
The Effects of Antiperspirant Aluminum Chlorohydrate on the Development of Antibiotic Resistance in Staphylococcus epidermidis

 , , ,
, , , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Growth Conditions
2.2. Detection of Antibiotic Resistance Genes
2.3. Determination of Minimum Inhibitory Concentration Values
2.4. Aluminum Chlorohydrate Exposure and Isolation of Resistant Bacteria against Test Antibiotics
2.5. Quantitative Reverse Transcriptase PCR Analysis of Target Genes
3. Results and Discussion
3.1. Detection of Antibiotic Resistance Genes
3.2. MIC Values of Aluminum Chlorohydrate
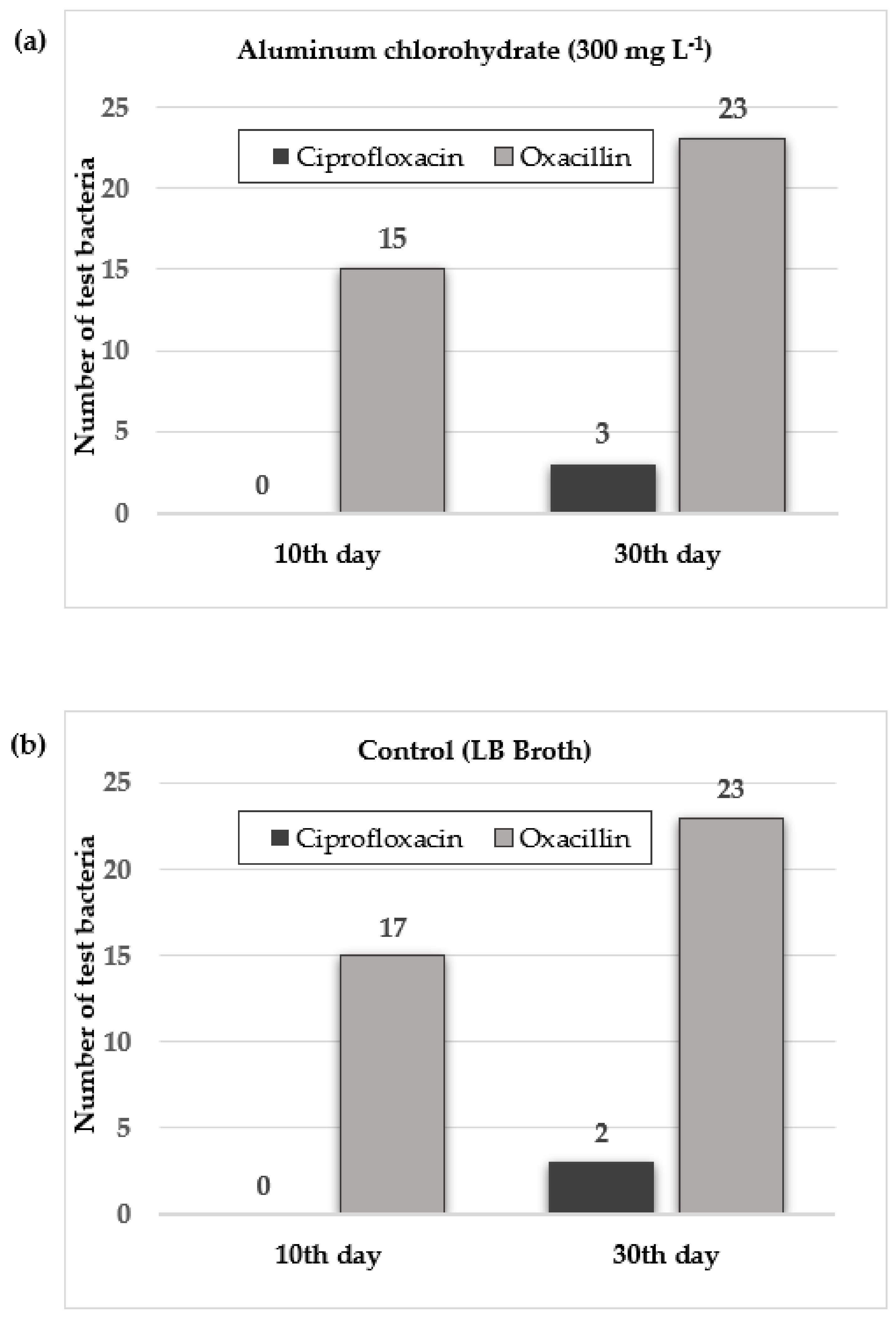
3.3. Antibiotic Resistance Development after Aluminum Chlorohydrate Exposure
3.4. Increase in the Minimum Inhibitory Concentration Values of the Test Bacteria
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Landers, T.F.; Cohen, B.; Wittum, T.E.; Larson, E.L. A review of antibiotic use in food animals: Perspective, policy, and potential. Public Health Rep. 2012, 127, 4–22. [Google Scholar] [CrossRef] [Green Version]
- Jin, M.; Lu, J.; Chen, Z.; Nguyen, S.H.; Mao, L.; Li, J.; Yuan, Z.; Guo, J. Antidepressant fluoxetine induces multiple antibiotics resistance in Escherichia coli via ROS-mediated mutagenesis. Environ. Int. 2018, 120, 421–430. [Google Scholar] [CrossRef] [PubMed]
- Morehead, M.S.; Scarbrough, C. Emergence of global antibiotic resistance. Prim. Care 2018, 45, 467–484. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention. Antibiotic Resistance Threats in the United States. 2019. Available online: https://www.cdc.gov/drugresistance/pdf/threats-report/2019-ar-threats-report508.pdf (accessed on 15 October 2021).
- World Health Organization. Antibiotic Resistance. Available online: https://www.who.int/news-room/factsheets/detail/antibiotic-resistance (accessed on 23 August 2021).
- Gurpinar, S.S.; Kart, D.; Eryilmaz, M. The effects of antidepressants fluoxetine, sertraline, and amitriptyline on the development of antibiotic resistance in Acinetobacter baumannii. Arch. Microbiol. 2022, 204, 230. [Google Scholar] [CrossRef]
- Haran, J.P.; McCormick, B.A. Aging, frailty, and the microbiome-how dysbiosis influences human aging and disease. Gastroenterology 2021, 160, 507–523. [Google Scholar] [CrossRef] [PubMed]
- Shanahan, F.; Ghosh, T.S.; O’toole, P.W. The healthy microbiome—What is the definition of a healthy gut microbiome? Gastroenterology 2021, 160, 483–494. [Google Scholar] [CrossRef] [PubMed]
- Otto, M. Staphylococcus epidermidis—The ‘accidental’ pathogen. Nat. Rev. Microbiol. 2009, 7, 555–567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brooks, G.F.; Carroll, K.C.; Butel, J.S.; Morse, S.A.; Mietzner, T.A. Jawetz, Melnickk ve Adelberg Tıbbi Mikrobiyoloji; Yenen, O.Ş., Ed.; Nobel Kitapevleri: Ankara, Turkey, 2014; p. 167. [Google Scholar]
- Fournière, M.; Latire, T.; Souak, D.; Feuilloley, M.G.J.; Bedoux, G. Staphylococcus epidermidis and Cutibacterium acnes: Two major sentinels of skin microbiota and the influence of cosmetics. Microorganisms 2020, 8, 1752. [Google Scholar] [CrossRef]
- Vuong, C.; Otto, M. Staphylococcus epidermidis infections. Microbes Infect. 2002, 4, 481–489. [Google Scholar] [CrossRef] [PubMed]
- Eryilmaz, M.; Gurpinar, S.S. Investigation of the antibacterial efficacy of some commonly used antiseptics in hospitals against biofilm forming and non-biofilm forming Staphylococcus epidermidis strains. J. Fac. Pharm. Ankara 2017, 41, 1–8. [Google Scholar] [CrossRef]
- Cerca, N.; Martins, S.; Cerca, F.; Jefferson, K.K.; Pier, G.B.; Oliveira, R.; Azeredo, J. Comparative assessment of antibiotic susceptibility of coagulase-negative Staphylococci in biofilm versus planktonic culture as assessed by bacterial enumeration or rapid XTT colorimetry. J. Antimicrob. Chemother. 2005, 56, 331–336. [Google Scholar] [CrossRef]
- Morgenstern, M.; Erichsen, C.; Von Rüden, C.; Metsemakers, W.J.; Kates, S.L.; Moriarty, T.F.; Hungerer, S. Staphylococcal orthopaedic device-related infections in older patients. Injury 2016, 47, 1427–1434. [Google Scholar] [CrossRef]
- Sabaté Brescó, M.; Harris, L.G.; Thompson, K.; Stanic, B.; Morgenstern, M.; O’Mahony, L.; Richards, R.G.; Moriarty, T.F. Pathogenic mechanisms and host interactions in Staphylococcus epidermidis device-related infection. Front. Microbiol. 2017, 8, 1401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bouiller, K.; Ilic, D.; Wicky, P.H.; Cholley, P.; Chirouze, C.; Bertrand, X. Spread of clonal linezolid-resistant Staphylococcus epidermidis in an intensive care unit associated with linezolid exposure. Eur. J. Clin. Microbiol. Infect. Dis. 2020, 39, 1271–1277. [Google Scholar] [CrossRef] [PubMed]
- Callewaert, C.; Hutapea, P.; Van de Wiele, T.; Boon, N. Deodorants and antiperspirants affect the axillary bacterial community. Arch. Dermatol. Res. 2014, 306, 701–710. [Google Scholar] [CrossRef]
- Glatz, M.; Jo, J.H.; Kennedy, E.A.; Polley, E.C.; Segre, J.A.; Simpson, E.L.; Kong, H.H. Emollient use alters skin barrier and microbes in infants at risk for developing atopic dermatitis. PLoS ONE 2018, 13, e0192443. [Google Scholar] [CrossRef] [Green Version]
- Aras, A.; Eryilmaz, M. Microbiological contamination of cosmetic products. J. Fac. Pharm. Ankara 2022, 46, 262–276. [Google Scholar]
- Holzle, E. Antiperspirants. In Dermatopharmacology of Topical Preparations; Springer: Berlin/Heidelberg, Germany, 2000; pp. 401–402. [Google Scholar]
- Martini, M.C. Déodorants et antitranspirants [Deodorants and antiperspirants]. Ann. Dermatol. Venereol. 2020, 147, 387–395. [Google Scholar] [CrossRef] [PubMed]
- Clinical and Laboratory Standards Institute (CLSI). Performance Standartds for Antimicrobial Susceptibility Testing, 30th ed.; CLSI Supplement M100; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2020. [Google Scholar]
- Vandecasteele, S.J.; Peetermans, W.E.; Merckx, R.; Van Eldere, J. Quantification of expression of Staphylococcus epidermidis housekeeping genes with Taqman quantitative PCR during in vitro growth and under different conditions. J. Bacteriol. 2001, 183, 7094–7101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamada, M.; Yoshida, J.; Hatou, S.; Yoshida, T.; Minagawa, Y. Mutations in the quinolone resistance determining region in Staphylococcus epidermidis recovered from conjunctiva and their association with susceptibility to various fluoroquinolones. Br. J. Ophthalmol. 2008, 92, 848–851. [Google Scholar] [CrossRef] [Green Version]
- Betanzos-Cabrera, G.; Juárez-Verdayes, M.A.; González-González, G.; Cancino-Díaz, M.E.; Cancino-Díaz, J.C. Gatifloxacin, moxifloxacin, and balofloxacin resistance due to mutations in the gyrA and parC genes of Staphylococcus epidermidis strains isolated from patients with endophthalmitis, corneal ulcers and conjunctivitis. Ophthalmic Res. 2009, 42, 43–48. [Google Scholar] [CrossRef]
- Juárez-Verdayes, M.A.; Parra-Ortega, B.; Hernández-Rodríguez, C.; Betanzos-Cabrera, G.; Rodríguez-Martínez, S.; Cancino-Diaz, M.E.; Cancino-Diaz, J.C. Identification and expression of nor efflux family genes in Staphylococcus epidermidis that act against gatifloxacin. Microb. Pathog. 2012, 52, 318–325. [Google Scholar] [CrossRef] [PubMed]
- Sully, E.K.; Malachowa, N.; Elmore, B.O.; Alexander, S.M.; Femling, J.K.; Gray, B.M.; DeLeo, F.R.; Otto, M.; Cheung, A.L.; Edwards, B.S.; et al. Selective chemical inhibition of agr quorum sensing in Staphylococcus aureus promotes host defense with minimal impact on resistance. PLoS Pathog. 2014, 10, e1004174. [Google Scholar] [CrossRef] [PubMed]
- Eladli, M.G.; Alharbi, N.S.; Khaled, J.M.; Kadaikunnan, S.; Alobaidi, A.S.; Alyahya, S.A. Antibiotic-resistant Staphylococcus epidermidis isolated from patients and healthy students comparing with antibiotic-resistant bacteria isolated from pasteurized milk. Saudi J. Biol. Sci. 2019, 26, 1285–1290. [Google Scholar] [CrossRef]
- Zen, J.M.; Yang, T.H.; Kumar, A.S.; Chen, Y.J.; Hsu, J.C.; Shih, Y. Detection of aluminum chlorohydrate content in antiperspirant deodorants using screen-printed silver electrodes by one drop analysis. Electroanalysis 2009, 21, 2272–2276. [Google Scholar] [CrossRef]
- Scientific Committee on Consumer Safety (SCCS). The SCCS Notes of Guidance for the Testing of Cosmetic Ingredients and Their Safety Evaluation, 10th ed.; SCCS/1628/18: 24-25; SCCS: Luxembourg, 2018. [Google Scholar]
- Kart, D.; Gurpinar, S.S.; Eryilmaz, M. Assessment of the anti-quorum sensing effect of Lactobacillus sp. metabolites on expression levels of QS-related genes in Pseudomonas aeruginosa PAO1. Turk. Bull. Hyg. Exp. Biol. 2020, 77, 311–318. [Google Scholar] [CrossRef]
- Yilmaz, E. Quinolones. Turkiye Klinikleri J. Inf. Dis. Special Topics 2017, 10, 99–105. [Google Scholar]
- Thai, T.; Salisbury, B.H.; Zito, P.M. Ciprofloxacin; [Updated 2 September 2022]; StatPearls: Treasure Island, FL, USA. Available online: https://www.ncbi.nlm.nih.gov/books/NBK535454/ (accessed on 15 October 2022).
- Shariati, A.; Arshadi, M.; Khosrojerdi, M.A.; Abedinzadeh, M.; Ganjalishahi, M.; Maleki, A.; Heidary, M.; Khoshnood, S. The resistance mechanisms of bacteria against ciprofloxacin and new approaches for enhancing the efficacy of this antibiotic. Front. Public Health 2022, 10, 1025633. [Google Scholar] [CrossRef] [PubMed]
- DeMarco, C.E.; Cushing, L.A.; Frempong-Manso, E.; Seo, S.M.; Jaravaza, T.A.; Kaatz, G.W. Efflux-related resistance to norfloxacin, dyes, and biocides in bloodstream isolates of Staphylococcus aureus. Antimicrob. Agents Chemother. 2007, 51, 3235–3239. [Google Scholar] [CrossRef] [Green Version]
- Qin, R.; Xiao, K.; Li, B.; Jiang, W.; Peng, W.; Zheng, J.; Zhou, H. The combination of catechin and epicatechin callate from Fructus Crataegi potentiates beta-lactam antibiotics against methicillin-resistant Staphylococcus aureus (MRSA) in vitro and in vivo. Int. J. Mol. Sci. 2013, 14, 1802–1821. [Google Scholar] [CrossRef] [Green Version]
- Bostanmaneshrad, A.; Nowroozi, J.; Eslami, G.; Hashemi, A. The expression of efflux pump genes in methicillin-resistant Staphylococcus aureus (MRSA) strains isolated from blood cultures in Iran. Arch. Clin. Infect. Dis. 2020, 15, e99804. [Google Scholar] [CrossRef]
- Sancak, B. Resistance mechanisms of MRSA: Epidemiology in the World and Turkey. Ankem. Derg. 2012, 26, 38–47. [Google Scholar]
- Çiftci, İ.H.; Altindis, M.; Cetinkaya, Z.; Asik, G.; Aktepe, O.C. Investigation of mecA genes in Staphylococcus strains isolated from clinical samples. Kocatepe Med. J. 2009, 10, 17–20. [Google Scholar]
- Xu, Z.; Shah, H.N.; Misra, R.; Chen, J.; Zhang, W.; Liu, Y.; Cutler, R.R.; Mkrtchyan, H.V. The prevalence, antibiotic resistance and mecA characterization of coagulase negative Staphylococci recovered from non-healthcare settings in London, UK. Antimicrob. Resist. Infect. Control. 2018, 7, 73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, Y.S.; Zhang, R.K.; Liu, Z.H.; Li, B.Z.; Yuan, Y.J. Microbial adaptation to enhance stress tolerance. Front. Microbiol. 2022, 13, 888746. [Google Scholar] [CrossRef]
- Wang, Y.; Lu, J.; Zhang, S.; Li, J.; Mao, L.; Yuan, Z.; Bond, P.L.; Guo, J. Non-antibiotic pharmaceuticals promote the transmission of multidrug resistance plasmids through intra- and intergenera conjugation. ISME J. 2021, 15, 2493–2508. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Exposure Day | Test Antibiotic | Aluminum Chlorohydrate (300 mg L−1) | Control * |
|---|---|---|---|
| 10th | CIP | - | - |
| OXA | 1, 2, 3, 5, 12, 13, 14, 15, 16, 17, 18, 20, 21, 22, 24 | 1, 2, 3, 4, 6, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 22 | |
| 30th | CIP | 18, 22, 24 | 14, 24 |
| OXA | 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 24 | 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 24 |
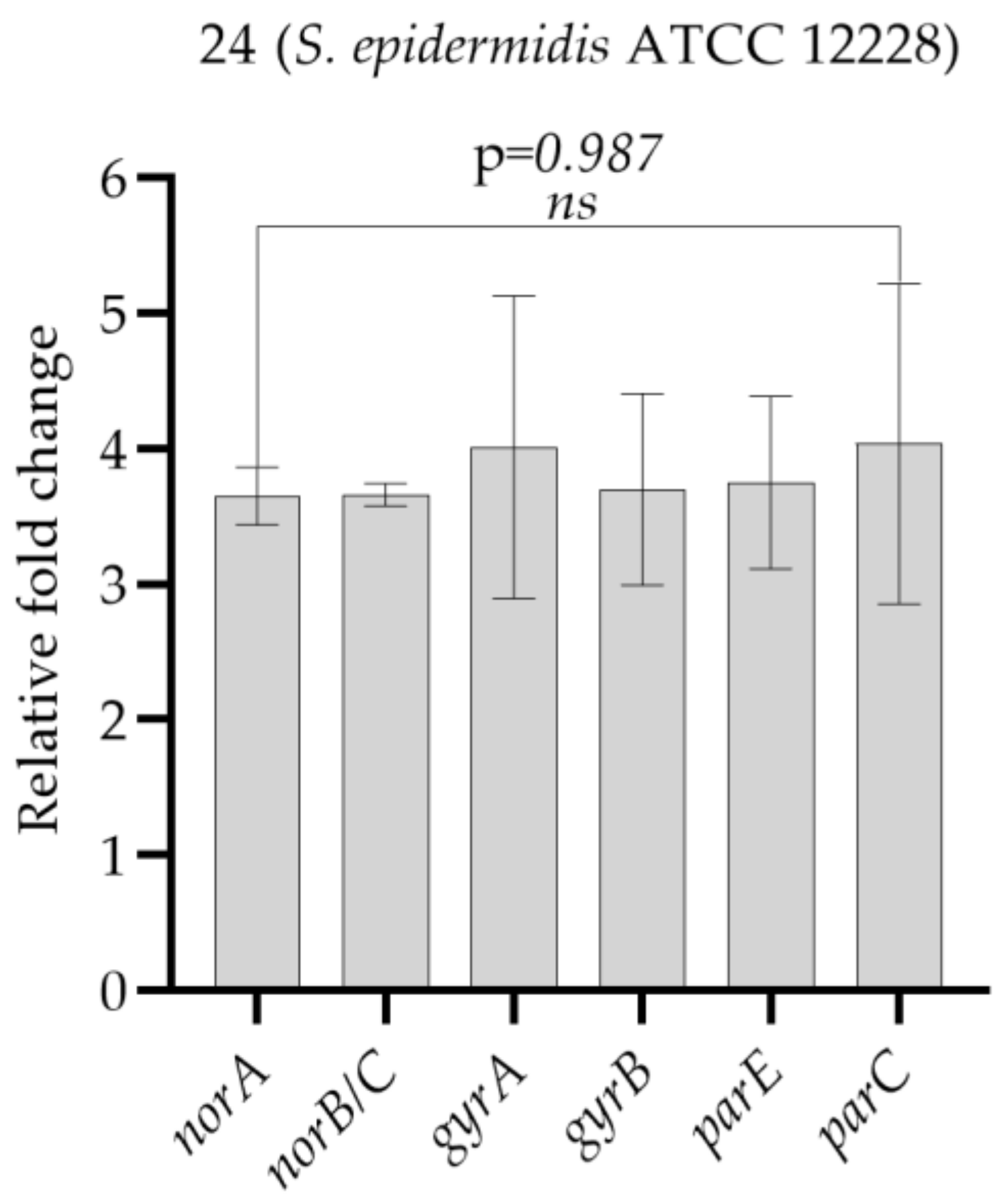
| Test Bacteria | norA | norB/C | gyrA | gyrB | parE | parC |
|---|---|---|---|---|---|---|
| 24 (S. epidermidis ATCC 12228) | 3.65 ± 0.21 | 3.66 ± 0.08 | 4.01 ± 1.12 | 3.70 ± 0.71 | 3.75 ± 0.65 | 4.03 ± 0.95 |
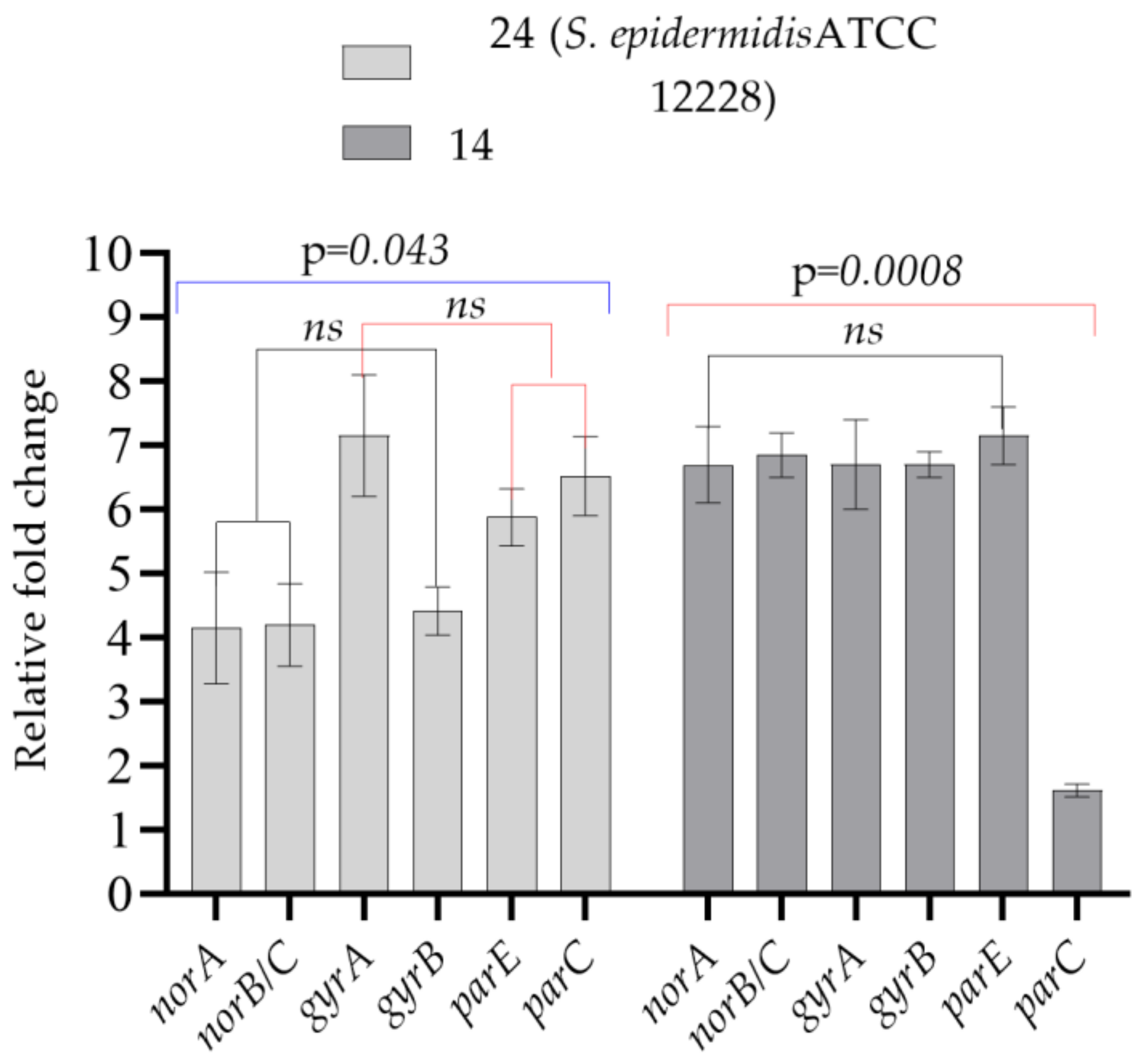
| Test Bacteria | norA | norB/C | gyrA | gyrB | parE | parC |
|---|---|---|---|---|---|---|
| 24 (S. epidermidis ATCC 12228) | 4.14 ± 1.23 | 4.20 ± 0.91 | 7.15 ± 1.34 | 4.42 ± 0.53 | 5.88 ± 0.63 | 6.52 ± 0.88 |
| 14 | 6.70 ± 0.85 | 6.85 ± 0.49 | 6.70 ± 0.99 | 6.70 ± 0.28 | 7.15 ± 0.64 | 1.61 ± 0.14 |
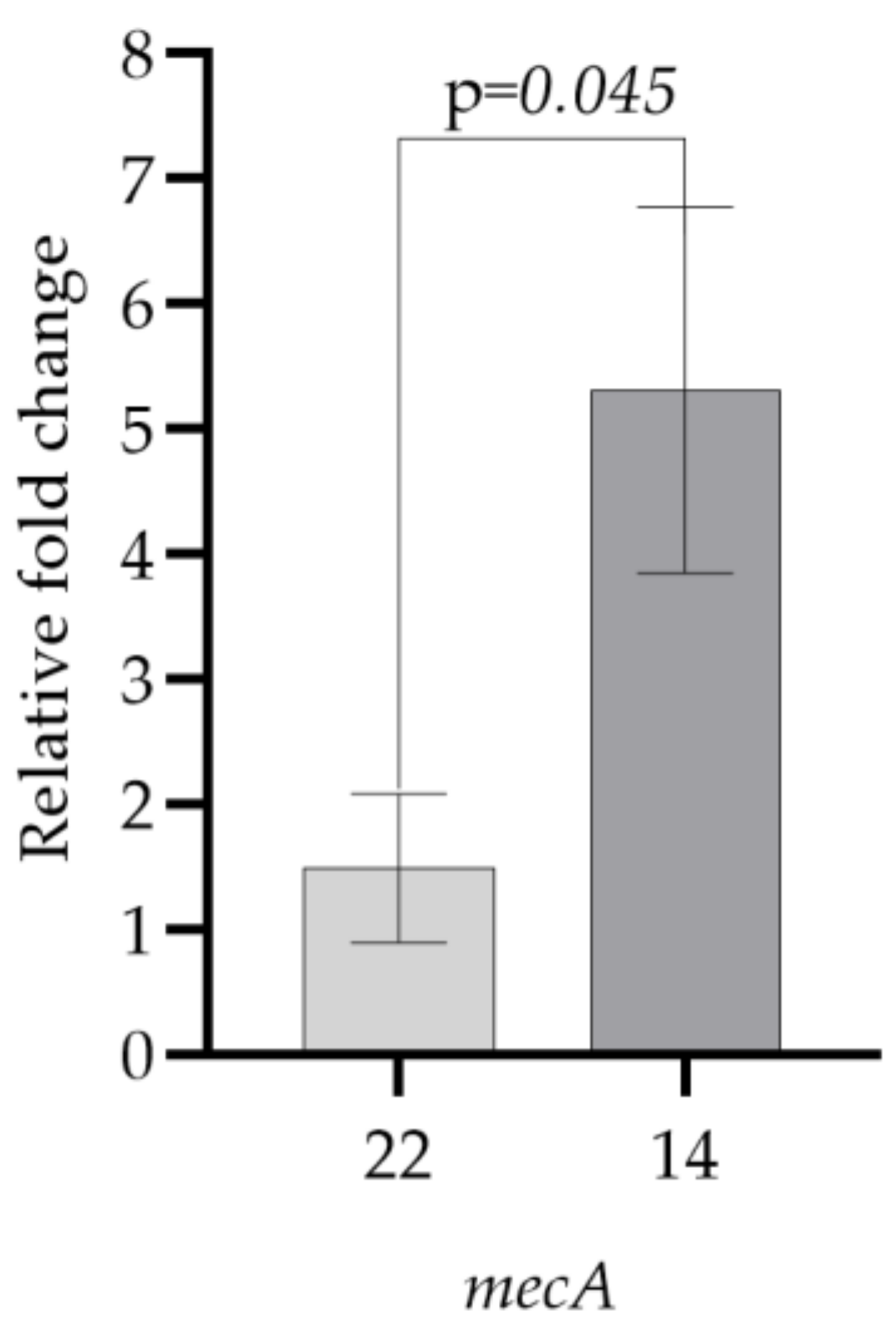
| Test Bacteria | mecA |
|---|---|
| 22 | 1.49 ± 0.59 |
| 14 | 5.32 ± 1.46 |
| Test Bacteria | mecA |
|---|---|
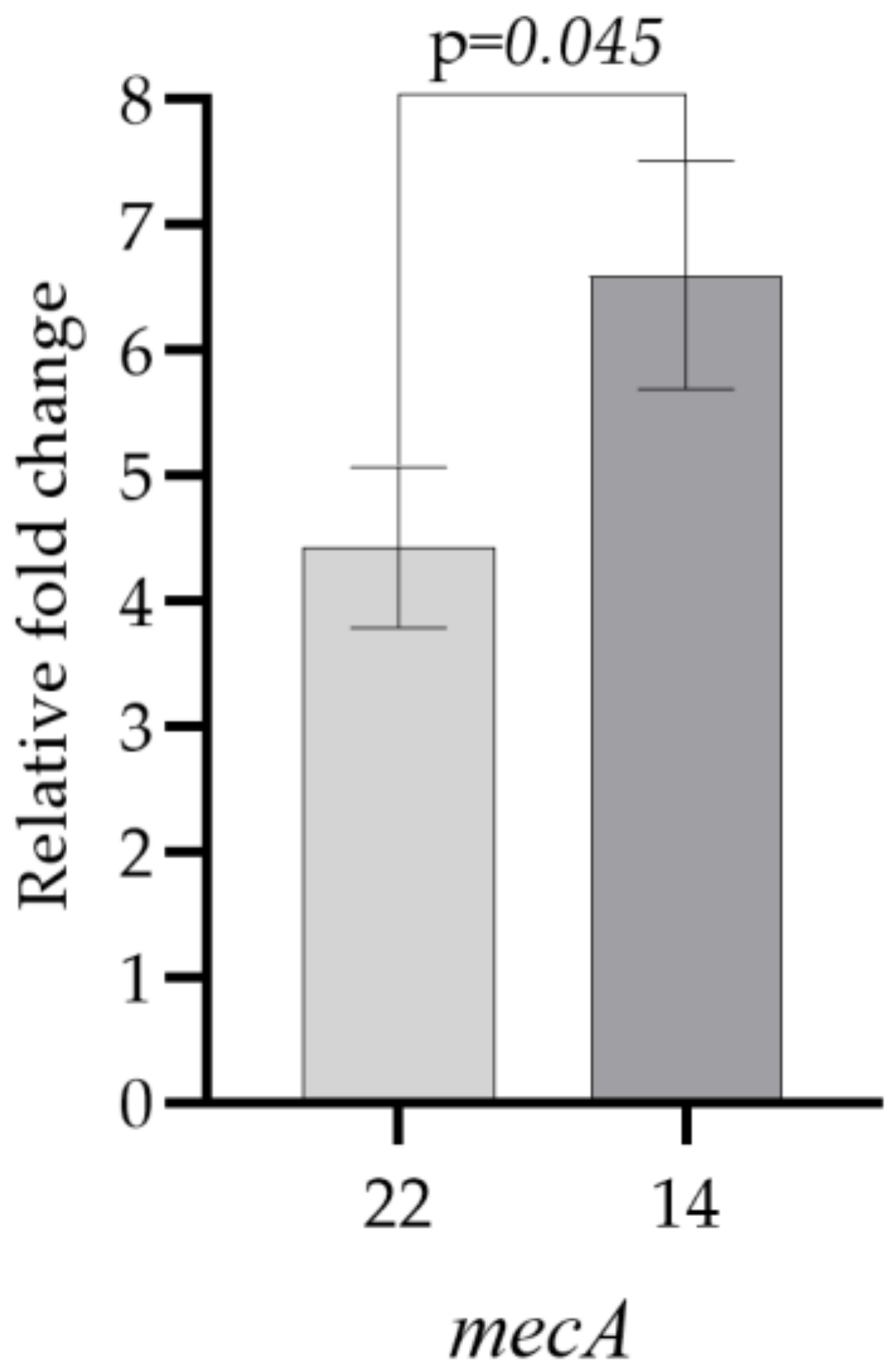
| 22 | 4.43 ± 0.91 |
| 14 | 6.60 ± 0.91 |
| Test Bacteria | Test Antibiotic | Pre-Exposure | Post-Exposure (30th Day) |
|---|---|---|---|
| 1 | CIP | 0.125 | 4 |
| 2 | OXA | 2 | 16 |
| 3 | OXA | 16 | >64 |
| 4 | OXA | 16 | >64 |
| 5 | OXA | 0.25 | 32 |
| 6 | OXA | 8 | 32 |
| 7 | OXA | 0.25 | 16 |
| 8 | OXA | 0.25 | 1 |
| 11 | OXA | 1 | 16 |
| CIP | 0.25 | 1 | |
| 12 | OXA | 8 | 32 |
| 13 | CIP | 0.25 | 8 |
| 14 | OXA | 64 | >64 |
| 16 | CIP | 0.125 | 0.5 |
| 17 | OXA | 0.5 | 16 |
| 18 | OXA | 0.25 | 64 |
| CIP | 0.25 | 8 | |
| 19 | OXA | 0.25 | 1 |
| 20 | OXA | 0.25 | 1 |
| CIP | <0.03125 | 0.125 | |
| 21 | OXA | 0.25 | 64 |
| CIP | 0.125 | 4 | |
| 22 | OXA | 2 | 4 |
| CIP | 0.25 | 4 | |
| 23 | OXA | 8 | 64 |
| 24 | OXA | 0.125 | 0.5 |
| CIP | 0.125 | 0.5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aras, A.; Rizvanoglu, S.S.; Tanriverdi, E.S.; Karaca, B.; Eryilmaz, M. The Effects of Antiperspirant Aluminum Chlorohydrate on the Development of Antibiotic Resistance in Staphylococcus epidermidis. Microorganisms 2023, 11, 948. https://doi.org/10.3390/microorganisms11040948
Aras A, Rizvanoglu SS, Tanriverdi ES, Karaca B, Eryilmaz M. The Effects of Antiperspirant Aluminum Chlorohydrate on the Development of Antibiotic Resistance in Staphylococcus epidermidis. Microorganisms. 2023; 11(4):948. https://doi.org/10.3390/microorganisms11040948
Chicago/Turabian StyleAras, Ayse, Suna Sibel Rizvanoglu, Elif Seren Tanriverdi, Basar Karaca, and Mujde Eryilmaz. 2023. "The Effects of Antiperspirant Aluminum Chlorohydrate on the Development of Antibiotic Resistance in Staphylococcus epidermidis" Microorganisms 11, no. 4: 948. https://doi.org/10.3390/microorganisms11040948