
Antimicrobial Drug Resistance in Poultry Production: Current Status and Innovative Strategies for Bacterial Control
 , , ,
, , ,
Abstract
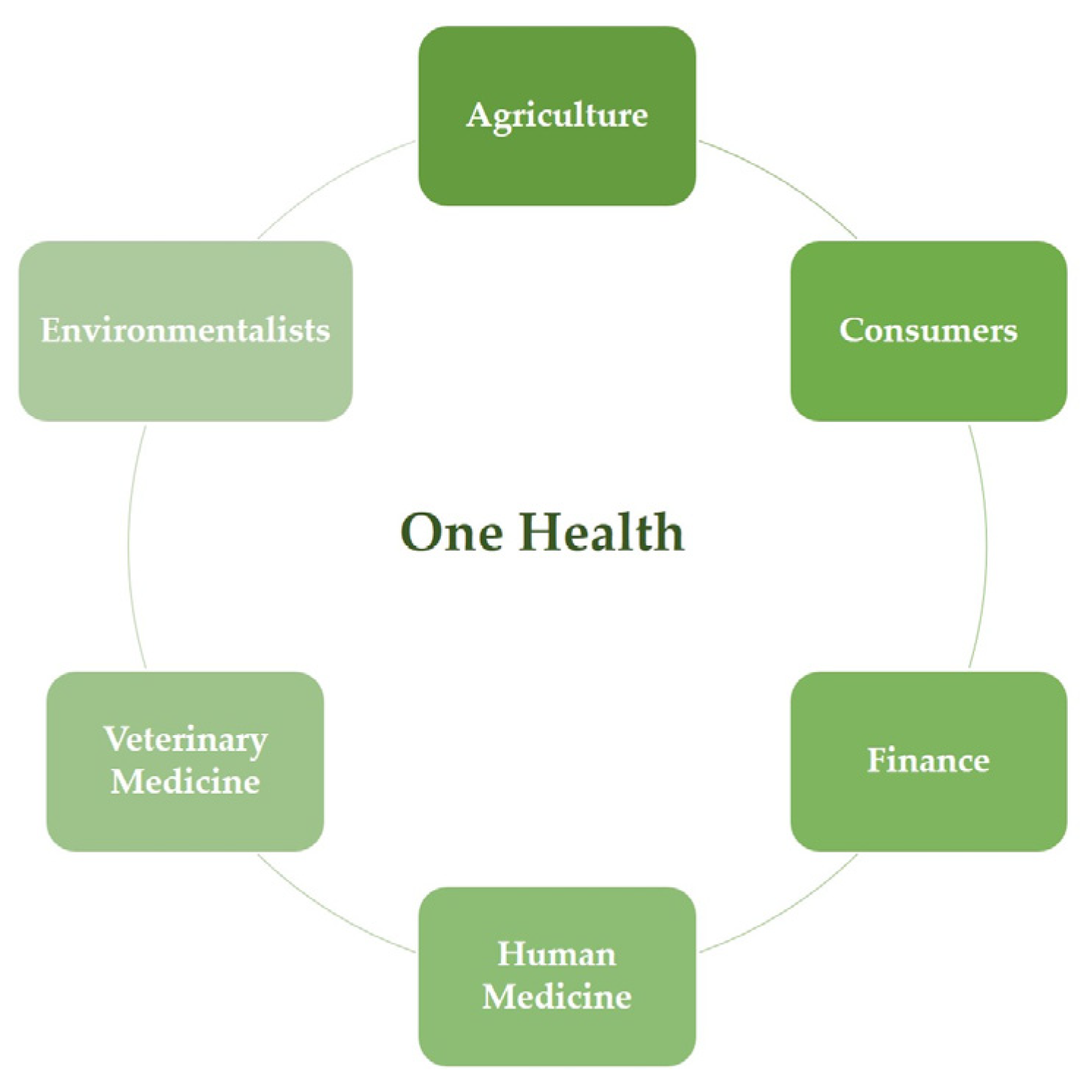
:1. Introduction
2. Antimicrobial Drug Resistance
2.1. Global Scenario
2.2. Antibiotics in Poultry Production
2.3. Development of Antimicrobial Drug Resistance
2.4. Transmission of Antimicrobial Drug Resistance
3. Important Pathogens in Poultry Production
3.1. Salmonella Enterica
3.2. Campylobacter spp.
3.3. Escherichia coli
3.4. Enterococcus spp.
3.5. Methicillin-Resistant Staphylococcus aureus
4. Strategies to Reduce Antimicrobial Drug Resistance in Poultry Production
4.1. Husbandry
4.2. Non-Conventional Antimicrobial Compounds
4.2.1. Antimicrobial Peptides
AMP Use as Growth Promoters
AMP Use as Immunomodulators
AMPs Used for Infection Therapy in Poultry
- Salmonella
- E. coli
- Campylobacter spp.
- C. perfringens

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| AMP | Origin | Target (Gram) | Trial | Dosage | Effect | Study |
|---|---|---|---|---|---|---|
| 3 bacteriocins | Paenibacillus polymyxa B. circulans | Campylobacter (−) | In vitro | na | Antimicrobial activity | [89] |
| Gallinacin-6 | Gallus gallus domesticus | C. jejuni (−), S. enterica (−), C. perfringens (+), E. coli (−) | In vitro | na | Antimicrobial activity | [93] |
| RSRP | Oryctolagus cuniculus—sacculus rotundus | E. coli (−) | In vivo | 0.1 mg of RSRP on d 7, 14, 21 and 28 | Immunomodulation; alteration of intestinal morphology | [83] |
| AS7 | Carnobacterium divergens AS7 | C. perfringens (+) | In vivo | 200 AU/g of feed for 42 days | Improvement of growth performance; alteration of intestinal microbiota | [94] |
| A-D-Asn | Pichia pastoris | - | In vivo | Basal diets with a A-D-Asn supplementation at 0, 2, 4, 6 and 8 mL/kg | Improvement of growth performance | [76] |
| BT | Brevibacillus texasporus | S. Enteritidis (−) | In vivo | 24 ppm BT peptide-supplemented diet | Immunomodulation in neonatal poultry; antimicrobial activity (prophylactic protection against Salmonella infections) | [81] |
| Nisin | Lactococcus lactis subsp. lactis | - | In vivo | Diet supplemented with various concentrations of nisin (100, 300, 900 and 2700 IU/g) | Improvement of growth performance; modulation of the GIT microbial ecology | [73] |
| CATH-2 | Chicken | S. Enteritidis (−) S. Typhimurium (−) | In vitro | na | Antimicrobial activity | [87] |
| A3 | Analog of Helicobacter pylori 2–20 | - | In vivo | Basal diet supplemented with 60 or 90 mg/kg AMP-A3 | Improvement of growth performance; alteration of intestinal microbiota | [77] |
| P5 | Analog of hybrid AMP CA-MA | - | In vivo | Basal diet supplemented with 40 and 60 mg/kg AMP-P5 | Improvement of growth performance; alteration of intestinal microbiota | [78] |
| Sublacin | B. subtilis 168 | C. perfringens (+) | In vivo | Chickens supplemented with sublancin at 2.88, 5.76 or 11.52 mg activity/L of water | Antimicrobial activity; alteration of intestinal morphology | [92] |
| Lactoferrin (bLf) | Bos taurus | E. coli (−) Salmonella (−) | In vivo | Diets with 130, 260 and 520 mg bLf/kg feed during the starter stage | Alteration of intestinal morphology | [82] |
| Nisin | Lactococcus lactis subsp. lactis | - | In vivo | 35-day administration of nisin at 2700 IU kg−1 through diet | Improvement of growth performance | [74] |
| Microcin J25 (Mcc) | Fecal strain of E. coli | E. coli AZ1 (−) Salmonella CVCC519(−) | In vivo | Basal diet with 0, 5 and 1 mg/kg Mcc J25 | Decrease in intestinal inflammation; alteration of intestinal microbiota; improvement of growth performance | [70] |
| Nisin | Lactococcus lactis subsp. lactis | - | In vivo | Administration of nisin in 2700 IU/kg via diet | Improvement of growth performance | [95] |
| cLFcuimera | Camel lactoferrin-derived peptide | E. coli (−), S. Enteritidis (−), S. aureus (+) | In vitro | na | Antimicrobial activity on both Gram-positive and Gram-negative avian pathogenic bacteria | [88] |
| cLF36 | Camel lactoferrin-derived peptide | E. coli (−) Clostridium spp. (+) | In vivo | 20 mg/kg AMP | Improvement of growth performance; immune modulator; alteration of intestinal microbiota | [45] |
| Plectasin (Ple) | Pseudoplectania nigrella | - | In vivo | Basal diet supplemented with 100 and 200 mg Ple/kg | Immunomodulation | [84] |
| Ctx(Ile21)-Ha | Hypsiboas albopunctatus | S. Enteriditis (−) | In vivo | 20 and 40 mg of Ctx(Ile21)-Ha/kg were included in the diet for 28 days | Antimicrobial activity | [64] |
| 11 AMPs | Chemically synthesized | C. jejuni (−) | In vitro | na | Antimicrobial activity | [61] |
4.2.2. Bacteriophages
Phage-Mediated Control at Pre-Harvest
- Salmonella
- Campylobacter spp.
- E. coli
- Staphylococcus aureus
| Phage | Origin | Target (Gram) | Administration Route | Dosage (PFU—Plaque-Forming Unit) | Effect | Study |
|---|---|---|---|---|---|---|
| Phage type 14, type 40 and type 141 | Raw human sewage | S. Typhimurium (−) | Feed | 105 and 1010 PFU/mL | Reduction in the viable numbers of S. Typhimurium in the crop, small intestine and caeca for up to 12 h after inoculation with smaller reductions in the liver at 24 and 48 h after infection | [108] |
| Phage R | Human sewage | E. coli (−) | Intramuscular Intracranial | 102–106 PFU 106–108 PFU | Long persistence of phages in tissues, which may be useful in prophylaxis and treatment of colibacillosis | [122] |
| DAF6 and SPR02 | Municipal and poultry processing waste | E. coli (−) | Aerosol Intramuscular | Aerosol spray: 7.65 × 108 (DAF6) and 2.83 × 109 (SPR02) PFU/mL; injection: 1.88 × 109 (DAF6) and 6.35 × 108 (SPR02) | Successful treatment of E. coli infection by intramuscular injection, which also could be used to prevent colibacillosis in poultry | [123] |
| Phage 69 (NCTC 12669) and Phage 71 (NCTC 12671) | National Collection of Type Cultures in the UK | C. jejuni (−) | Feed | 4 × 109 to 2 × 1010 PFU | Significant decrease in Campylobacter colonization | [116] |
| CP8 and CP34 | Ceca and upper and lower intestines of chicken | Campylobacter (−) | Feed | log10 5, 7 and 9 PFU | Decrease in the bacterial load depending on the amount of phage and time of administration | [117] |
| Esc-A | Sewage | E. coli (−) | Oral | 105 PFU | More efficient decrease in the death rate compared to chloramphenicol treatment | [125] |
| CP220 | nd | C. jejuni (−) C. coli (−) | Oral gavage | log10 5, 7 and 9 PFU | Decrease in Campylobacter colonization | [118] |
| phiCcoIBB35, phiCcoIBB37 and phiCcoIBB12 | Poultry intestinal contents | C. jejuni (−) C. coli (−) | Oral gavage/feed | Phage cocktail with 1 × 106–1.5 × 107 PFU | Reduction in the number of C. jejuni (Experiment 1) and C. coli (Experiment 2) colonization in chickens | [119] |
| UAB_Phi20, UAB_Phi78 and UAB_Phi87 | Chicken cloacae and pig rectal swabs | Salmonella spp. (−) | Oral | Phage cocktail with 1010 PFU/animal | The frequent treatment of the chickens with bacteriophages, especially prior to colonization of the intestinal tract by Salmonella, is required to achieve effective bacterial reduction over time | [109] |
| ST4, L13 and SG3 | Sewage water treatment | S. Gallinarum (−) | Feed | Phage cocktail with 108 PFU/kg | Significant decrease in bacterial isolation from the organs and mortality in chickens treated with the bacteriophages | [110] |
| Phages NCTC12672, 12673, 12674 and 12678 | British phage typing scheme | C. jejuni (−) | Drinking water | Phage cocktail with log10 5.8 to 7.5 PFU/bird | Decrease in Campylobacter load | [120] |
| Phages 1 (NCTC 12673), 2 (NCTC 12674), 5 (NCTC 12678) and 13 (NCTC 12672) | National Collection of Type Cultures | C. jejuni (−) | Oral | Single phage or a four-phage cocktail (107 PFU/bird) | Permanent reduction in Campylobacter load by the phage cocktail, as well as by the single phage. However, the cocktail delayed the emergence of phage resistance | [121] |
| nd | Sewage water taken at broiler farm | S. Typhimurium (−) S. Enteritidis (−) | Oral | 1.18 × 1011 PFU/chick to1.03 × 1012 PFU/chick | No detection of Salmonella in the cecum after the last (5th) dose | [111] |
4.2.3. Probiotics
| Probiotic | Microorganisms | Target (Gram) | Dosage (CFU—Colony-Forming Unit) | Effect | Study |
|---|---|---|---|---|---|
| FM-B11® | Lactobacillus spp. | - | 106 CFU/mL via drinking water | Significant reduction in mortality and increase in broiler chick performance | [132] |
| FM-B11® | Eleven lactic acid bacterial isolates | S. Enteritidis (−) | Oral doses of 104,106 and 108 CFU/bird | Possible reduction in Salmonella Enteritidis in neonatal chicks. | [133] |
| FM-B11® | Lactobacillus spp. | S. Enteritidis (−) | 4 × 106 CFU/mL via oral gavage | Significant reductions in the concentrations of S. Enteritidis within the ceca, and the timing of FM-B11® treatment affects S. Enteritidis-associated reductions | [135] |
| - | Bacillus spp. | S. Typhimurium (−) | Directly fed microbials at 106 spores/g of feed | Significantly lower cecal S. Typhimurium load and increased performance | [134] |
| Gallipro® | B. subtilis | S. Enteritidis (−) | 0.02% probiotic of diet supplementation | No significant effect at non-contaminated environment, showing a greater efficacy at a pathogen contaminated environment and improving immune response of infected chickens | [136] |
| - | E. faecium PXN33 Ligilactobacillus salivarius (ex Lactobacillus salivarius [131]) 59 | S. Enteritidis (−) | Oral gavage with 1 × 109 CFU of probiotics | Prevention of S. Enteritidis colonization of poultry | [137] |
| - | Bacillus spp. | S. Typhimurium (−) | na | Reinstatement of the microbial genera displaced by S. Typhimurium challenge | [138] |
| - | Bacillus spp. | S. Enteritidis (−) | Feed supplemented in concentration of 454 g/ton | Reduction in the load of Salmonella in the ceca. | [139] |
| - | B. subtilis CSL2 | Salmonella (−) | Feed supplemented in concentration of 1.0 × 107 CFU/g of feed | Modulation in the microbiota, potentially protecting against S. Gallinarum infection | [140] |
| - | L. salivarius | S. Pullorum (−) | Feed supplemented in concentrations of 107, 108 and 109 CFU/kg of feed | Enhancement in S. Pullorum infection resistance in broilers challenged with Aflatoxin B1 | [141] |
| PrimaLac (Star Labs, St. Joseph, MO, USA) | Lactobacillus acidophilus, Lactobacillus casei, Bifidobacterium thermophilus and Enterococcus faecium | C. jejuni (−) | Minimum of 1.04 × 108 CFU/g | Reduction in the presence of C. jejuni, but no significant effect on the growth performance of broilers | [142] |
| - | Lactobacillus plantarum PCS 20 and Bifidobacterium longum PCB 133 | C. jejuni (−) | Oral gavages in concentration of 108 CFU for 15 days | B. longum PCB 133 led to significant reduction in C. jejuni concentration in poultry feces. | [143] |
| - | Lactobacillus gasseri SBT2055 | C. jejuni (−) | Oral gavages in concentration of 1 × 108 CFU CFU for 14 days | Significant reduction in cecum colonization by C. jejuni at 14 days after infection. | [144] |
| - | Lactobacillus gasseri SBT2055 | C. jejuni (−) | na | Significant reduction in cecum colonization by C. jejuni. | [145] |
| - | Bacillus spp. L. salivarus subsp. salivarius L. salivarus subsp. salicinus | Campylobacter(−) | na | These strains had significantly reduced C. jejuni counts at 14 days after infection. | [146] |
4.2.4. Nanoparticles
| NP | Target (Gram) | Origin | Trial | Dosage | Effect | Study |
|---|---|---|---|---|---|---|
| ZnO | Salmonella spp. (−), E. coli (−), S. aureus (+) | Lactiplantibacillus plantarum (ex Lactobacillus plantarum [131] TA4 | In vitro | na | Effective antibacterial activity, particularly against S. aureus | [149] |
| Ag | C. jejuni (−) | High purity metals and high purity demineralized water | In vivo | 50 ppm of AgNPs in the drinking water for 30 days | Reduced growth, impaired immune functions and no antibacterial effect on different intestinal bacterial groups | [168] |
| C. perfringens (+) | Chemical reduction of silver nitrate | In vivo | 150 µg of AgNPs via crop gavage for 5 days post infection | Reduced colonization of C. perfringens in the intestine, positive impact on gut health integrity, and no impact on immune system | [162] | |
| Fe3O4 | S. Enteritidis (−) | FeCl3 and NaAc.3H2O | In vivo | Oral 50 mg/kg Fe3O4-NPs | Pretreatment could significantly decrease the invasion of S. Enteritidis in chickens and attenuate morphological changes caused by the infection | [69] |
| Chitosan | E. coli (−) | nd | In vitro | na | Antibacterial activity against biofilm-forming strains | [164] |
| Chitosan encapsulation | - | Crab shell and sodium triphosphate pentabasic | In vivo | Non-encapsulated (2) and nanoencapsulated extracts of Aloe vera, dill and nettle roots (3) in 0.02, 0.025 and 0.05% to starter, grower and finisher diets for 42 days | Nanoencapsulation of dill extract could improve growth performance and can be used as a substitute for AGPs in the diet | [152] |
| - | nd | In vivo | Diet with 2.5% of Phaleria macrocarpa fruit extract (2) with nanoencapsulated Phaleria, macrocarpa fruits extract (NEPM) 2.5% NEPM (3) and with 5.0% NEPM (4) | Diet supplementation with 5.0% of NEMP had positive impact on modulation in the intestinal microbiota and limitation in the growth of pathogenic bacteria | [166] |
5. Challenges of the Application of These Innovative Compounds
5.1. Antimicrobial Peptides
5.2. Bacteriophages
5.3. Nanoparticles
6. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- United Nations Department of Economic and Social Affairs. World Population Prospects 2022—Summary of Results. 2022. Available online: https://www.un.org/development/desa/pd/sites/www.un.org.development.desa.pd/files/wpp2022_summary_of_results.pdf (accessed on 15 November 2022).
- Richie, H.; Rosado, P.; Roser, M. Meat and Dairy Production. Available online: https://ourworldindata.org/meat-production (accessed on 16 November 2022).
- EuroStat. Agricultural Production—Livestock and Meat. Available online: https://ec.europa.eu/eurostat/statistics-explained/index.php?title=Agricultural_production_-_livestock_and_meat#Poultry (accessed on 15 December 2022).
- FAO. Gateway to Poultry Production and Products. Available online: https://www.fao.org/poultry-production-products/production/en/ (accessed on 15 March 2023).
- Nazeer, N.; Uribe-Diaz, S.; Rodriguez-Lecompte, J.C.; Ahmed, M. Antimicrobial Peptides as an Alternative to Relieve Antimicrobial Growth Promoters in Poultry. Br. Poult. Sci. 2021, 62, 672–685. [Google Scholar] [CrossRef] [PubMed]
- Espinosa-Marrón, A.; Adams, K.; Sinno, L.; Cantu-Aldana, A.; Tamez, M.; Marrero, A.; Bhupathiraju, S.N.; Mattei, J. Environmental Impact of Animal-Based Food Production and the Feasibility of a Shift toward Sustainable Plant-Based Diets in the United States. Front. Sustain. 2022, 3, 19. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention. One Health Basics. Available online: https://www.cdc.gov/onehealth/basics/index.html (accessed on 30 November 2022).
- Mohr, K.I. History of Antibiotics. Curr. Top. Microbiol. Immunol. 2016, 398, 237–272. [Google Scholar] [CrossRef] [PubMed]
- Founou, L.L.; Founou, R.C.; Essack, S.Y. Antibiotic Resistance in the Food Chain: A Developing Country-Perspective. Front. Microbiol. 2016, 7, 1881. [Google Scholar] [CrossRef] [PubMed]
- Verraes, C.; Van Boxstael, S.; Van Meervenne, E.; Van Coillie, E.; Butaye, P.; Catry, B.; de Schaetzen, M.A.; Van Huffel, X.; Imberechts, H.; Dierick, K.; et al. Antimicrobial Resistance in the Food Chain: A Review. Int. J. Environ. Res. Public Health 2013, 10, 2643–2669. [Google Scholar] [CrossRef] [Green Version]
- EIP AGRI Focus Group. Reducing Antimicrobial Use in Poultry Farming. 2021. Available online: https://ec.europa.eu/eip/agriculture/sites/default/files/eip-agri_fg_reducing_antimicrobial_use_in_poultry_farming_final_report_2021_en.pdf (accessed on 2 November 2022).
- Davies, J.; Davies, D. Origins and Evolution of Antibiotic Resistance. Microbiol. Mol. Biol. Rev. 2010, 74, 417–433. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization. Global Action Plan on Antimicrobial Resistance; World Health Organization: Geneva, Switzerland, 2015; pp. 1–28.
- EMA. Categorisation of Antibiotics Used in Animals Promotes Responsible Use to Protect Public and Animal Health. Available online: https://www.ema.europa.eu/en/news/categorisation-antibiotics-used-animals-promotes-responsible-use-protect-public-animal-health (accessed on 15 January 2023).
- Al Sattar, A.; Chisty, N.N.; Irin, N.; Uddin, M.H.; Hasib, F.M.Y.; Hoque, M.A. Knowledge and Practice of Antimicrobial Usage and Resistance among Poultry Farmers: A Systematic Review, Meta-Analysis, and Meta-Regression. Vet. Res. Commun. 2023. [Google Scholar] [CrossRef]
- Joerger, R.D. Alternatives to Antibiotics: Bacteriocins, Antimicrobial Peptides and Bacteriophages. Poult. Sci. 2003, 82, 640–647. [Google Scholar] [CrossRef]
- Rodrigues, G.; Santos, L.S.; Franco, O.L. Antimicrobial Peptides Controlling Resistant Bacteria in Animal Production. Front. Microbiol. 2022, 13, 874153. [Google Scholar] [CrossRef]
- Castanon, J.I.R. History of the Use of Antibiotic as Growth Promoters in European Poultry Feeds. Poult. Sci. 2007, 86, 2466–2471. [Google Scholar] [CrossRef]
- Hudson, J.A.; Frewer, L.J.; Jones, G.; Brereton, P.A.; Whittingham, M.J.; Stewart, G. The Agri-Food Chain and Antimicrobial Resistance: A Review. Trends Food Sci. Technol. 2017, 69, 131–147. [Google Scholar] [CrossRef] [Green Version]
- Rushton, J. Anti-Microbial Use in Animals: How to Assess the Trade-Offs. Zoonoses Public Health 2015, 62, 10–21. [Google Scholar] [CrossRef] [Green Version]
- Luiken, R.E.C.; Van Gompel, L.; Munk, P.; Sarrazin, S.; Joosten, P.; Dorado-García, A.; Borup Hansen, R.; Knudsen, B.E.; Bossers, A.; Wagenaar, J.A.; et al. Associations between Antimicrobial Use and the Faecal Resistome on Broiler Farms from Nine European Countries. J. Antimicrob. Chemother. 2019, 74, 2596–2604. [Google Scholar] [CrossRef] [Green Version]
- Krysiak, K.; Konkol, D.; Korczyński, M. Overview of the Use of Probiotics in Poultry Production. Animals 2021, 11, 1620. [Google Scholar] [CrossRef] [PubMed]
- Holmes, A.H.; Moore, L.S.P.; Sundsfjord, A.; Steinbakk, M.; Regmi, S.; Karkey, A.; Guerin, P.J.; Piddock, L.J.V. Understanding the Mechanisms and Drivers of Antimicrobial Resistance. Lancet 2016, 387, 176–187. [Google Scholar] [CrossRef] [PubMed]
- Bester, L.A.; Essack, S.Y. Antibiotic Resistance via the Food Chain: Fact or Fiction? S. Afr. J. Sci. 2010, 106, 1–5. [Google Scholar] [CrossRef]
- Acar, J.F.; Moulin, G. Antimicrobial Resistance at Farm Level. Rev. Sci. Tech. 2006, 25, 775–792. [Google Scholar] [CrossRef]
- Marshall, B.M.; Levy, S.B. Food Animals and Antimicrobials: Impacts on Human Health. Clin. Microbiol. Rev. 2011, 24, 718–733. [Google Scholar] [CrossRef] [Green Version]
- Da Costa, P.M.; Loureiro, L.; Matos, A.J.F. Transfer of Multidrug-Resistant Bacteria between Intermingled Ecological Niches: The Interface between Humans, Animals and the Environment. Int. J. Environ. Res. Public Health 2013, 10, 278–294. [Google Scholar] [CrossRef] [Green Version]
- Djordjevic, S.P.; Stokes, H.W.; Chowdhury, P.R. Mobile Elements, Zoonotic Pathogens and Commensal Bacteria: Conduits for the Delivery of Resistance Genes into Humans, Production Animals and Soil Microbiota. Front. Microbiol. 2013, 4, 86. [Google Scholar] [CrossRef] [Green Version]
- Żbikowska, K.; Michalczuk, M.; Dolka, B. The Use of Bacteriophages in the Poultry Industry. Animals 2020, 10, 872. [Google Scholar] [CrossRef]
- de Saraiva, M.M.S.; Lim, K.; do Monte, D.F.M.; Givisiez, P.E.N.; Alves, L.B.R.; de Neto, O.C.F.; Kariuki, S.; Júnior, A.B.; de Oliveira, C.J.B.; Gebreyes, W.A. Antimicrobial Resistance in the Globalized Food Chain: A One Health Perspective Applied to the Poultry Industry. Braz. J. Microbiol. 2022, 53, 465–486. [Google Scholar] [CrossRef] [PubMed]
- European, T.; One, U.; Report, Z. The European Union One Health 2020 Zoonoses Report. EFSA J. 2021, 19, e06971. [Google Scholar] [CrossRef]
- Lorenzo-Rebenaque, L.; Malik, D.J.; Catalá-Gregori, P.; Torres, J.; Marin, C.; Sevilla-Navarro, S. Microencapsulated Bacteriophages Incorporated in Feed for Salmonella Control in Broilers. Vet. Microbiol. 2022, 274, 109579. [Google Scholar] [CrossRef]
- Ricke, S.C. Strategies to Improve Poultry Food Safety, a Landscape Review. Annu. Rev. Anim. Biosci. 2021, 9, 379–400. [Google Scholar] [CrossRef] [PubMed]
- Chai, S.J.; Cole, D.; Nisler, A.; Mahon, B.E. Poultry: The Most Common Food in Outbreaks with Known Pathogens, United States, 1998–2012. Epidemiol. Infect. 2017, 145, 316–325. [Google Scholar] [CrossRef] [Green Version]
- Cosby, D.E.; Cox, N.A.; Harrison, M.A.; Wilson, J.L.; Jeff Buhr, R.; Fedorka-Cray, P.J. Salmonella and Antimicrobial Resistance in Broilers: A Review. J. Appl. Poult. Res. 2015, 24, 408–426. [Google Scholar] [CrossRef]
- Issenhuth-Jeanjean, S.; Roggentin, P.; Mikoleit, M.; Guibourdenche, M.; De Pinna, E.; Nair, S.; Fields, P.I.; Weill, F.X. Supplement 2008–2010 (No. 48) to the White-Kauffmann-Le Minor Scheme. Res. Microbiol. 2014, 165, 526–530. [Google Scholar] [CrossRef] [Green Version]
- Antunes, P.; Mourão, J.; Campos, J.; Peixe, L. Salmonellosis: The Role of Poultry Meat. Clin. Microbiol. Infect. 2016, 22, 110–121. [Google Scholar] [CrossRef] [Green Version]
- Alali, W.Q.; Hofacre, C.L. Preharvest Food Safety in Broiler Chicken Production. In Preharvest Food Safety; Thakur, S., Kniel, K.E., Eds.; ASM Press: Washington, DC, USA, 2018; pp. 69–86. [Google Scholar] [CrossRef]
- Veterinary Medicines Directorate. UK One Health Report—Joint Report on Antibiotic Use and Antibiotic Resistance, 2013–2017; Veterinary Medicines Directorate: Addlestone, UK, 2019. Available online: https://www.gov.uk/government/uploads/system/uploads/attachment_data/file/447319/One_Health_Report_July2015.pdf (accessed on 20 December 2022).
- Silva, J.; Leite, D.; Fernandes, M.; Mena, C.; Gibbs, P.A.; Teixeira, P. Campylobacter spp. As a Foodborne Pathogen: A Review. Front. Microbiol. 2011, 2, 200. [Google Scholar] [CrossRef] [Green Version]
- Sahin, O.; Kassem, I.I.; Shen, Z.; Lin, J.; Rajashekara, G.; Sahin, O.; Kassem, A.I.I.; Shen, B.Z.; Lin, A.J.; Rajashekara, C.G.; et al. Campylobacter in Poultry: Ecology and Potential Interventions. Avian Dis. 2015, 59, 185–200. [Google Scholar] [CrossRef]
- Van Deun, K.; Pasmans, F.; Ducatelle, R.; Flahou, B.; Vissenberg, K.; Martel, A.; Van den Broeck, W.; Van Immerseel, F.; Haesebrouck, F. Colonization Strategy of Campylobacter jejuni Results in Persistent Infection of the Chicken Gut. Vet. Microbiol. 2008, 130, 285–297. [Google Scholar] [CrossRef]
- Hermans, D.; Pasmans, F.; Messens, W.; Martel, A.; Van Immerseel, F.; Rasschaert, G.; Heyndrickx, M.; Van Deun, K.; Haesebrouck, F. Poultry as a Host for the Zoonotic Pathogen Campylobacter jejuni. Vector-Borne Zoonotic Dis. 2012, 12, 89–98. [Google Scholar] [CrossRef] [Green Version]
- Public Health England. Summary of Antimicrobial Prescribing Guidance: Managing Common Infections. 2018. Available online: https://assets.publishing.service.gov.uk/government/uploads/system/uploads/attachment_data/file/994444/Common_Infect_PHE_context_references_and_rationale_May_2021_Bites_and_Eczema__1_.pdf (accessed on 20 January 2023).
- Daneshmand, A.; Kermanshahi, H.; Sekhavati, M.H.; Javadmanesh, A.; Ahmadian, M. Antimicrobial Peptide, CLF36, Affects Performance and Intestinal Morphology, Microflora, Junctional Proteins, and Immune Cells in Broilers Challenged with E. coli. Sci. Rep. 2019, 9, 14176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Islam, M.S.; Nayeem, M.M.H.; Sobur, M.A.; Ievy, S.; Islam, M.A.; Rahman, S.; Kafi, M.A.; Ashour, H.M.; Rahman, M.T. Virulence Determinants and Multidrug Resistance of Escherichia coli Isolated from Migratory Birds. Antibiotics 2021, 10, 190. [Google Scholar] [CrossRef] [PubMed]
- Bass, L.; Liebert, C.A.; Lee, M.D.; Summers, A.O.; White, D.G.; Thayer, S.G.; Maurer, J.J. Incidence and Characterization of Integrons, Genetic Elements Mediating Multiple-Drug Resistance, in Avian Escherichia coli. Antimicrob. Agents Chemother. 1999, 43, 2925–2929. [Google Scholar] [CrossRef] [Green Version]
- Nolan, L.K.; Barnes, H.J.; Vaillancourt, J.; Abdul-aziz, T.; Logue, C.M. Colibacillosis. In Diseases of Poultry; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2013; pp. 751–805. [Google Scholar]
- Mitchell, N.M.; Johnson, J.R.; Johnston, B.; Curtiss, R.; Mellata, M. Zoonotic Potential of Escherichia coli Isolates from Retail Chicken Meat Products and Eggs. Appl. Environ. Microbiol. 2015, 81, 1177–1187. [Google Scholar] [CrossRef] [Green Version]
- Dea, M.O.; Sahibzada, S.; Jordan, D.; Laird, T.; Lee, T.; Hewson, K.; Pang, S.; Abraham, R.; Coombs, G.W.; Harris, T.; et al. Genomic, Antimicrobial Resistance, and Public Health Insights into Enterococcus spp. from Australian Chickens. J. Clin. Microbiol. 2019, 57, e00319. [Google Scholar]
- Kabir, S.M.L. The Role of Probiotics in the Poultry Industry. Int. J. Mol. Sci. 2009, 10, 3531–3546. [Google Scholar] [CrossRef] [PubMed]
- Souillard, R.; Laurentie, J.; Kempf, I.; Le Caër, V.; Le Bouquin, S.; Serror, P.; Allain, V. Increasing Incidence of Enterococcus-Associated Diseases in Poultry in France over the Past 15 Years. Vet. Microbiol. 2022, 269, 109426. [Google Scholar] [CrossRef]
- Devriese, L.A.; Cawerts, K.; Hermans, K.; Wood, A.M. Enterococcus cecorum Septicaemia as a Cause of Bone and Joint Lesions Resulting in Lameness in Broiler Chickens. Vlaams Diergeneeskd. Tijdschr. 2002, 71, 219–221. [Google Scholar]
- Khan, H.A.; Ahmad, A.; Mehboob, R. Nosocomial Infections and Their Control Strategies. Asian Pac. J. Trop. Biomed. 2015, 5, 509–514. [Google Scholar] [CrossRef] [Green Version]
- Lee, G.Y.; Lee, S.I.; Do Kim, S.; Park, J.H.; Kim, G.-B.; Yang, S.-J. Clonal Distribution and Antimicrobial Resistance of Methicillin-Susceptible and -Resistant Staphylococcus aureus Strains Isolated from Broiler Farms, Slaughterhouses, and Retail Chicken Meat. Poult. Sci. 2020, 101, 102070. [Google Scholar] [CrossRef] [PubMed]
- Feßler, A.T.; Kadlec, K.; Hassel, M.; Hauschild, T.; Eidam, C.; Ehricht, R.; Monecke, S.; Schwarz, S. Characterization of Methicillin-Resistant Staphylococcus aureus Isolates from Food and Food Products of Poultry Origin in Germany. Appl. Environ. Microbiol. 2011, 77, 7151–7157. [Google Scholar] [CrossRef] [Green Version]
- Verkade, E.; Kluytmans, J. Livestock-Associated Staphylococcus aureus CC398: Animal Reservoirs and Human Infections. Infect. Genet. Evol. 2014, 21, 523–530. [Google Scholar] [CrossRef]
- Chakraborty, N.; Jha, D.; Roy, I.; Kumar, P.; Gaurav, S.S.; Marimuthu, K.; Ng, O.T.; Lakshminarayanan, R.; Verma, N.K.; Gautam, H.K. Nanobiotics against Antimicrobial Resistance: Harnessing the Power of Nanoscale Materials and Technologies. J. Nanobiotechnol. 2022, 20, 375. [Google Scholar] [CrossRef]
- Sommer, J.; Trautner, C.; Witte, A.K.; Fister, S.; Schoder, D.; Rossmanith, P.; Mester, P.-J. Don’t Shut the Stable Door after the Phage Has Bolted—The Importance of Bacteriophage Inactivation in Food Environments. Viruses 2019, 11, 468. [Google Scholar] [CrossRef] [Green Version]
- Rawson, T.; Dawkins, M.S.; Bonsall, M.B. A Mathematical Model of Campylobacter Dynamics within a Broiler Flock. Front. Microbiol. 2019, 10, 1940. [Google Scholar] [CrossRef] [Green Version]
- Line, J.E.; Seal, B.S.; Garrish, J.K. Selected Antimicrobial Peptides Inhibit in Vitro Growth of Campylobacter spp. Appl. Microbiol. 2022, 2, 688–700. [Google Scholar] [CrossRef]
- Cruz, G.S.; dos Santos, A.T.; de Brito, E.H.S.; Rádis-Baptista, G. Cell-Penetrating Antimicrobial Peptides with Anti-Infective Activity against Intracellular Pathogens. Antibiotics 2022, 11, 1772. [Google Scholar] [CrossRef]
- Lai, Y.; Gallo, R.L. AMPed Up Immunity: How Antimicrobial Peptides Have Multiple Roles in Immune Defense. Trends Imunnology 2009, 30, 131–141. [Google Scholar] [CrossRef] [Green Version]
- Roque-Borda, C.A.; Pereira, L.P.; Guastalli, E.A.L.; Soares, N.M.; Mac-Lean, P.A.B.; Salgado, D.D.; Meneguin, A.B.; Chorilli, M.; Vicente, E.F. HPMCP-Coated Microcapsules Containing the Ctx(Ile21)-Ha Antimicrobial Peptide Reduce the Mortality Rate Caused by Resistant Salmonella enteritidis in Laying Hens. Antibiotics 2021, 10, 616. [Google Scholar] [CrossRef] [PubMed]
- Magana, M.; Pushpanathan, M.; Santos, A.L.; Leanse, L.; Fernandez, M.; Ioannidis, A.; Giulianotti, M.A.; Apidianakis, Y.; Bradfute, S.; Ferguson, A.L.; et al. The Value of Antimicrobial Peptides in the Age of Resistance. Lancet Infect. Dis. 2020, 20, e216–e230. [Google Scholar] [CrossRef] [PubMed]
- Cunha, E.; Tavares, L.; Veiga, A.S.; Oliveira, M. Fighting Antimicrobial Resistance: The Biomedical Use of the Antimicrobial Peptide Nisin. In Advances in Medicine and Biology; Nova Science Publishers: Hauppauge, NY, USA, 2019; pp. 79–112. [Google Scholar]
- Yadav, S.; Jha, R. Strategies to Modulate the Intestinal Microbiota and Their Effects on Nutrient Utilization, Performance, and Health of Poultry. J. Anim. Sci. Biotechnol. 2019, 10, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Apajalahti, J.; Vienola, K. Interaction between Chicken Intestinal Microbiota and Protein Digestion. Anim. Feed Sci. Technol. 2016, 221, 323–330. [Google Scholar] [CrossRef] [Green Version]
- Shen, Y.; Xiao, Y.; Zhang, S.; Wu, S.; Gao, L.; Shi, S. Fe3O4 Nanoparticles Attenuated Salmonella Infection in Chicken Liver Through Reactive Oxygen and Autophagy via PI3K/Akt/MTOR Signaling. Front. Physiol. 2020, 10, 1580. [Google Scholar] [CrossRef] [Green Version]
- Wang, G. Effect of Antimicrobial Peptide Microcin J25 on Growth Performance, Immune Regulation, and Intestinal Microbiota in Broiler Chickens Challenged with Escherichia coli and Salmonella. Animals 2020, 10, 345. [Google Scholar] [CrossRef] [Green Version]
- Iseppi, R.; Lauková, A.; Sabia, C. Bacteriocin-Producing Probiotic Bacteria: A Natural Solution for Increasing Efficiency and Safety of Livestock Food Production. Front. Microbiol. 2021, 12, 675483. [Google Scholar] [CrossRef] [PubMed]
- Kierończyk, B.; Rawski, M.; Mikołajczak, Z.; Świątkiewicz, S.; Józefiak, D. Nisin as a Novel Feed Additive: The Efffects on Gut Microbial Modulation and Activity, Histological Parameters, and Growth Performance of Broiler Chickens. Animals 2020, 10, 101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Józefiak, D.; Kierończyk, B.; Juśkiewicz, J.; Zduńczyk, Z.; Rawski, M.; Długosz, J.; Sip, A.; Højberg, O. Dietary Nisin Modulates the Gastrointestinal Microbial Ecology and Enhances Growth Performance of the Broiler Chickens. PLoS ONE 2013, 8, e85347. [Google Scholar] [CrossRef] [Green Version]
- Kierończyk, B.; Pruszyńska-Oszmałek, E.; Światkiewicz, S.; Rawski, M.; Długosz, J.; Engberg, R.M.; Józefiak, D. The Nisin Improves Broiler Chicken Growth Performance and Interacts with Salinomycin in Terms of Gastrointestinal Tract Microbiota Composition. J. Anim. Feed Sci. 2016, 25, 309–316. [Google Scholar] [CrossRef]
- Jozefiak, D.; Sip, A.; Rawski, M.; Rutkowski, A.; Kaczmarek, S.; Hojberg, O.; Jensen, B.B.; Engberg, R.M. Dietary Divercin Modifies Gastrointestinal Microbiota and Improves Growth Performance in Broiler Chickens. Br. Poult. Sci. 2011, 52, 492–499. [Google Scholar] [CrossRef] [PubMed]
- Wen, L.F.; He, J.G. Dose-Response Effects of an Antimicrobial Peptide, a Cecropin Hybrid, on Growth Performance, Nutrient Utilisation, Bacterial Counts in the Digesta and Intestinal Morphology in Broilers. Br. J. Nutr. 2012, 108, 1756–1763. [Google Scholar] [CrossRef] [Green Version]
- Choi, S.C.; Ingale, S.L.; Kim, J.S.; Park, Y.K.; Kwon, I.K.; Chae, B.J. An Antimicrobial Peptide-A3: Effects on Growth Performance, Nutrient Retention, Intestinal and Faecal Microflora and Intestinal Morphology of Broilers. Br. Poult. Sci. 2013, 54, 738–746. [Google Scholar] [CrossRef]
- Choi, S.C.; Ingale, S.L.; Kim, J.S.; Park, Y.K.; Kwon, I.K.; Chae, B.J. Effects of Dietary Supplementation with an Antimicrobial Peptide-P5 on Growth Performance, Nutrient Retention, Excreta and Intestinal Microflora and Intestinal Morphology of Broilers. Anim. Feed Sci. Technol. 2013, 185, 78–84. [Google Scholar] [CrossRef]
- Sochacki, K.A.; Barns, K.J.; Bucki, R.; Weisshaar, J.C. Real-Time Attack on Single Escherichia coli Cells by the Human Antimicrobial Peptide LL-37. Proc. Natl. Acad. Sci. USA 2011, 108, E77–E81. [Google Scholar] [CrossRef] [Green Version]
- Peng, L.; Scheenstra, M.R.; van Harten, R.M.; Haagsman, H.P.; Veldhuizen, E.J.A. The Immunomodulatory Effect of Cathelicidin-B1 on Chicken Macrophages. Vet. Res. 2020, 51, 122. [Google Scholar] [CrossRef]
- Kogut, M.H.; Genovese, K.J.; He, H.; Swaggerty, C.L.; Jiang, Y. Modulation of Chicken Intestinal Immune Gene Expression by Small Cationic Peptides as Feed Additives during the First Week Posthatch. Clin. Vaccine Immunol. 2013, 20, 1440–1448. [Google Scholar] [CrossRef] [Green Version]
- Aguirre, A.A.T.; Acda, S.P.; Angeles, A.A.; Cynthia Oliveros, M.R.; Merca, F.E.; Cruz, F.A. Effect of Bovine Lactoferrin on Growth Performance and Intestinal Histologic Features of Broilers. Philipp. J. Vet. Anim. Sci. 2015, 41, 11–20. [Google Scholar]
- Liu, T.; She, R.; Wang, K.; Bao, H.; Zhang, Y.; Luo, D.; Hu, Y.; Ding, Y.; Wang, D.; Peng, K. Effects of Rabbit Sacculus Rotundus Antimicrobial Peptides on the Intestinal Mucosal Immunity in Chickens. Poult. Sci. 2008, 87, 250–254. [Google Scholar] [CrossRef]
- Ma, J.L.; Zhao, L.H.; Sun, D.D.; Zhang, J.; Guo, Y.P.; Zhang, Z.Q.; Ma, Q.G.; Ji, C.; Zhao, L.H. Effects of Dietary Supplementation of Recombinant Plectasin on Growth Performance, Intestinal Health and Innate Immunity Response in Broilers. Probiotics Antimicrob. Proteins 2020, 12, 214–223. [Google Scholar] [CrossRef] [PubMed]
- Hernández-González, J.C.; Martínez-Tapia, A.; Lazcano-Hernández, G.; García-Pérez, B.E.; Castrejón-Jiménez, N.S. Bacteriocins from Lactic Acid Bacteria. A Powerful Alternative as Antimicrobials, Probiotics, and Immunomodulators in Veterinary Medicine. Animals 2021, 11, 979. [Google Scholar] [CrossRef] [PubMed]
- Turner, J.; Cho, Y.; Dinh, N.N.; Waring, A.J.; Lehrer, R.I. Activities of LL-37, a Cathelin-Associated Antimicrobial Peptide of Human Neutrophils. Antimicrob. Agents Chemother. 1998, 42, 2206–2214. [Google Scholar] [CrossRef] [Green Version]
- Veldhuizen, E.J.A.; Brouwer, E.C.; Schneider, V.A.F.; Fluit, A.C. Chicken Cathelicidins Display Antimicrobial Activity against Multiresistant Bacteria without Inducing Strong Resistance. PLoS ONE 2013, 8, e61964. [Google Scholar] [CrossRef] [Green Version]
- Tanhaiean, A.; Azghandi, M.; Razmyar, J.; Mohammadi, E.; Sekhavati, M.H. Recombinant Production of a Chimeric Antimicrobial Peptide in E. coli and Assessment of Its Activity against Some Avian Clinically Isolated Pathogens. Microb. Pathog. 2018, 122, 73–78. [Google Scholar] [CrossRef] [PubMed]
- Svetoch, E.A.; Stern, N.J.; Eruslanov, B.V.; Kovalev, Y.N.; Volodina, L.I.; Perelygin, V.V.; Mitsevich, E.V.; Mitsevich, I.P.; Pokhilenko, V.D.; Borzenkov, V.N.; et al. Isolation of Bacillus circulans and Paenibacillus polymyxa Strains Inhibitory to Campylobacter jejuni and Characterization of Associated Bacteriocins. J. Food Prot. 2005, 68, 11–17. [Google Scholar] [CrossRef] [Green Version]
- Daeschel, M.A.; Klaenhammer, T.R. Association of a 13.6-Megadalton Plasmid in Pedicococcus pentosaceus with Bacteriocin Activity. Appl. Environ. Microbiol. 1985, 50, 1538–1541. [Google Scholar] [CrossRef] [Green Version]
- Grilli, E.; Messina, M.; Catelli, E.; Morlacchini, M.; Piva, A. Pediocin a Improves Growth Performance of Broilers Challenged with Clostridium perfringens. Poult. Sci. 2009, 88, 2152–2158. [Google Scholar] [CrossRef]
- Wang, S.; Zeng, X.F.; Wang, Q.W.; Zhu, J.L.; Peng, Q.; Hou, C.; Thacker, P.; Qiao, S.Y. The Antimicrobial Peptide Sublancin Ameliorates Necrotic Enteritis Induced by Clostridium perfringens in Broilers. Am. Soc. Anim. Sci. 2015, 93, 4750–4760. [Google Scholar] [CrossRef]
- Van Dijk, A.; Veldhuizen, E.J.A.; Kalkhove, S.I.C.; Tjeerdsma-Van Bokhoven, J.L.M.; Romijn, R.A.; Haagsman, H.P. The β-Defensin Gallinacin-6 Is Expressed in the Chicken Digestive Tract and Has Antimicrobial Activity against Food-Borne Pathogens. Antimicrob. Agents Chemother. 2007, 51, 912–922. [Google Scholar] [CrossRef] [Green Version]
- Józefiak, D.; Sip, A.; Rutkowski, A.; Rawski, M.; Kaczmarek, S.; Wołuń-Cholewa, M.; Engberg, R.M.; Højberg, O. Lyophilized Carnobacterium divergens AS7 Bacteriocin Preparation Improves Performance of Broiler Chickens Challenged with Clostridium perfringens. Poult. Sci. 2012, 91, 1899–1907. [Google Scholar] [CrossRef]
- Kieronczyk, B.; Sassek, M.; Pruszynska-Oszmalek, E.; Kolodziejski, P.; Rawski, M.; Swiatkiewicz, S.; Józefiak, D. The Physiological Response of Broiler Chickens to the Dietary Supplementation of the Bacteriocin Nisin and Ionophore Coccidiostats. Poult. Sci. 2017, 96, 4026–4037. [Google Scholar] [CrossRef]
- Duckworth, D.H. Who Discovered Bacteriophage? Bacteriol. Rev. 1976, 40, 793–802. [Google Scholar] [CrossRef]
- Manrique, P.; Bolduc, B.; Walk, S.T.; Van Oost, J.D.; De Vos, W.M.; Young, M.J. Healthy Human Gut Phageome. Proc. Natl. Acad. Sci. USA 2016, 113, 10400–10405. [Google Scholar] [CrossRef] [Green Version]
- Grant, A.; Hashem, F.; Parveen, S. Salmonella and Campylobacter: Antimicrobial Resistance and Bacteriophage Control in Poultry. Food Microbiol. 2016, 53, 104–109. [Google Scholar] [CrossRef]
- Janež, N.; Loc-Carrillo, C. Use of Phages to Control Campylobacter spp. J. Microbiol. Methods 2013, 95, 68–75. [Google Scholar] [CrossRef] [Green Version]
- Ushanov, L.; Lasareishvili, B.; Janashia, I.; Zautner, A.E. Application of Campylobacter jejuni Phages: Challenges and Perspectives. Animals 2020, 10, 279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fortier, L.-C.; Sekulovic, O. Importance of Prophages to Evolution and Virulence of Bacterial Pathogens. Virulence 2013, 4, 354–365. [Google Scholar] [CrossRef]
- O’Flaherty, S.; Ross, R.P.; Coffey, A. Bacteriophage and Their Lysins for Elimination of Infectious Bacteria: Review Article. FEMS Microbiol. Rev. 2009, 33, 801–819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ackermann, H.W. Bacteriophage Observations and Evolution. Res. Microbiol. 2003, 154, 245–251. [Google Scholar] [CrossRef] [PubMed]
- Stone, E.; Campbell, K.; Grant, I.; McAuliffe, O. Understanding and Exploiting Phage-Host Interactions. Viruses 2019, 11, 567. [Google Scholar] [CrossRef] [Green Version]
- Nikolic, I.; Vukovic, D.; Gavric, D.; Cvetanovic, J.; Aleksic Sabo, V.; Gostimirovic, S.; Narancic, J.; Knezevic, P. An Optimized Checkerboard Method for Phage-Antibiotic Synergy Detection. Viruses 2022, 14, 1542. [Google Scholar] [CrossRef]
- Alaoui Mdarhri, H.; Benmessaoud, R.; Yacoubi, H.; Seffar, L.; Guennouni Assimi, H.; Hamam, M.; Boussettine, R.; Filali-Ansari, N.; Lahlou, F.A.; Diawara, I.; et al. Alternatives Therapeutic Approaches to Conventional Antibiotics: Advantages, Limitations and Potential Application in Medicine. Antibiotics 2022, 11, 1826. [Google Scholar] [CrossRef] [PubMed]
- Lin, D.M.; Koskella, B.; Lin, H.C. Phage Therapy: An Alternative to Antibiotics in the Age of Multi-Drug Resistance. World J. Gastrointest. Pharmacol. Ther. 2017, 8, 162. [Google Scholar] [CrossRef]
- Berchieri, A.; Lovell, M.A.; Barrow, P.A. The Activity in the Chicken Alimentary Tract of Bacteriophages Lytic for Salmonella typhimurium. Res. Microbiol. 1991, 142, 541–549. [Google Scholar] [CrossRef]
- Bardina, C.; Spricigo, D.A.; Cortés, P.; Llagostera, M. Significance of the Bacteriophage Treatment Schedule in Reducing Salmonella Colonization of Poultry. Appl. Environ. Microbiol. 2012, 78, 6600–6607. [Google Scholar] [CrossRef] [Green Version]
- Hong, S.S.; Jeong, J.; Lee, J.; Kim, S.; Min, W.; Myung, H. Therapeutic Effects of Bacteriophages against Salmonella gallinarum Infection in Chickens. J. Microbiol. Biotechnol. 2013, 23, 1478–1483. [Google Scholar] [CrossRef] [Green Version]
- Nabil, N.M.; Tawakol, M.M.; Hassan, H.M. Assessing the Impact of Bacteriophages in the Treatment of Salmonella in Broiler Chickens. Infect. Ecol. Epidemiol. 2018, 8, 1539056. [Google Scholar] [CrossRef] [PubMed]
- Clavijo, V.; Baquero, D.; Hernandez, S.; Farfan, J.C.; Arias, J.; Arévalo, A.; Donado-Godoy, P.; Vives-Flores, M. Phage Cocktail SalmoFREE® Reduces Salmonella on a Commercial Broiler Farm. Poult. Sci. 2019, 98, 5054–5063. [Google Scholar] [CrossRef]
- Wójcik, E.A.; Wojtasik, A.; Górecka, E.; Stańczyk, M. Application of Bacteriophage Preparation Bafasal® Іn Broiler Chickens Experimentally Exposed to Salmonella spp. Наукoвo-технічний бюлетень Державнoгo наукoвo-дoсліднoгo кoнтрoльнoгo інституту ветеринарних препаратів та кoрмoвих дoбавoк і Інституту біoлoгії тварин 2015, 16, 241–250. [Google Scholar]
- Proteon-Pharmaceuticals. Bafasal®. Available online: https://www.proteonpharma.com/bafasal-product/ (accessed on 28 November 2022).
- Nowaczek, A.; Urban-Chmiel, R.; Dec, M.; Puchalski, A.; Stępień-Pyśniak, D.; Marek, A.; Pyzik, E. Campylobacter spp. and Bacteriophages from Broiler Chickens: Characterization of Antibiotic Susceptibility Profiles and Lytic Bacteriophages. Microbiologyopen 2019, 8, e00784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wagenaar, J.A.; Bergen, M.A.P.V.; Mueller, M.A.; Wassenaar, T.M.; Carlton, R.M. Phage Therapy Reduces Campylobacter jejuni Colonization in Broilers. Vet. Microbiol. 2005, 109, 275–283. [Google Scholar] [CrossRef]
- Loc Carrillo, C.; Atterbury, R.J.; El-Shibiny, A.; Connerton, P.L.; Dillon, E.; Scott, A.; Connerton, I.F. Bacteriophage Therapy to Reduce Campylobacter jejuni Colonization of Broiler Chickens. Appl. Environ. Microbiol. 2005, 71, 6554–6563. [Google Scholar] [CrossRef] [Green Version]
- El-Shibiny, A.; Scott, A.; Timms, A.; Metawea, Y.; Connerton, P.; Connerton, I. Application of a Group II Campylobacter Bacteriophage to Reduce Strains of Campylobacter jejuni and Campylobacter coli Colonizing Broiler Chickens. J. Food Prot. 2009, 72, 733–740. [Google Scholar] [CrossRef] [Green Version]
- Carvalho, C.M.; Gannon, B.W.; Halfhide, D.E.; Santos, S.B.; Hayes, C.M.; Roe, J.M.; Azeredo, J. The in Vivo Efficacy of Two Administration Routes of a Phage Cocktail to Reduce Numbers of Campylobacter coli and Campylobacter jejuni in Chickens. BMC Microbiol. 2010, 10, 232. [Google Scholar] [CrossRef] [Green Version]
- Kittler, S.; Fischer, S.; Abdulmawjood, A.; Glünder, G.; Kleina, G. Effect of Bacteriophage Application on Campylobacter jejuni Loads in Commercial Broiler Flocks. Appl. Environ. Microbiol. 2013, 79, 7525–7533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fischer, S.; Kittler, S.; Klein, G.; Glünder, G. Impact of a Single Phage and a Phage Cocktail Application in Broilers on Reduction of Campylobacter jejuni and Development of Resistance. PLoS ONE 2013, 8, e78543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barrow, P.; Lovell, M.; Berchieru, A.J. Use of Lytic Bacteriophage for Control of Experimental Escherichia coli Septicemia and Meningitis in Chickens and Calves. Clin. Diagn. Lab. Immunol. 1998, 5, 89–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huff, W.E.; Huff, G.R.; Rath, N.C.; Balog, J.M.; Donoghue, A.M. Evaluation of Aerosol Spray and Intramuscular Injection of Bacteriophage to Treat an Escherichia coli Respiratory Infection. Poult. Sci. 2003, 82, 1108–1112. [Google Scholar] [CrossRef]
- Huff, G.; Huff, W.; Rath, N.; Donoghue, A. Critical Evaluation of Bacteriophage to Prevent and Treat Colibacillosis in Poultry. J. Ark. Acad. Sci. 2009, 63, 93–98. [Google Scholar]
- Xie, H.; Zhuang, X.; Kong, J.; Ma, G.; Zhang, H. Bacteriophage Esc-A Is an Efficient Therapy for Escherichia coli 3-1 Caused Diarrhea in Chickens. J. Gen. Appl. Microbiol. 2005, 51, 159–163. [Google Scholar] [CrossRef] [PubMed]
- Marek, A.; Pyzik, E.; Stępień-Pyśniak, D.; Urban-Chmiel, R.; Nowaczek, A. Characterization of Bacteriophages and Their Carriage in Staphylococcus aureus Isolated from Broilers in Poland. Br. Poult. Sci. 2019, 60, 373–380. [Google Scholar] [CrossRef] [PubMed]
- Saint-Cyr, M.J.; Guyard-Nicodème, M.; Messaoudi, S.; Chemaly, M.; Cappelier, J.M.; Dousset, X.; Haddad, N. Recent Advances in Screening of Anti-Campylobacter Activity in Probiotics for Use in Poultry. Front. Microbiol. 2016, 7, 553. [Google Scholar] [CrossRef] [Green Version]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. Expert Consensus Document: The International Scientific Association for Probiotics and Prebiotics Consensus Statement on the Scope and Appropriate Use of the Term Probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tellez, G.; Pixley, C.; Wolfenden, R.E.; Layton, S.L.; Hargis, B.M. Probiotics/Direct Fed Microbials for Salmonella Control in Poultry. Food Res. Int. 2012, 45, 628–633. [Google Scholar] [CrossRef]
- Farnell, M.B.; Donoghue, A.M.; Solis De Los Santos, F.; Blore, P.J.; Hargis, B.M.; Tellez, G.; Donoghue, D.J. Upregulation of Oxidative Burst and Degranulation in Chicken Heterophils Stimulated with Probiotic Bacteria. Poult. Sci. 2006, 85, 1900–1906. [Google Scholar] [CrossRef]
- Zheng, J.; Wittouck, S.; Salvetti, E.; Franz, C.M.A.P.; Harris, H.M.B.; Mattarelli, P.; O’Toole, P.W.; Pot, B.; Vandamme, P.; Walte, J.; et al. A Taxonomic Note on the Genus Lactobacillus: Description of 23 Novel Genera, Emended Description of the Genus Lactobacillus Beijerinck 1901, and Union of Lactobacillaceae and Leuconostocaceae. Microbiol. Soc. 2020, 70, 2782–2858. [Google Scholar] [CrossRef]
- Vicente, J.L.; Aviña, L.; Torres-Rodriguez, A.; Hargis, B.; Tellez, G. Effect of a Lactobacillus Spp-Based Probiotic Culture Product on Broiler Chicks Performance under Commercial Conditions. Int. J. Poult. Sci. 2007, 6, 154–156. [Google Scholar] [CrossRef] [Green Version]
- Higgins, S.E.; Higgins, J.P.; Wolfenden, A.D.; Henderson, S.N.; Torres-Rodriguez, A.; Tellez, G.; Hargis, B. Evaluation of a Lactobacillus-Based Probiotic Culture for the Reduction of Salmonella Enteritidis in Neonatal Broiler Chicks. Poult. Sci. 2008, 87, 27–31. [Google Scholar] [CrossRef]
- Shivaramaiah, S.; Pumford, N.R.; Morgan, M.J.; Wolfenden, R.E.; Wolfenden, A.D.; Torres-Rodriguez, A.; Hargis, B.M.; Tellez, G. Evaluation of Bacillus Species as Potential Candidates for Direct-Fed Microbials in Commercial Poultry. Poult. Sci. 2011, 90, 1574–1580. [Google Scholar] [CrossRef]
- Higgins, J.P.; Higgins, S.E.; Wolfenden, A.D.; Henderson, S.N.; Torres-Rodriguez, A.; Vicente, J.L.; Hargis, B.M.; Tellez, G. Effect of Lactic Acid Bacteria Probiotic Culture Treatment Timing on Salmonella Enteritidis in Neonatal Broilers. Poult. Sci. 2010, 89, 243–247. [Google Scholar] [CrossRef]
- Sadeghi, A.A.; Shawrang, P.; Shakorzadeh, S. Immune Response of Salmonella Challenged Broiler Chickens Fed Diets Containing Gallipro®, a Bacillus Subtilis Probiotic. Probiotics Antimicrob. Proteins 2015, 7, 24–30. [Google Scholar] [CrossRef] [PubMed]
- Carter, A.; Adams, M.; La Ragione, R.M.; Woodward, M.J. Colonisation of Poultry by Salmonella Enteritidis S1400 Is Reduced by Combined Administration of Lactobacillus salivarius 59 and Enterococcus faecium PXN-33. Vet. Microbiol. 2017, 199, 100–107. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.; Chousalkar, K.K. Salmonella Typhimurium Infection Disrupts but Continuous Feeding of Bacillus Based Probiotic Restores Gut Microbiota in Infected Hens. J. Anim. Sci. Biotechnol. 2020, 11, 29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Price, P.T.; Gaydos, T.A.; Berghaus, R.D.; Baxter, V.; Hofacre, C.L.; Sims, M.D. Salmonella Enteritidis Reduction in Layer Ceca with a Bacillus Probiotic. Vet. World 2020, 13, 184–187. [Google Scholar] [CrossRef] [Green Version]
- Oh, J.K.; Pajarillo, E.A.B.; Chae, J.P.; Kim, I.H.; Kang, D.K. Protective Effects of Bacillus subtilis against Salmonella Infection in the Microbiome of Hy-Line Brown Layers. Asian-Australas. J. Anim. Sci. 2017, 30, 1332–1339. [Google Scholar] [CrossRef]
- Chen, X.; Ishfaq, M.; Wang, J. Effects of Lactobacillus salivarius Supplementation on the Growth Performance, Liver Function, Meat Quality, Immune Responses and Salmonella pullorum Infection Resistance of Broilers Challenged with Aflatoxin B1. Poult. Sci. 2022, 101, 101651. [Google Scholar] [CrossRef]
- Willis, W.L.; Reid, L. Investigating the Effects of Dietary Probiotic Feeding Regimens on Broiler Chicken Production and Campylobacter jejuni Presence. Poult. Sci. 2008, 87, 606–611. [Google Scholar] [CrossRef]
- Santini, C.; Baffoni, L.; Gaggia, F.; Granata, M.; Gasbarri, R.; Di Gioia, D.; Biavati, B. Characterization of Probiotic Strains: An Application as Feed Additives in Poultry against Campylobacter jejuni. Int. J. Food Microbiol. 2010, 141, S98–S108. [Google Scholar] [CrossRef]
- Nishiyama, K.; Seto, Y.; Yoshioka, K.; Kakuda, T.; Takai, S.; Yamamoto, Y.; Mukai, T. Lactobacillus Gasseri SBT2055 Reduces Infection by and Colonization of Campylobacter jejuni. PLoS ONE 2014, 9, e108827. [Google Scholar] [CrossRef]
- Nishiyama, K.; Nakazato, A.; Ueno, S.; Seto, Y.; Kakuda, T.; Takai, S.; Yamamoto, Y.; Mukai, T. Cell Surface-associated Aggregation-promoting Factor from Lactobacillus gasseri SBT 2055 Facilitates Host Colonization and Competitive Exclusion of Campylobacter jejuni. Mol. Microbiol. 2015, 98, 712–726. [Google Scholar] [CrossRef] [PubMed]
- Arsi, K.; Donoghue, A.M.; Woo-Ming, A.; Blore, P.J.; Donoghue, D.J. The Efficacy of Selected Probiotic and Prebiotic Combinations in Reducing Campylobacter Colonization in Broiler Chickens. J. Appl. Poult. Res. 2015, 24, 327–334. [Google Scholar] [CrossRef]
- Mohan, V. The Role of Probiotics in the Inhibition of Campylobacter jejuni Colonization and Virulence Attenuation. Eur. J. Clin. Microbiol. Infect. Dis. 2015, 34, 1503–1513. [Google Scholar] [CrossRef] [PubMed]
- Abd El-Ghany, W.A.; Shaalan, M.; Salem, H.M. Nanoparticles Applications in Poultry Production: An Updated Review. Worlds. Poult. Sci. J. 2021, 77, 1001–1025. [Google Scholar] [CrossRef]
- Yusof, H.M.; Rahman, N.A.; Mohamad, R.; Zaidan, U.H.; Samsudin, A.A. Antibacterial Potential of Biosynthesized Zinc Oxide An In Vitro Study. Animals 2021, 11, 2093. [Google Scholar] [CrossRef]
- Ye, Q.; Chen, W.; Huang, H.; Tang, Y.; Wang, W.; Meng, F.; Wang, H.; Zheng, Y. Iron and Zinc Ions, Potent Weapons against Multidrug-Resistant Bacteria. Appl. Microbiol. Biotechnol. 2020, 104, 5213–5227. [Google Scholar] [CrossRef]
- Siddiqui, S.A.; Bahmid, N.A.; Taha, A.; Abdel-Moneim, A.M.E.; Shehata, A.M.; Tan, C.; Kharazmi, M.S.; Li, Y.; Assadpour, E.; Castro-Muñoz, R.; et al. Bioactive-Loaded Nanodelivery Systems for the Feed and Drugs of Livestock; Purposes, Techniques and Applications. Adv. Colloid Interface Sci. 2022, 308, 102772. [Google Scholar] [CrossRef] [PubMed]
- Meimandipour, A.; Emamzadeh, A.N.; Soleimani, A. Effects of Nanoencapsulated Aloe Vera, Dill and Nettle Root Extract as Feed Antibiotic Substitutes in Broiler Chickens. Arch. Anim. Breed. 2017, 60, 1–7. [Google Scholar] [CrossRef]
- Soni, M.; Mehta, P.; Soni, A.; Goswami, G.K. Green Nanoparticles: Synthesis and Applications. IOSR J. Biotechnol. Biochem. 2018, 4, 78–83. [Google Scholar] [CrossRef]
- Acevedo-Villanueva, K.Y.; Akerele, G.O.; Hakeem, W.G.A.; Renu, S.; Shanmugasundaram, R.; Selvaraj, R.K. A Novel Approach against Salmonella: A Review of Polymeric Nanoparticle Vaccines for Broilers and Layers. Vaccines 2021, 9, 1041. [Google Scholar] [CrossRef]
- Tigabu, B.; Getachew, A. Treatment of Antibiotic-Resistant Bacteria by Nanoparticles: Current Approaches and Prospects. Ann. Adv. Chem. 2022, 6, 001–009. [Google Scholar] [CrossRef]
- Kazemnia, A.; Ahmadi, M.; Mardani, K.; Moradi, M.; Darvishzadeh, R. Combined Efficacy of Silver Nanoparticles and Commercial Antibiotics on Different Phylogenetic Groups of Escherichia coli. J. Hell. Vet. Med. Soc. 2019, 70, 1647–1654. [Google Scholar] [CrossRef] [Green Version]
- Lopez-Carrizales, M.; Velasco, K.I.; Castillo, C.; Flores, A.; Magaña, M.; Martinez-Castanon, G.A.; Martinez-Gutierrez, F. In Vitro Synergism of Silver Nanoparticles with Antibiotics as an Alternative Treatment in Multiresistant Uropathogens. Antibiotics 2018, 7, 50. [Google Scholar] [CrossRef] [Green Version]
- Ahmadi, A.; Ahmadi, P.; Ehsani, A. Development of an Active Packaging System Containing Zinc Oxide Nanoparticles for the Extension of Chicken Fillet Shelf Life. Food Sci. Nutr. 2020, 8, 5461–5473. [Google Scholar] [CrossRef] [PubMed]
- Devi, J.S.; Bhimba, B.V. Antibacterial and Antifungal Activity of Silver Nanoparticles Synthesized Using Hypnea Muciformis. Biosci. Biotechnol. Res. Asia 2014, 11, 235–238. [Google Scholar] [CrossRef]
- Finley, P.J.; Norton, R.; Austin, C.; Mitchell, A.; Zank, S.; Durham, P. Unprecedented Silver Resistance in Clinically Isolated Enterobacteriaceae: Major Implications for Burn and Wound Management. Antimicrob. Agents Chemother. 2015, 59, 4734–4741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salas-Orozco, M.; Niño-Martínez, N.; Martínez-Castañón, G.A.; Méndez, F.T.; Jasso, M.E.C.; Ruiz, F. Mechanisms of Resistance to Silver Nanoparticles in Endodontic Bacteria: A Literature Review. J. Nanomater. 2019, 2019, 7630316. [Google Scholar] [CrossRef] [Green Version]
- Salem, H.M.; Ismael, E.; Shaalan, M. Evaluation of the Effects of Silver Nanoparticles against Experimentally Induced Necrotic Enteritis in Broiler Chickens. Int. J. Nanomed. 2021, 16, 6783–6796. [Google Scholar] [CrossRef]
- Song, Z.; Lv, J.; Sheikhahmadi, A.; Uerlings, J.; Everaert, N. Attenuating Effect of Zinc and Vitamin E on the Intestinal Oxidative Stress Induced by Silver Nanoparticles in Broiler Chickens. Biol. Trace Elem. Res. 2017, 180, 306–313. [Google Scholar] [CrossRef]
- Ali, N.M.; Bakheet, A.A. Evaluation of the Inhibitory Effect of Chitosan Nanoparticles on Biofilm Forming Escherichia coli Isolated from Omphalitis Cases. J. Adv. Vet. Res. 2020, 10, 213–218. [Google Scholar]
- Patric Joshua, P.; Valli, C.; Balakrishnan, V. Effect of in Ovo Supplementation of Nano Forms of Zinc, Copper, and Selenium on Post-Hatch Performance of Broiler Chicken. Vet. World 2016, 9, 287–294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ningsih, N.; Ariyadi, B.; Dono, N.D.; Supadmo; Zuprizal. The Effect of Nanoencapsulated Phaleria Macrocarpa Fruits Extract in Drinking Water on Jejunal Histomorphology of Broiler Chickens. Trop. Anim. Sci. J. 2019, 42, 106–112. [Google Scholar] [CrossRef]
- Wang, L.; Hu, C.; Shao, L. The Antimicrobial Activity of Nanoparticles: Present Situation and Prospects for the Future. Int. J. Nanomed. 2017, 12, 1227–1249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vadalasetty, K.P.; Lauridsen, C.; Engberg, R.M.; Vadalasetty, R.; Kutwin, M.; Chwalibog, A.; Sawosz, E. Influence of Silver Nanoparticles on Growth and Health of Broiler Chickens after Infection with Campylobacter jejuni. BMC Vet. Res. 2018, 14, 1. [Google Scholar] [CrossRef]
- Li, Z.; Hu, Y.; Yang, Y.; Lu, Z.; Wang, Y. Antimicrobial Resistance in Livestock: Antimicrobial Peptides Provide a New Solution for a Growing Challenge. Anim. Front. 2018, 8, 21–29. [Google Scholar] [CrossRef]
- Moretta, A.; Scieuzo, C.; Petrone, A.M.; Salvia, R.; Manniello, M.D.; Franco, A.; Lucchetti, D.; Vassallo, A.; Vogel, H.; Sgambato, A.; et al. Antimicrobial Peptides: A New Hope in Biomedical and Pharmaceutical Fields. Front. Cell. Infect. Microbiol. 2021, 11, 668632. [Google Scholar] [CrossRef]
- Abdi, M.; Mirkalantari, S.; Amirmozafari, N. Bacterial Resistance to Antimicrobial Peptides. J. Pept. Sci. 2019, 25, e3210. [Google Scholar] [CrossRef]
- Kumar, R.; Ali, S.A.; Singh, S.K.; Bhushan, V.; Mathur, M.; Jamwal, S.; Mohanty, A.K.; Kaushik, J.K.; Kumar, S. Antimicrobial Peptides in Farm Animals: An Updated Review on Its Diversity, Function, Modes of Action and Therapeutic Prospects. Vet. Sci. 2020, 7, 206. [Google Scholar] [CrossRef]
- Dijksteel, G.S.; Ulrich, M.M.W.; Middelkoop, E.; Boekema, B.K.H.L. Review: Lessons Learned from Clinical Trials Using Antimicrobial Peptides (AMPs). Front. Microbiol. 2021, 12, 616979. [Google Scholar] [CrossRef]
- Torres-Acosta, M.A.; Clavijo, V.; Vaglio, C.; González-Barrios, A.F.; Vives-Flórez, M.J.; Rito-Palomares, M. Economic Evaluation of the Development of a Phage Therapy Product for the Control of Salmonella in Poultry. Biotechnol. Prog. 2019, 35, e2852. [Google Scholar] [CrossRef]
- Greer, G.G. Bacteriophage Control of Foodborne Bacteria. J. Food Prot. 2005, 68, 1102–1111. [Google Scholar] [CrossRef]
- Oechslin, F. Resistance Development to Bacteriophages Occurring during Bacteriophage Therapy. Viruses 2018, 10, 351. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Li, Z. CRISPR-Cas Systems: Overview, Innovations and Applications in Human Disease Research and Gene Therapy. Comput. Struct. Biotechnol. J. 2020, 18, 2401–2415. [Google Scholar] [CrossRef]
- Sulakvelidze, A. Using Lytic Bacteriophages to Eliminate or Significantly Reduce Contamination of Food by Foodborne Bacterial Pathogens. J. Sci. Food Agric. 2013, 93, 3137–3146. [Google Scholar] [CrossRef] [PubMed]
- Wernicki, A.; Nowaczek, A.; Urban-Chmiel, R. Bacteriophage Therapy to Combat Bacterial Infections in Poultry. Virol. J. 2017, 14, 179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gómez-Gómez, C.; Blanco-Picazo, P.; Brown-Jaque, M.; Quirós, P.; Rodríguez-Rubio, L.; Cerdà-Cuellar, M.; Muniesa, M. Infectious Phage Particles Packaging Antibiotic Resistance Genes Found in Meat Products and Chicken Feces. Sci. Rep. 2019, 9, 13281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panáček, A.; Kvítek, L.; Smékalová, M.; Večeřová, R.; Kolář, M.; Röderová, M.; Dyčka, F.; Šebela, M.; Prucek, R.; Tomanec, O.; et al. Bacterial Resistance to Silver Nanoparticles and How to Overcome It. Nat. Nanotechnol. 2018, 13, 65–71. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Carlsson, F.; Edman, M.; Hummelgård, M.; Jonsson, B.G.; Bylund, D.; Olin, H. Escherichia coli Bacteria Develop Adaptive Resistance to Antibacterial ZnO Nanoparticles. Adv. Biosyst. 2018, 2, 1800019. [Google Scholar] [CrossRef] [PubMed]
- Larkin Mchugh, G.; Moellering, R.C.; Hopkins, C.C.; Swartz, M.N. Salmonella Typhimurium Resistant to Silver Nitrate, Chloramphenicol, and Ampicillin: A New Threat in Burn Units? Lancet 1975, 305, 235–240. [Google Scholar] [CrossRef]
- Fisinin, V.I.; Miroshnikov, S.A.; Sizova, E.A.; Ushakov, A.S.; Miroshnikova, E.P. Metal Particles as Trace-Element Sources: Current State and Future Prospects. Worlds Poult. Sci. J. 2018, 74, 523–540. [Google Scholar] [CrossRef]
- Ognik, K.; Cholewińska, E.; Czech, A.; Kozłowski, K.; Wlazło, Ł.; Nowakowicz-Dębek, B.; Szlązak, R.; Tutaj, K. Effect of Silver Nanoparticles on the Immune, Redox, and Lipid Status of Chicken Blood. Czech J. Anim. Sci. 2016, 61, 450–461. [Google Scholar] [CrossRef] [Green Version]
- Von Wintersdorff, C.J.H.; Penders, J.; Van Niekerk, J.M.; Mills, N.D.; Majumder, S.; Van Alphen, L.B.; Savelkoul, P.H.M.; Wolffs, P.F.G. Dissemination of Antimicrobial Resistance in Microbial Ecosystems through Horizontal Gene Transfer. Front. Microbiol. 2016, 7, 173. [Google Scholar] [CrossRef] [PubMed] [Green Version]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abreu, R.; Semedo-Lemsaddek, T.; Cunha, E.; Tavares, L.; Oliveira, M. Antimicrobial Drug Resistance in Poultry Production: Current Status and Innovative Strategies for Bacterial Control. Microorganisms 2023, 11, 953. https://doi.org/10.3390/microorganisms11040953
Abreu R, Semedo-Lemsaddek T, Cunha E, Tavares L, Oliveira M. Antimicrobial Drug Resistance in Poultry Production: Current Status and Innovative Strategies for Bacterial Control. Microorganisms. 2023; 11(4):953. https://doi.org/10.3390/microorganisms11040953
Chicago/Turabian StyleAbreu, Raquel, Teresa Semedo-Lemsaddek, Eva Cunha, Luís Tavares, and Manuela Oliveira. 2023. "Antimicrobial Drug Resistance in Poultry Production: Current Status and Innovative Strategies for Bacterial Control" Microorganisms 11, no. 4: 953. https://doi.org/10.3390/microorganisms11040953