
Predatory Strategies of Myxococcus xanthus: Prey Susceptibility to OMVs and Moonlighting Enzymes

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains and Growth Conditions
2.2. OMV Production and Characterisation
2.3. Size Exclusion Chromatography
2.4. OMV Fusion, OMV Killing and Cellular Predation Assays
2.5. Cloning of M. xanthus gapA and pgk Genes
2.6. Expression and Purification of 6His-GAPDH and 6His-PGK
2.7. Enzymatic Assays of GAPDH Activity
2.8. Assays of Prey Cell Lysis and Prey Growth/Inhibition
3. Results
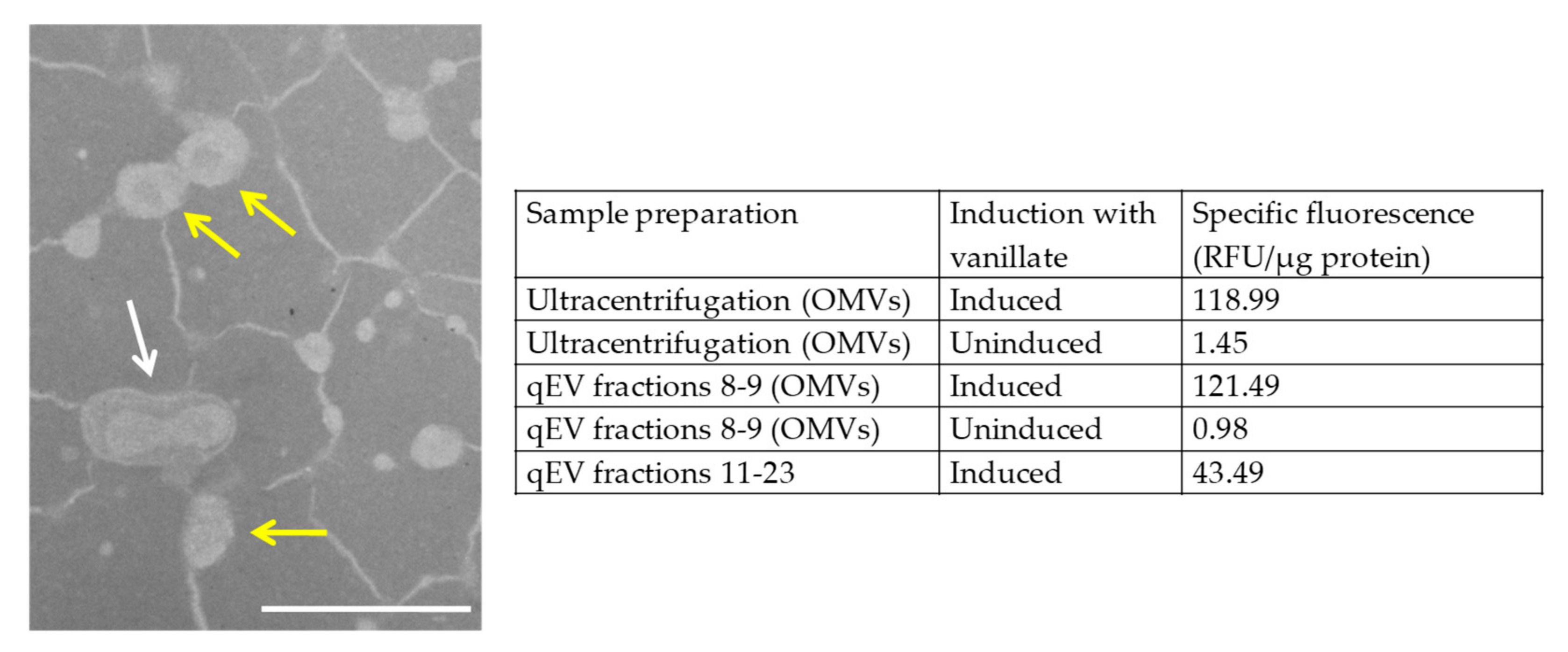
3.1. Inducing Expression of mCherry in M. xanthus Produces Fluorescent OMVs
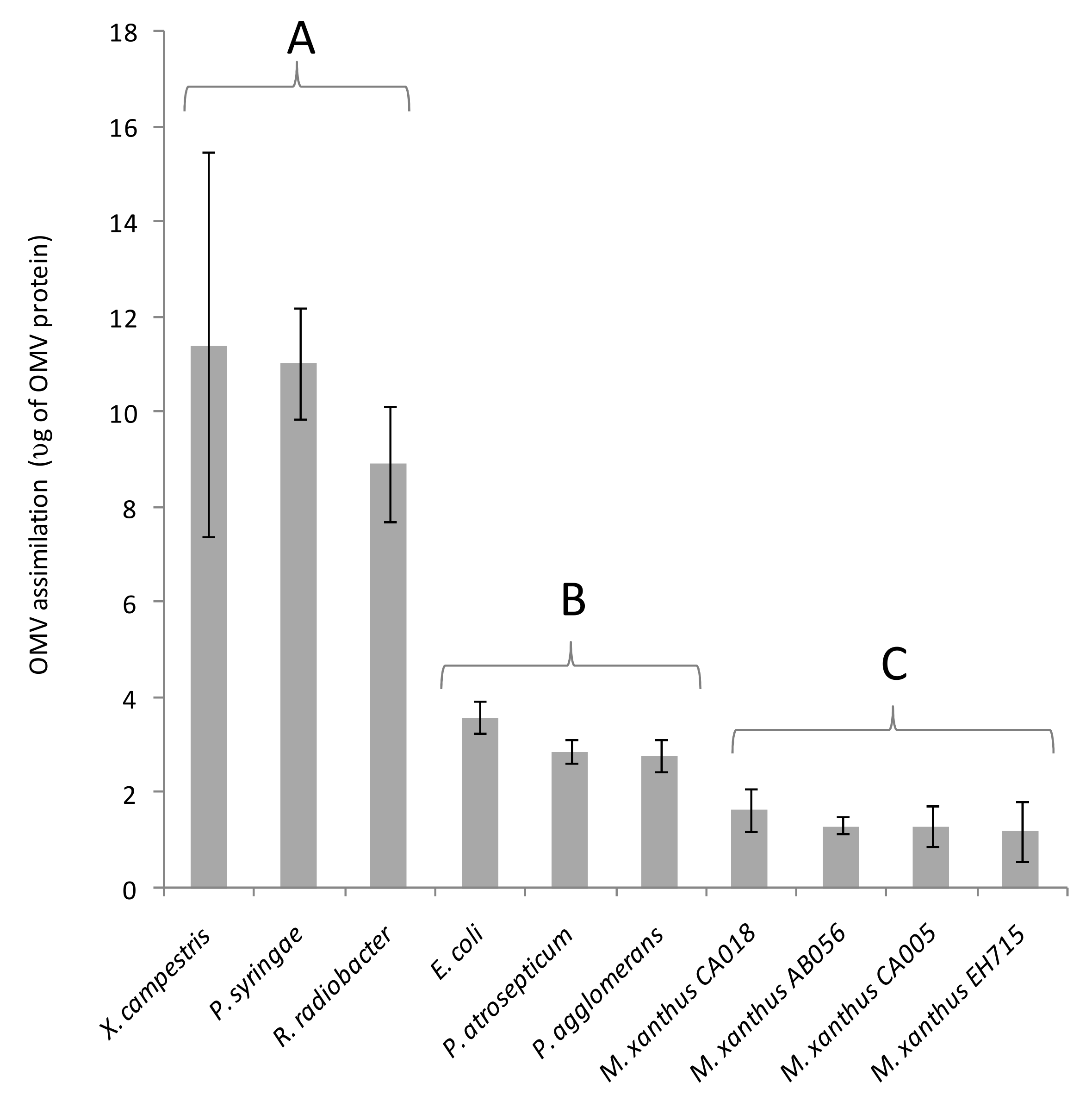
3.2. Fluorescent OMVs Allow Quantification of Cargo Protein Delivery to Recipient Cells
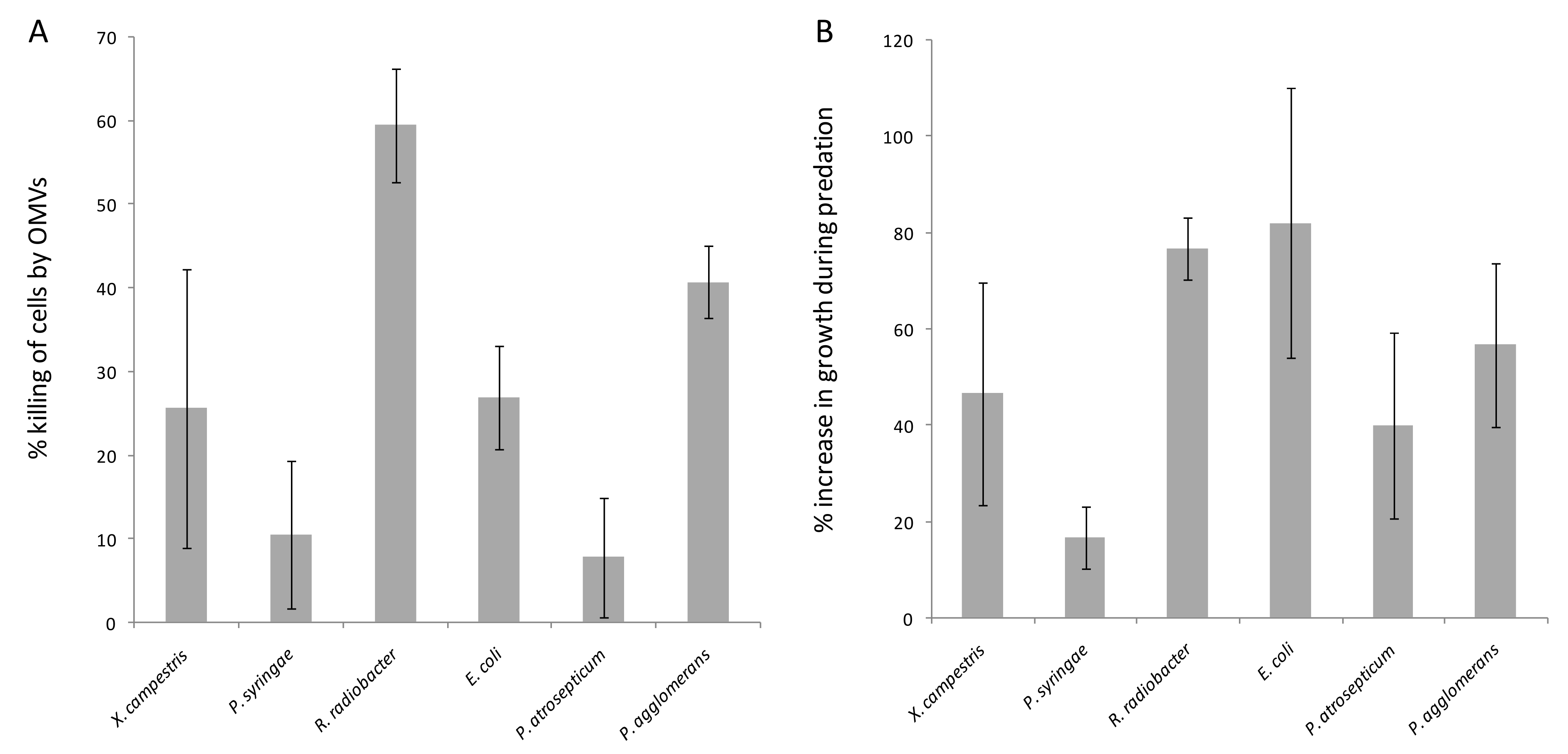
3.3. Cargo Protein Uptake Does Not Correlate with OMV Predatory Activity
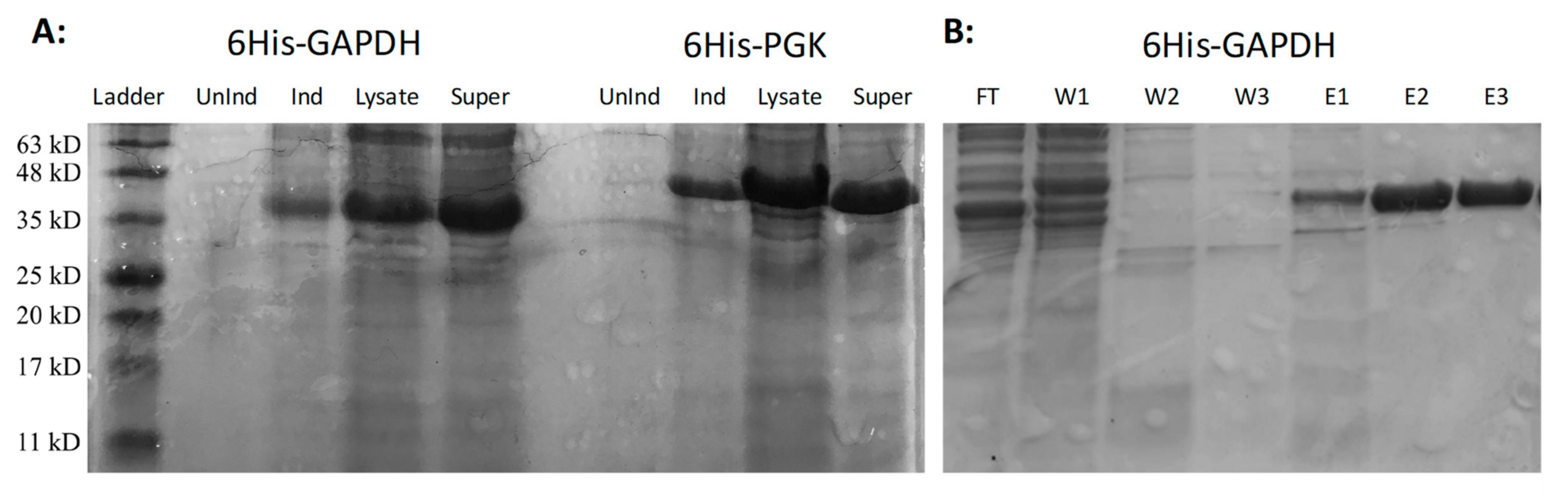
3.4. Expression and Purification of Active M. xanthus 6His-GAPDH
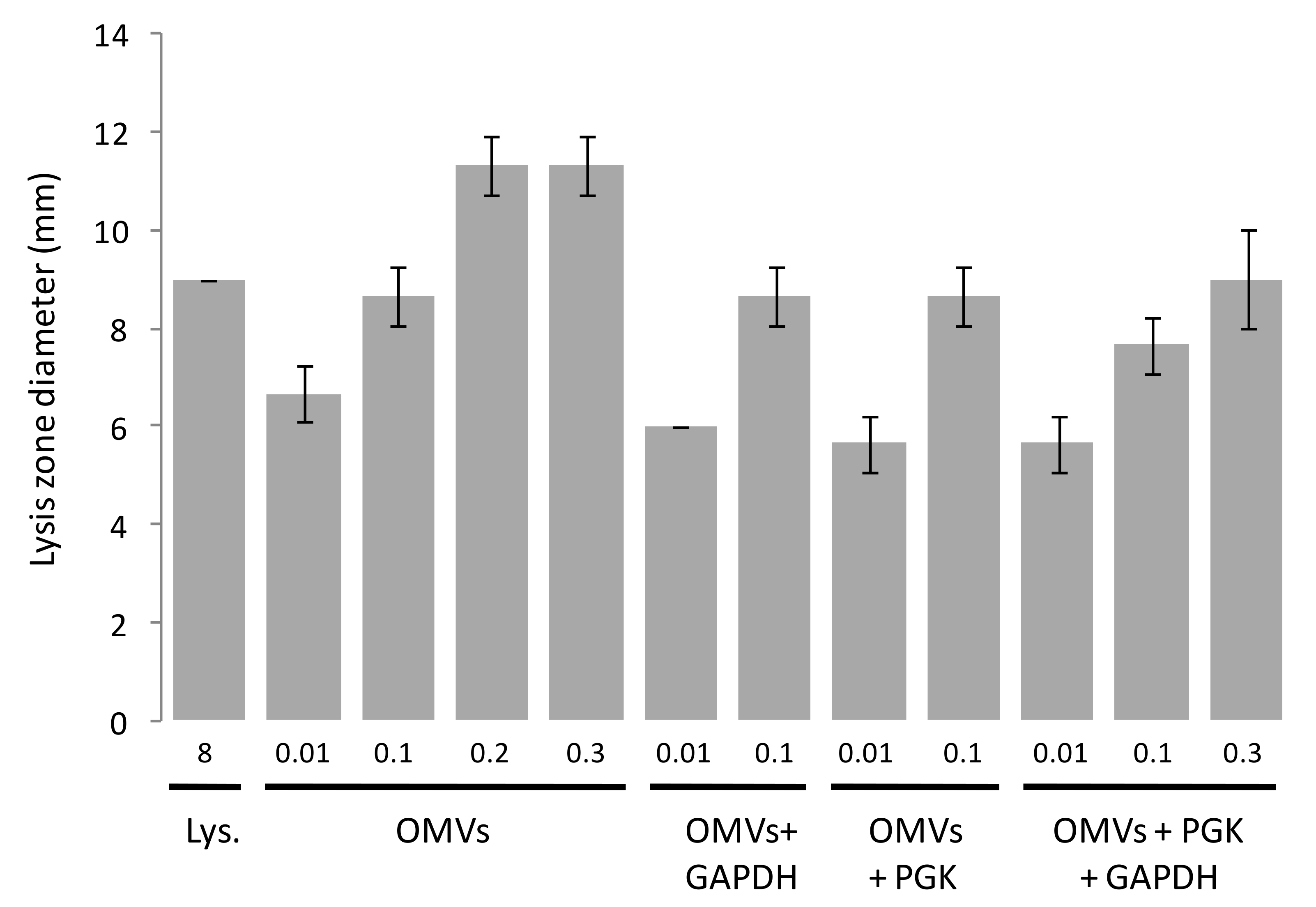
3.5. OMV-Dependent Lysis of Prey Is Not Stimulated by 6His-GAPDH in a Solid-Medium Based Assay
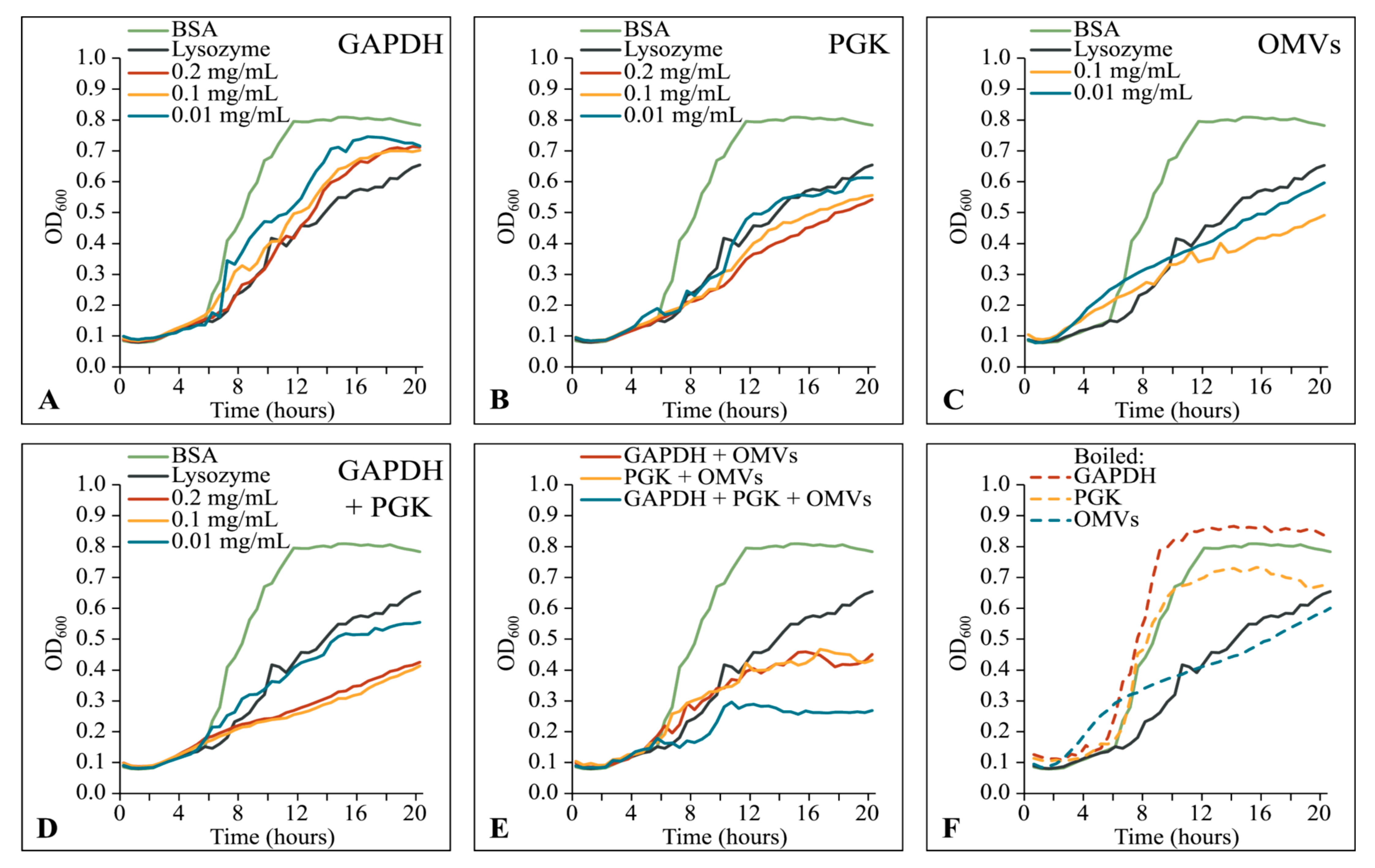
3.6. Native M. xanthus 6His-GAPDH and 6His-PGK Inhibit Prey Growth Independently of OMVs
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pérez, J.; Moraleda-Muñoz, A.; Marcos-Torres, F.J.; Muñoz-Dorado, J. Bacterial predation: 75 years and counting! Environ. Microbiol. 2016, 18, 766–779. [Google Scholar] [CrossRef] [PubMed]
- Furness, E.; Whitworth, D.E.; Zwarycz, A. Predatory interactions between myxobacteria and their prey. In The Ecology of Predation at the Microscale; Jurkevitch, E., Mitchell, R.J., Eds.; Springer: Cham, Switzerland, 2020; pp. 1–36. [Google Scholar]
- Morgan, A.D.; MacLean, R.C.; Hillesland, K.L.; Velicer, G.J. Comparative analysis of Myxococcus predation on soil Bacteria. Appl. Environ. Microbiol. 2010, 76, 6920–6927. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livingstone, P.G.; Morphew, R.M.; Whitworth, D.E. Myxobacteria are able to prey broadly upon clinically-relevant pathogens, exhibiting a prey range which cannot be explained by phylogeny. Front. Microbiol. 2017, 8, 1593. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livingstone, P.G.; Morphew, R.M.; Cookson, A.R.; Whitworth, D.E. Genome analysis, metabolic potential, and predatory capabilities of Herpetosiphon llansteffanense sp. nov. Appl. Environ. Microbiol. 2018, 84, e01040. [Google Scholar] [CrossRef] [Green Version]
- Marshall, R.C.; Whitworth, D.E. Is “wolf-pack” predation by antimicrobial bacteria cooperative? Cell behaviour and predatory mechanisms indicate profound selfishness, even when working alongside Kin. Bioessays 2019, 41, 1800247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thiery, S.; Kaimer, C. The Predation Strategy of Myxococcus xanthus. Front. Microbiol. 2020, 11, 2. [Google Scholar] [CrossRef] [Green Version]
- Berleman, J.E.; Scott, J.; Chumley, T.; Kirby, J.R. Predataxis behavior in Myxococcus xanthus. Proc. Natl. Acad. Sci. USA 2008, 105, 17127–17132. [Google Scholar] [CrossRef] [Green Version]
- Lloyd, D.G.; Whitworth, D.E. The myxobacterium Myxococcus xanthus can sense and respond to the quorum signals secreted by potential prey organisms. Front. Microbiol. 2017, 8, 439. [Google Scholar] [CrossRef]
- Seef, S.; Herrou, J.; de Boissier, P.; My, L.; Brasseur, G.; Robert, D.; Jain, R.; Mercier, R.; Cascales, E.; Habermann, B.H.; et al. A Tad-like apparatus is required for contact-dependent prey killing in predatory social bacteria. Elife 2021, 10, e72409. [Google Scholar] [CrossRef]
- Thiery, S.; Turowski, P.; Berleman, J.E.; Kaimer, C. The predatory soil bacterium Myxococcus xanthus combines a Tad- and an atypical type 3-like protein secretion system to kill bacterial cells. Cell Rep. 2022, 40, 111340. [Google Scholar] [CrossRef]
- Evans, A.G.; Davey, H.M.; Cookson, A.; Currinn, H.; Cooke-Fox, G.; Stanczyk, P.J.; Whitworth, D.E. Predatory activity of Myxococcus xanthus outer-membrane vesicles and properties of their hydrolase cargo. Microbiology 2012, 158, 2742–2752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livingstone, P.G.; Millard, A.D.; Swain, M.T.; Whitworth, D.E. Transcriptional changes when Myxococcus xanthus preys on Escherichia coli suggest myxobacterial predators are constitutively toxic but regulate their feeding. Microb. Genom. 2018, 4, 152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toyofuku, M.; Schild, S.; Kaparakis-Liaskos, M.; Eberl, L. Composition and functions of bacterial membrane vesicles. Nat. Rev. Microbiol. 2023. [Google Scholar] [CrossRef]
- Kahnt, J.; Aguiluz, K.; Koch, J.; Treuner-Lange, A.; Konovalova, A.; Huntley, S.; Hoppert, M.; Søgaard-Andersen, L.; Hedderich, R. Profiling the outer membrane proteome during growth and development of the social bacterium Myxococcus xanthus by selective biotinylation and analyses of outer membrane vesicles. J. Proteome Res. 2010, 9, 5197–5208. [Google Scholar] [CrossRef]
- Berleman, J.E.; Allen, S.; Danielewicz, M.A.; Remis, J.P.; Gorur, A.; Cunha, J.; Hadi, M.Z.; Zusman, D.R.; Northen, T.R.; Witkowska, H.E.; et al. The lethal cargo of Myxococcus xanthus outer membrane vesicles. Front. Microbiol. 2014, 5, 474. [Google Scholar] [CrossRef] [PubMed]
- Whitworth, D.E.; Slade, S.E.; Mironas, A. Composition of distinct sub-proteomes in Myxococcus xanthus: Metabolic cost and amino acid availability. Amino Acids 2015, 47, 2521–2531. [Google Scholar] [CrossRef] [PubMed]
- Zwarycz, A.S.; Livingstone, P.G.; Whitworth, D.E. Within-species variation in OMV cargo proteins: The Myxococcus xanthus OMV pan-proteome. Mol. Omics 2020, 16, 387–397. [Google Scholar] [CrossRef]
- Whitworth, D.E. Myxobacterial vesicles death at a distance? Adv. Appl. Microbiol. 2011, 75, 1–31. [Google Scholar]
- Kadurugamuwa, J.L.; Beveridge, T.J. Bacteriolytic effect of membrane vesicles from Pseudomonas aeruginosa on other bacteria including pathogens: Conceptually new antibiotics. J. Bacteriol. 1996, 178, 2767–2774. [Google Scholar] [CrossRef] [Green Version]
- Kadurugamuwa, J.L.; Beveridge, T.J. Membrane vesicles derived from Pseudomonas aeruginosa and Shigella flexneri can be integrated into the surfaces of other gram-negative bacteria. Microbiology 1999, 145, 2051–2060. [Google Scholar] [CrossRef] [Green Version]
- Schulz, E.; Goes, A.; Garcia, R.; Panter, F.; Koch, M.; Müller, R.; Fuhrmann, K.; Fuhrmann, G. Biocompatible bacteria-derived vesicles show inherent antimicrobial activity. J. Control. Release 2018, 290, 46–55. [Google Scholar] [CrossRef] [PubMed]
- Goes, A.; Lapuhs, P.; Kuhn, T.; Schulz, E.; Richter, R.; Panter, F.; Dahlem, C.; Koch, M.; Garcia, R.; Kiemer, A.K.; et al. Myxobacteria-Derived Outer Membrane Vesicles: Potential Applicability Against Intracellular Infections. Cells 2020, 9, 194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livingstone, P.G.; Morphew, R.M.; Whitworth, D.E. Genome sequencing and pan-genome analysis of 23 Corallococcus spp. strains reveal unexpected diversity, with particular plasticity of predatory gene sets. Front. Microbiol. 2018, 9, 3187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chambers, J.; Sparks, N.; Sydney, N.; Livingstone, P.G.; Cookson, A.R.; Whitworth, D.E. Comparative Genomics and Pan-Genomics of the Myxococcaceae, including a Description of Five Novel Species: Myxococcus eversor sp. nov., Myxococcus llanfairpwllgwyngyllgogerychwyrndrobwllllantysiliogo-gogochensis sp. nov., Myxococcus vastator sp. nov., Pyxidicoccus caerfyrddinensis sp. nov., and Pyxidicoccus trucidator sp. nov. Genome Biol. Evol. 2020, 12, 2289–2302. [Google Scholar]
- Sutton, D.; Livingstone, P.G.; Furness, E.; Swain, M.T.; Whitworth, D.E. Genome-Wide Identification of Myxobacterial Predation Genes and Demonstration of Formaldehyde Secretion as a Potentially Predation-Resistant Trait of Pseudomonas aeruginosa. Front. Microbiol. 2019, 10, 2650. [Google Scholar] [CrossRef]
- Whitworth, D.E.; Sydney, N.; Radford, E.J. Myxobacterial Genomics and Post-Genomics: A Review of Genome Biology, Genome Sequences and Related ‘Omics Studies. Microorganisms 2021, 9, 2143. [Google Scholar] [CrossRef]
- Sydney, N.; Swain, M.T.; So, J.M.T.; Hoiczyk, E.; Tucker, N.P.; Whitworth, D.E. The Genetics of Prey Susceptibility to Myxobacterial Predation: A Review, Including an Investigation into Pseudomonas aeruginosa Mutations Affecting Predation by Myxococcus xanthus. Microb. Physiol. 2021, 31, 57–66. [Google Scholar] [CrossRef]
- Kotrbová, A.; Štěpka, K.; Maška, M.; Pálenik, J.J.; Ilkovics, L.; Klemová, D.; Kravec, M.; Hubatka, F.; Dave, Z.; Hampl, A.; et al. TEM ExosomeAnalyzer: A computer-assisted software tool for quantitative evaluation of extracellular vesicles in transmission electron microscopy images. J. Extracell. Vesicles 2019, 8, 1560808. [Google Scholar] [CrossRef] [Green Version]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Sambrook, J.; Russell, D.W. Preparation and Transformation of Competent E. coli Using Calcium Chloride. CSH Protoc. 2006, 2006, pdb.prot3932. [Google Scholar]
- Zhang, J.; Snyder, S.H. Nitric oxide stimulates auto-ADP-ribosylation of glyceraldehyde-3-phosphate dehydrogenase. Proc. Natl. Acad. Sci. USA 1992, 89, 9382–9385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Z.; Clarke, A.J.; Beveridge, T.J. Gram-negative bacteria produce membrane vesicles which are capable of killing other bacteria. J. Bacteriol. 1998, 180, 5478–5483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lynch, J.B.; Alegado, R.A. Spheres of Hope, Packets of Doom: The Good and Bad of Outer Membrane Vesicles in Interspecies and Ecological Dynamics. J. Bacteriol. 2017, 199, e00012-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toyofuku, M.; Morinaga, K.; Hashimoto, Y.; Uhl, J.; Shimamura, H.; Inaba, H.; Schmitt-Kopplin, P.; Eberl, L.; Nomura, N. Membrane vesicle-mediated bacterial communication. ISME J. 2017, 11, 1504–1509. [Google Scholar] [CrossRef] [Green Version]
- Kadurugamuwa, J.L.; Beveridge, T.J. Delivery of the non-membrane-permeative antibiotic gentamicin into mammalian cells by using Shigella flexneri membrane vesicles. Antimicrob. Agents Chemother. 1998, 42, 1476–1483. [Google Scholar] [CrossRef] [Green Version]
- O’Donoghue, E.J.; Krachler, A.M. Mechanisms of outer membrane vesicle entry into host cells. Cell. Microbiol. 2016, 18, 1508–1517. [Google Scholar] [CrossRef]
- Xiao, Y.; Wei, X.; Ebright, R.; Wall, D. Antibiotic production by myxobacteria plays a role in predation. J. Bacteriol. 2011, 193, 4626–4633. [Google Scholar] [CrossRef] [Green Version]
- Schäberle, T.F.; Lohr, F.; Schmitz, A.; König, G.M. Antibiotics from myxobacteria. Nat. Prod. Rep. 2014, 31, 953–972. [Google Scholar] [CrossRef]
- Kumar, S.; Yadav, A.K.; Chambel, P.; Kaur, R. Molecular and functional characterization of myxobacteria isolated from soil in India. 3 Biotech. 2017, 7, 112. [Google Scholar] [CrossRef] [Green Version]
- Exner, M.; Bhattacharya, S.; Christiansen, B.; Gebel, J.; Goroncy-Bermes, P.; Hartemann, P.; Heeg, P.; Ilschner, C.; Kramer, A.; Larson, E.; et al. Antibiotic resistance: What is so special about multidrug-resistant Gram-negative bacteria? GMS Hyg. Infect. Control 2017, 12, Doc05. [Google Scholar]
- Brown, L.; Wolf, J.M.; Prados-Rosales, R.; Casadevall, A. Through the wall: Extracellular vesicles in Gram-positive bacteria, mycobacteria and fungi. Nat. Rev. Microbiol. 2015, 13, 620–630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pérez, J.; Muñoz-Dorado, J.; Braña, A.F.; Shimkets, L.J.; Sevillano, L.; Santamaría, R.I. Myxococcus xanthus induces actinorhodin overproduction and aerial mycelium formation by Streptomyces coelicolor. Microb. Biotechnol. 2011, 4, 175–183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Müller, S.; Strack, S.N.; Hoefler, B.C.; Straight, P.D.; Kearns, D.B.; Kirby, J.R. Bacillaene and sporulation protect Bacillus subtilis from predation by Myxococcus xanthus. Appl. Environ. Microbiol. 2014, 80, 5603–5610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pérez, J.; Jiménez-Zurdo, J.I.; Martínez-Abarca, F.; Millán, V.; Shimkets, L.J.; Muñoz-Dorado, J. Rhizobial galactoglucan determines the predatory pattern of Myxococcus xanthus and protects Sinorhizobium meliloti from predation. Environ. Microbiol. 2014, 16, 2341–2350. [Google Scholar] [CrossRef] [Green Version]
- Müller, S.; Strack, S.N.; Ryan, S.E.; Kearns, D.B.; Kirby, J.R. Predation by Myxococcus xanthus induces Bacillus subtilis to form spore-filled megastructures. Appl. Environ. Microbiol. 2015, 81, 203–210. [Google Scholar] [CrossRef] [Green Version]
- Wei, X.; Vassallo, C.N.; Pathak, D.T.; Wall, D. Myxobacteria produce outer membrane-enclosed tubes in unstructured environments. J. Bacteriol. 2014, 196, 1807–1814. [Google Scholar] [CrossRef] [Green Version]
- Kulkarni, H.M.; Jagannadham, M.V. Biogenesis and multifaceted roles of outer membrane vesicles from Gram-negative bacteria. Microbiology 2014, 160, 2109–2121. [Google Scholar] [CrossRef] [Green Version]
- Berleman, J.E.; Chumley, T.; Cheung, P.; Kirby, J.R. Rippling is a predatory behavior in Myxococcus xanthus. J. Bacteriol. 2006, 188, 5888–5895. [Google Scholar] [CrossRef] [Green Version]
- Vassallo, C.; Pathak, D.T.; Cao, P.; Zuckerman, D.M.; Hoiczyk, E.; Wall, D. Cell rejuvenation and social behaviors promoted by LPS exchange in myxobacteria. Proc. Natl. Acad. Sci. USA 2015, 112, E2939–E2946. [Google Scholar] [CrossRef] [Green Version]
- McMahon, H.T.; Boucrot, E. Membrane curvature at a glance. J. Cell Sci. 2015, 128, 1065–1070. [Google Scholar] [CrossRef] [Green Version]
- Garcia, R.; Pistorius, D.; Stadler, M.; Müller, R. Fatty acid-related phylogeny of myxobacteria as an approach to discover polyunsaturated omega-3/6 Fatty acids. J. Bacteriol. 2011, 193, 1930–1942. [Google Scholar] [CrossRef] [Green Version]
- Ahrendt, T.; Wolff, H.; Bode, H.B. Neutral and Phospholipids of the Myxococcus xanthus Lipodome during Fruiting Body Formation and Germination. Appl. Environ. Microbiol. 2015, 81, 6538–6547. [Google Scholar] [CrossRef] [Green Version]
- Livingstone, P.G.; Ingleby, O.; Girdwood, S.; Cookson, A.R.; Morphew, R.M.; Whitworth, D.E. Predatory Organisms with Untapped Biosynthetic Potential: Descriptions of Novel Corallococcus Species C. aberystwythensis sp. nov., C. carmarthensis sp. nov., C. exercitus sp. nov., C. interemptor sp. nov., C. llansteffanensis sp. nov., C. praedator sp. nov., C. sicarius sp. nov., and C. terminator sp. nov. Appl. Environ. Microbiol. 2020, 86, e01931-19. [Google Scholar]
- Singh, N.; Bhalla, N. Moonlighting Proteins. Annu. Rev. Genet. 2020, 54, 265–285. [Google Scholar] [CrossRef] [PubMed]
- Glaser, P.E.; Gross, R.W. Rapid plasmenylethanolamine-selective fusion of membrane bilayers catalyzed by an isoform of glyceraldehyde-3-phosphate dehydrogenase: Discrimination between glycolytic and fusogenic roles of individual isoforms. Biochemistry 1995, 34, 12193–12203. [Google Scholar] [CrossRef] [PubMed]
- Wagener, J.; Schneider, J.J.; Baxmann, S.; Kalbacher, H.; Borelli, C.; Nuding, S.; Küchler, R.; Wehkamp, J.; Kaeser, M.D.; Mailänder-Sanchez, D.; et al. A peptide derived from the highly conserved protein GAPDH is involved in tissue protection by different antifungal strategies and epithelial immunomodulation. J. Investig. Dermatol. 2013, 133, 144–153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henderson, B.; Martin, A.C. Protein moonlighting: A new factor in biology and medicine. Biochem. Soc. Trans. 2014, 42, 1671–1678. [Google Scholar] [CrossRef]
- Fu, Q.; Wei, Z.; Liu, X.; Xiao, P.; Lu, Z.; Chen, Y. Glyceraldehyde-3-phosphate dehydrogenase, an immunogenic Streptococcus equi ssp. zooepidemicus adhesion protein and protective antigen. J. Microbiol. Biotechnol. 2013, 23, 579–585. [Google Scholar] [CrossRef] [Green Version]
- Whitworth, D.E.; Morgan, B.H. Synergism Between Bacterial GAPDH and OMVs: Disparate Mechanisms but Co-Operative Action. Front. Microbiol. 2015, 6, 1231. [Google Scholar] [CrossRef] [Green Version]
- Kinoshita, H.; Ohuchi, S.; Arakawa, K.; Watanabe, M.; Kitazawa, H.; Saito, T. Isolation of lactic acid bacteria bound to the porcine intestinal mucosa and an analysis of their moonlighting adhesins. Biosci. Microbiota Food Health 2016, 35, 185–196. [Google Scholar] [CrossRef] [Green Version]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zwarycz, A.S.; Page, T.; Nikolova, G.; Radford, E.J.; Whitworth, D.E. Predatory Strategies of Myxococcus xanthus: Prey Susceptibility to OMVs and Moonlighting Enzymes. Microorganisms 2023, 11, 874. https://doi.org/10.3390/microorganisms11040874
Zwarycz AS, Page T, Nikolova G, Radford EJ, Whitworth DE. Predatory Strategies of Myxococcus xanthus: Prey Susceptibility to OMVs and Moonlighting Enzymes. Microorganisms. 2023; 11(4):874. https://doi.org/10.3390/microorganisms11040874
Chicago/Turabian StyleZwarycz, Allison S., Thomas Page, Gabriela Nikolova, Emily J. Radford, and David E. Whitworth. 2023. "Predatory Strategies of Myxococcus xanthus: Prey Susceptibility to OMVs and Moonlighting Enzymes" Microorganisms 11, no. 4: 874. https://doi.org/10.3390/microorganisms11040874