladhadh M. A Review of Modern Methods for the Detection of Foodborne Pathogens. Microorganisms. 2023; 11(5):1111. https://doi.org/10.3390/microorganisms11051111
Chicago/Turabian StyleAladhadh, Mohammed. 2023. "A Review of Modern Methods for the Detection of Foodborne Pathogens" Microorganisms 11, no. 5: 1111. https://doi.org/10.3390/microorganisms11051111
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.