
Promising Probiotic Properties of the Yeasts Isolated from Rabilé, a Traditionally Fermented Beer Produced in Burkina Faso
 , ,
, ,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Microorganisms and Growth Conditions
2.2. Molecular Identification of Yeast Isolates
2.2.1. Amplification of the 5.8S-ITS Region and RFLP Analysis
2.2.2. Yeast Strain Identification by Sequencing
2.3. Growth Capacity at 37 °C
2.4. Survival of Simulated Gastrointestinal (GI) Digestion
2.4.1. Tolerance of Pepsin Presence and Acidic pH
2.4.2. Tolerance of Bile Salts
2.5. Auto-Aggregation Ability
2.6. Co-Aggregation Activity
2.7. Hydrophobicity
2.8. Hemolytic Activity
2.9. Antioxidant Activity
2.10. Antibacterial Activity
2.11. Statistical Analyses
3. Results
3.1. Molecular Identification of Yeast Strains
3.2. Tolerance at 37 °C (Body Temperature)
3.3. Survival of Simulated Gastrointestinal Conditions
3.3.1. Tolerance of Pepsin 0.3% and pH 2.5 at 37 °C
3.3.2. Bile Salt Tolerance
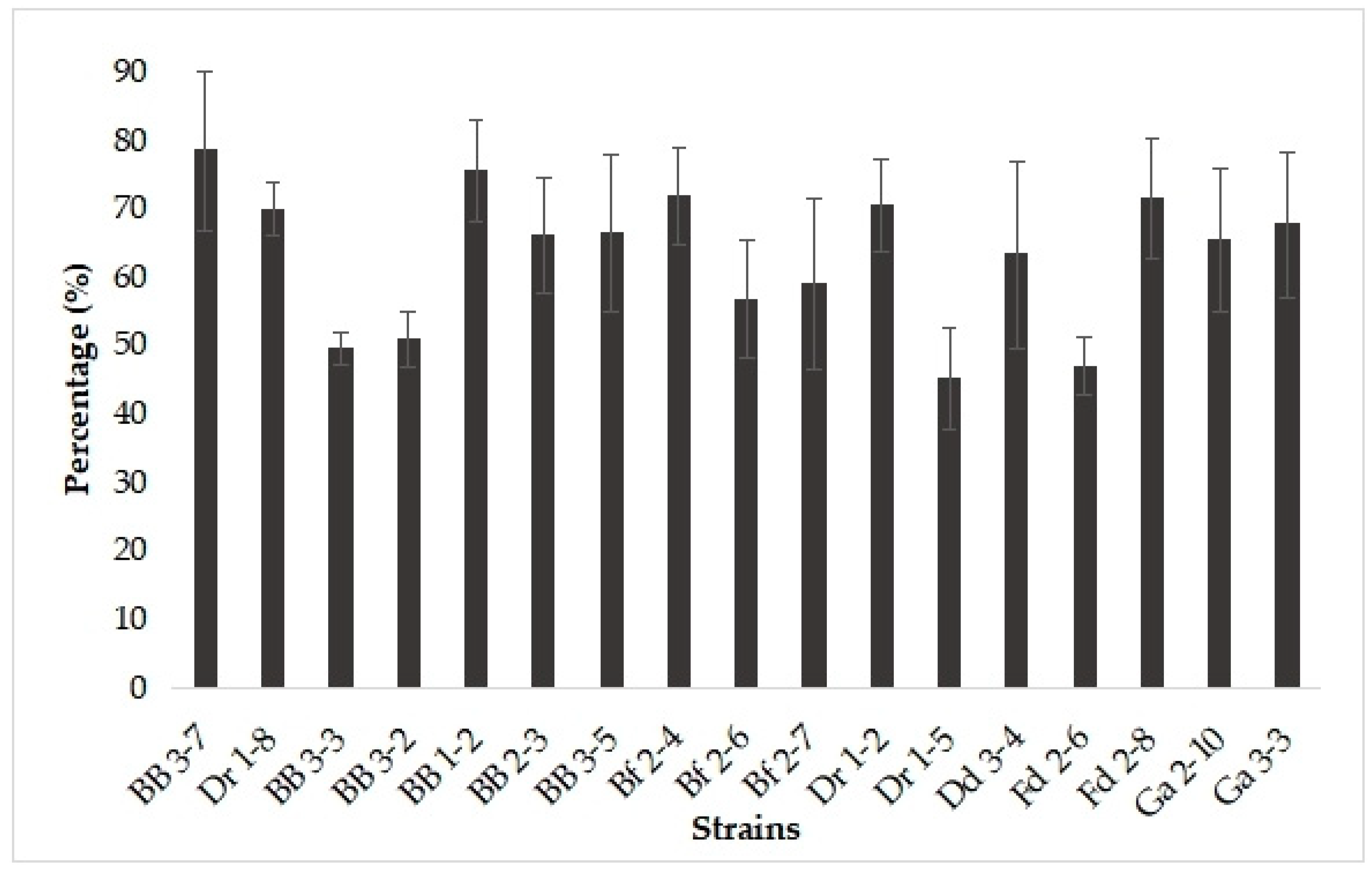
3.4. Auto-Aggregation, Co-Aggregation, and Hydrophobicity of Yeast Isolates
3.5. Hemolytic Activity
3.6. Antioxidant Activity
3.7. Antibacterial Activities of the Yeast Strains
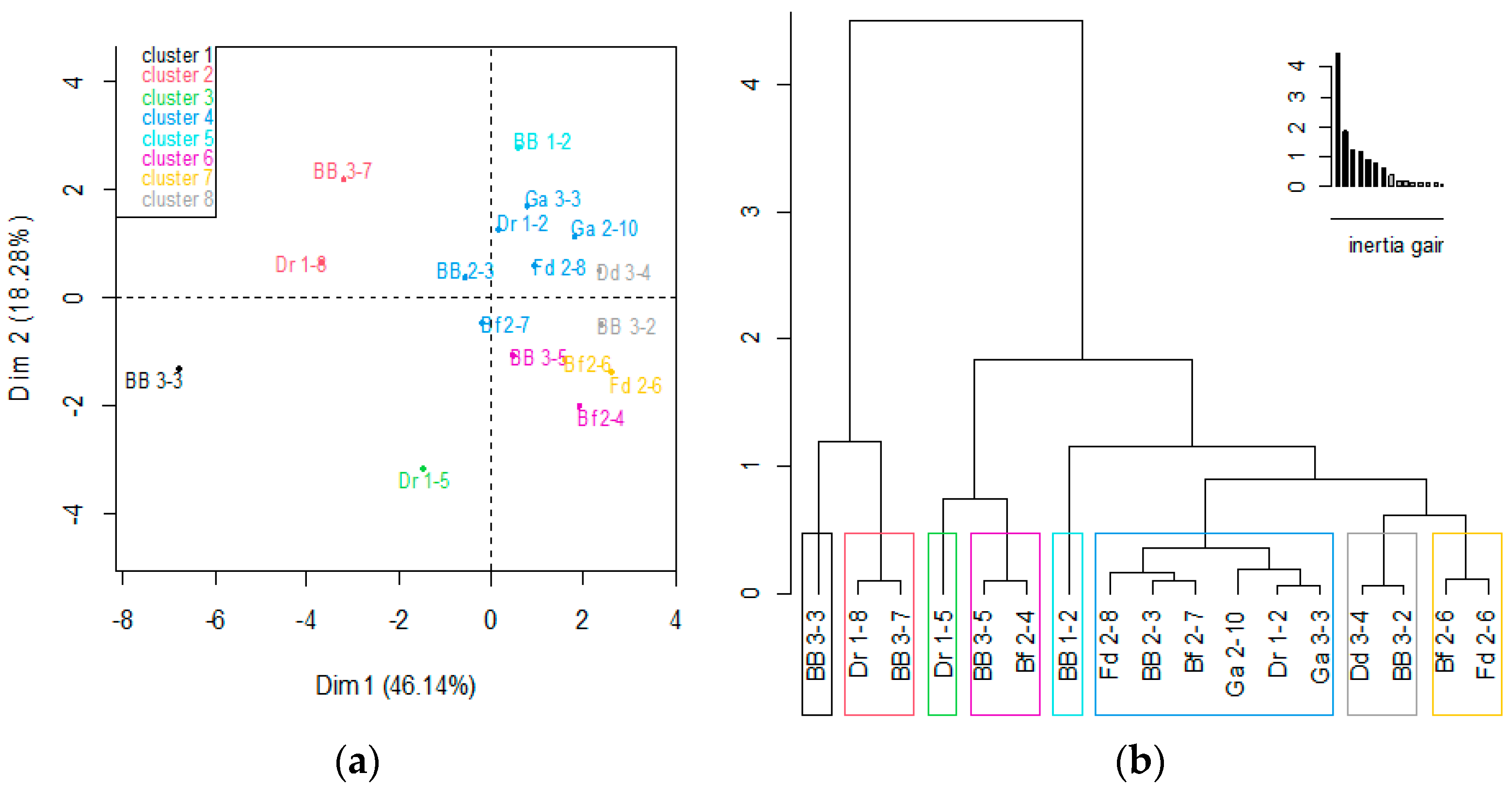
3.8. Selection of Yeasts with the Highest Probiotic Potential
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- FAO. The Future of Food and Agriculture—Alternative Pathways to 2050; FAO: Rome, Italy, 2018. [Google Scholar]
- Choi, K.R.; Yu, H.E.; Lee, S.Y. Microbial food: Microorganisms repurposed for our food. Microb. Biotechnol. 2022, 15, 18–25. [Google Scholar] [CrossRef]
- Setta, M.C.; Matemu, A.; Mbega, E.R. Potential of probiotics from fermented cereal-based beverages in improving health of poor people in Africa. J. Food Sci. Technol. 2020, 57, 3935–3946. [Google Scholar] [CrossRef] [PubMed]
- Franz, C.M.; Huch, M.; Mathara, J.M.; Abriouel, H.; Benomar, N.; Reid, G.; Galvez, A.; Holzapfel, W.H. African fermented foods and probiotics. Int. J. Food Microbiol. 2014, 190, 84–96. [Google Scholar] [CrossRef] [PubMed]
- Diaz, M.; Kellingray, L.; Akinyemi, N.; Adefiranye, O.O.; Olaonipekun, A.B.; Bayili, G.R.; Ibezim, J.; Du Plessis, A.S.; Houngbédji, M.; Kamya, D.; et al. Comparison of the microbial composition of African fermented foods using amplicon sequencing. Sci. Rep. 2019, 9, 13863. [Google Scholar] [CrossRef] [Green Version]
- Obafemi, Y.D.; Oranusi, S.U.; Ajanaku, K.O.; Akinduti, P.A.; Leech, J.; Cotter, P.D. African fermented foods: Overview, emerging benefits, and novel approaches to microbiome profiling. Npj Sci. Food 2022, 6, 15. [Google Scholar] [CrossRef]
- Kayitesi, E.; Onojakpor, O.; Moyo, S.M. Highlighting the impact of lactic-acid-bacteria-derived flavours or aromas on sensory perception of African fermented cereals. Fermentation 2023, 9, 111. [Google Scholar] [CrossRef]
- Mehlomakulu, N.N.; Moyo, S.M. Yeast derived metabolites and their impact on nutritional and bioactive properties of African fermented maize products. Food Biosci. 2023, 51, 102300. [Google Scholar] [CrossRef]
- FAO; WHO. Guidelines for the Evaluation of Probiotics in Food; Food and Agriculture Organization of the United Nations and World Health Organization Working Group Report; FAO: London, ON, Canada, 2002. [Google Scholar]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. Expert consensus document: The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef] [Green Version]
- Picard, C.; Fioramonti, J.; Francois, A.; Robinson, T.; Neant, F.; Matuchansky, C. Review article: Bifidobacteria as probiotic agents–physiological effects and clinical benefits. Aliment. Pharmacol. Ther. 2005, 22, 495–512. [Google Scholar] [CrossRef]
- Kumar, H.; Collado, M.C.; Wopereis, H.; Salminen, S.; Knol, J.; Roeselers, G. The Bifidogenic Effect Revisited—Ecology and Health Perspectives of Bifidobacterial Colonization in Early Life. Microorganisms 2020, 8, 1855. [Google Scholar] [CrossRef]
- Chen, J.; Chen, X.; Ho, C.L. Recent development of probiotic bifidobacteria for treating human diseases. Front. Bioeng. Biotechnol. 2021, 9, 770248. [Google Scholar] [CrossRef] [PubMed]
- Jeong, J.-J.; Park, H.J.; Cha, M.G.; Park, E.; Won, S.-M.; Ganesan, R.; Gupta, H.; Gebru, Y.A.; Sharma, S.P.; Lee, S.B.; et al. The Lactobacillus as a probiotic: Focusing on liver diseases. Microorganisms 2022, 10, 288. [Google Scholar] [CrossRef] [PubMed]
- Torres-Maravilla, E.; Reyes-Pavón, D.; Benítez-Cabello, A.; González-Vázquez, R.; Ramírez-Chamorro, L.M.; Langella, P.; Bermúdez-Humarán, L.G. Strategies for the identification and assessment of bacterial strains with specific probiotic traits. Microorganisms 2022, 10, 1389. [Google Scholar] [CrossRef]
- Chou, L.-S.; Weimer, B. Isolation and characterization of acid- and bile-tolerant isolates from strains of Lactobacillus acidophilus. J. Dairy Sci. 1999, 82, 23–31. [Google Scholar] [CrossRef] [Green Version]
- Csutak, O.; Sabău, E.; Pelinescu, D.; Corbu, V.; Cîrpici, I.; Vassu, T. Molecular identification and metabolic screening of some yeast strains from foods. AgroLife Sci. J. 2016, 5, 51–58. [Google Scholar]
- Johansen, P.G.; Owusu-Kwarteng, J.; Parkouda, C.; Padonou, S.W.; Jespersen, L. Occurrence and importance of yeasts in indigenous fermented food and beverages produced in Sub-Saharan Africa. Front. Microbiol. 2019, 10, 1789. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rai, A.K.; Pandey, A.; Sahoo, D. Biotechnological potential of yeasts in functional food industry. Trends Food Sci. Technol. 2019, 83, 129–137. [Google Scholar] [CrossRef]
- Tullio, V. Yeast genomics and its applications in biotechnological processes: What is our present and near future? J. Fungi 2022, 8, 752. [Google Scholar] [CrossRef]
- Zhimo, V.Y.; Biasi, A.; Kumar, A.; Feygenberg, O.; Salim, S.; Vero, S.; Wisniewski, M.; Droby, S. Yeasts and bacterial consortia from kefir grains are effective biocontrol agents of postharvest diseases of Fruits. Microorganisms 2020, 8, 428. [Google Scholar] [CrossRef] [Green Version]
- Barbulescu, I.D.; Dumitrache, C.; Diguta, C.F.; Matei, P.M.; Frincu, M.; Marculescu, S.I.; Ciric, A.I.; Tudor, V.; Boroiu, E.M.; Teodorescu, R. Evolution at the microfermenter level of the growth dynamics of Saccharomyces cerevisiae and Starmella bacillaris yeasts with potential for use in winemaking at the pietroasa winery. AgroLife Sci. J. 2022, 11, 9–16. [Google Scholar] [CrossRef]
- Chen, P.; Zhang, Q.; Dang, H.; Liu, X.; Tian, F.; Zhao, J.; Zhang, H.; Chen, W. Screening for potential new probiotic based on probiotic properties and α-glucosidase inhibitory activity. Food Control. 2014, 35, 65–72. [Google Scholar] [CrossRef]
- Abid, R.; Waseem, H.; Ali, J.; Ghazanfar, S.; Muhammad Ali, G.; Elasbali, A.M.; Alharethi, S.H. Probiotic yeast Saccharomyces: Back to nature to improve human health. J. Fungi 2022, 8, 444. [Google Scholar] [CrossRef] [PubMed]
- Vysoka, M.; Szotkowski, M.; Slaninova, E.; Dzuricka, L.; Strecanska, P.; Blazkova, J.; Marova, I. Oleaginous Yeast Extracts and Their Possible Effects on Human Health. Microorganisms 2023, 11, 492. [Google Scholar] [CrossRef] [PubMed]
- Kunyeit, L.; Rao, R.P.; Anu-Appaiah, K.A. Yeasts originating from fermented foods, their potential as probiotics and therapeutic implication for human health and disease. Crit. Rev. Food Sci. Nutr. 2023. [Google Scholar] [CrossRef]
- Staniszewski, A.; Kordowska-Wiater, M. Probiotic and potentially probiotic yeasts—Characteristics and food application. Foods 2021, 10, 1306. [Google Scholar] [CrossRef]
- Tamang, J.P.; Lama, S. Probiotic properties of yeasts in traditional fermented foods and beverages. J. Appl. Microbiol. 2022, 132, 3533–3542. [Google Scholar] [CrossRef]
- Gil-Rodríguez, A.M.; Carrascosa, A.V.; Requena, T. Yeasts in foods and beverages: In vitro characterisation of probiotic traits. LWT-Food Sci. Technol. 2015, 64, 1156–1162. [Google Scholar] [CrossRef] [Green Version]
- Pihurov, M.; Păcularu-Burada, B.; Cotârleţ, M.; Vasile, M.A.; Bahrim, G.E. Novel insights for metabiotics production by using artisanal probiotic cultures. Microorganisms 2021, 9, 2184. [Google Scholar] [CrossRef]
- Pais, P.; Almeida, V.; Yılmaz, M.; Teixeira, M.C. Saccharomyces boulardii: What makes it tick as successful probiotic? J. Fungi 2020, 6, 78. [Google Scholar] [CrossRef]
- Fu, J.; Liu, J.; Wen, X.; Zhang, G.; Cai, J.; Qiao, Z.; An, Z.; Zheng, J.; Li, L. Unique probiotic properties and bioactive metabolites of Saccharomyces boulardii. Probiotics Antimicrob. Proteins 2022, 1–16. [Google Scholar] [CrossRef]
- Diguță, C.F.; Mihai, C.; Toma, R.C.; Cîmpeanu, C.; Matei, F. In vitro assessment of yeasts strains with probiotic attributes for aquaculture use. Foods 2023, 12, 124. [Google Scholar] [CrossRef]
- Palma, M.L.; Zamith-Miranda, D.; Martins, F.S.; Bozza, F.A.; Nimrichter, L.; Montero-Lomeli, M.; Marques, E.T.A.; Douradinha, B. Probiotic Saccharomyces cerevisiae strains as biotherapeutic tools: Is there room for improvement? Appl. Microbiol. Biotechnol. 2015, 99, 6563–6570. [Google Scholar] [CrossRef] [PubMed]
- Motey, G.A.; Johansen, P.G.; Owusu-Kwarteng, J.; Ofori, L.A.; Obiri-Danso, K.; Siegumfeldt, H.; Larsen, N.; Jespersen, L. Probiotic potential of Saccharomyces cerevisiae and Kluyveromyces marxianus isolated from West African spontaneously fermented cereal and milk products. Yeast 2020, 37, 403–412. [Google Scholar] [CrossRef] [PubMed]
- Ogunremi, O.R.; Sanni, A.I.; Agrawal, R. Probiotic potentials of yeasts isolated from some cereal-based Nigerian traditional fermented food products. J. Appl. Microbiol. 2015, 119, 797–808. [Google Scholar] [CrossRef] [PubMed]
- Corbu, V.; Csutak, O. Biodiversity studies on Pichia kudriavzevii from Romanian spontaneous fermented products. AgroLife Sci. J. 2020, 9, 104–114. [Google Scholar]
- Corbu, V.M.; Csutak, O. Molecular and physiological diversity of indigenous yeasts isolated from spontaneously fermented wine wort from Ilfov county, Romania. Microorganisms 2023, 11, 37. [Google Scholar] [CrossRef]
- Adisa, A.M.; Ifesan, O.B.T.; Enujiugha, V.N.; Adepeju, A.B. Microbiological and probiotic assessment of yeast isolated from wholegrain millet sourdoughs. J. Adv. Microbiol. 2020, 20, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Shruthi, B.; Deepa, N.; Somashekaraiah, R.; Adithi, G.; Divyashree, S.; Sreenivasa, M.Y. Exploring biotechnological and functional characteristics of probiotic yeasts: A review. Biotechnol. Rep. 2022, 34, e00716. [Google Scholar] [CrossRef]
- De Melo Pereira, G.V.; de Oliveira Coelho, B.; Magalhães Júnior, A.I.; Thomaz-Soccol, V.; Soccol, C.R. How to select a probiotic? A review and update of methods and criteria. Biotechnol. Adv. 2018, 36, 2060–2076. [Google Scholar] [CrossRef]
- Alkalbani, N.S.; Osaili, T.M.; Al-Nabulsi, A.A.; Olaimat, A.N.; Liu, S.-Q.; Shah, N.P.; Apostolopoulos, V.; Ayyash, M.M. Assessment of yeasts as potential probiotics: A review of gastrointestinal tract conditions and investigation methods. J. Fungi 2022, 8, 365. [Google Scholar] [CrossRef]
- Fernández-Pacheco, P.; Pintado, C.; Briones Pérez, A.; Arévalo-Villena, M. Potential probiotic strains of Saccharomyces and non-Saccharomyces: Functional and Biotechnological Characteristics. J. Fungi 2021, 7, 177. [Google Scholar] [CrossRef]
- Elghandour, M.M.Y.; Tan, Z.L.; Abu Hafsa, S.H.; Adegbeye, M.J.; Greiner, R.; Ugbogu, E.A.; Cedillo Monroy, J.; Salem, A.Z.M. Saccharomyces cerevisiae as a probiotic feed additive to non and pseudo-ruminant feeding: A review. J. Appl. Microbiol. 2019, 28, 658–674. [Google Scholar]
- Pang, Y.; Zhang, H.; Wen, H.; Wan, H.; Wu, H.; Chen, Y.; Li, S.; Zhang, L.; Sun, X.; Li, B.; et al. Yeast probiotic and yeast products in enhancing livestock feeds utilization and performance: An overview. J. Fungi 2022, 8, 1191. [Google Scholar] [CrossRef] [PubMed]
- Burdick Sanchez, N.C.; Broadway, P.R.; Carroll, J.A. Influence of yeast products on modulating metabolism and immunity in cattle and Swine. Animals 2021, 11, 371. [Google Scholar] [CrossRef] [PubMed]
- Keita, I.; Somda, M.K.; Savadogo, A.; Mogmenga, I.; Koita, O.; Traore, A.S. Isolation and molecular identification of yeast strains from “Rabilé” a starter of local fermented drink. Afr. J. Biotechnol. 2016, 15, 823–829. [Google Scholar]
- Keita, I.; Mogmenga, I.; Somda, M.K.; Doumbia, L.; Ouattara, A.S.; Koita, O.A.; Traore, A.S. Yeasts from a traditional ferment Rabile: Molecular and physiological characterization. Int. J. Biotechnol. Mol. Biol. Res. 2022, 12, 1–8. [Google Scholar]
- Mogmenga, I.; Dadiré, Y.; Somda, M.K.; Keita, I.; Ezeogu, L.I.; Ugwuanyi, J.; Traoré, A.S. Isolation and identification of indigenous yeasts from “Rabilé”, a starter culture used for production of traditional beer “dolo”, a condiment in Burkina Faso. Adv. Microbiol. 2019, 9, 646. [Google Scholar] [CrossRef] [Green Version]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Esteve-Zarzoso, B.; Belloch, C.; Uruburu, F.; Querol, A. Identification of yeasts by RFLP analysis of the 5.85 rRNA gene and the two ribosomal internal transcribed spacers. Int. J. Syst. Bacteriol. 1999, 49, 329–337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vauterin, L.; Vauterin, P. Computer-aided objective comparison of electrophoresis patterns for grouping and identification of microorganisms. Eur. Microbiol. 1992, 1, 37–41. [Google Scholar]
- Burns, P.; Vinderola, G.; Binetti, A.; Quiberoni, A.; de los Gavilán-Reyes, C.G.; Reinheimer, J. Bile-resistant derivatives obtained from non-intestinal dairy lactobacilli. Int. Dairy J. 2008, 18, 377–385. [Google Scholar] [CrossRef]
- Pedersen, L.L.; Owusu-Kwarteng, J.; Thorsen, L.; Jespersen, L. Biodiversity and probiotic potential of yeasts isolated from fura, a West African spontaneously fermented cereal. Int. J. Food Microbiol. 2012, 159, 144–151. [Google Scholar] [CrossRef] [PubMed]
- Binetti, A.; Carrasco, M.; Reinheimer, J.; Suárez, V. Yeasts from autochthonous cheese starters: Technological and functional properties. J. Appl. Microbiol. 2013, 115, 434–444. [Google Scholar] [CrossRef]
- Kos, B.; Šušković, J.; Vuković, S.; Šimpraga, M.; Frece, J.; Matošić, S. Adhesion and aggregation ability of probiotic strain Lactobacillus acidophillus M92. J. Appl. Microbiol. 2003, 94, 981–987. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Menezes, A.G.T.; Ramos, C.L.; Cenzi, G.; Melo, D.S.; Dias, D.R.; Schwan, R.F. Probiotic potential, antioxidant activity, and phytase production of indigenous yeasts isolated from indigenous fermented foods. Probiotics Antimicrob. Proteins 2020, 12, 280–288. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.-S.; Ma, Y.; Maubois, J.-L.; Chen, L.-J.; Liu, Q.-H.; Guo, J.-P. Identifcation of yeasts from raw milk and selection for some specific antioxidant properties. Int. J. Dairy Technol. 2010, 63, 47–54. [Google Scholar] [CrossRef]
- Greppi, A.; Saubade, F.; Botta, C.; Humblot, C.; Guyot, J.-P.; Cocolin, L. Potential probiotic Pichia kudriavzevii strains and their ability to enhance folate content of traditional cereal-based African fermented food. Food Microbiol. 2017, 62, 169–177. [Google Scholar] [CrossRef]
- Heitmann, M.; Zannini, E.; Arendt, E. Impact of Saccharomyces cerevisiae metabolites produced during fermentation on bread quality parameters: A review. Crit. Rev. Food Sci. Nutr. 2018, 58, 1152–1164. [Google Scholar] [CrossRef]
- Fernandez-Pacheco, P.; Arévalo-Villena, M.; Bevilacqua, A.; Corbo, M.R.; Briones Pérez, A. Probiotic characteristics in Saccharomyces cerevisiae strains: Properties for application in food industries. LWT 2018, 97, 332–340. [Google Scholar] [CrossRef]
- Fernández-Pacheco, P.; Rosa, I.Z.; Arévalo-Villena, M.; Gomes, E.; Pérez, A.B. Study of potential probiotic and biotechnological properties of non-Saccharomyces yeasts from fruit Brazilian ecosystems. Braz. J. Microbiol. 2021, 52, 2129–2144. [Google Scholar] [CrossRef]
- Trunk, T.; Khalil, H.S.; Leo, J.C. Bacterial autoaggregation. AIMS Microbiol. 2018, 4, 140–164. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Yeast Isolates | PCR ProductSize (bp) | Restriction Fragments (bp) | Identification | ||
|---|---|---|---|---|---|
| HinfI | HhaI | HaeIII | |||
| BB 1-2; BB 2-3; BB 3-2; BB 3-5; Bf 2-4; Bf 2-6; Bf 2-7; Dr 1-2; Dr 1-5; Dd 3-4; Fd 2-6; Fd 2-8; Ga 2-10; Ga 3-3 | 850 | 370 + 110 | 370 + 320 + 150 | 320 + 230 + 170 + 130 | Saccharomyces cerevisiae |
| BB 3-7; Dr 1-8 | 510 | 220 + 150 | 210 +180 + 70 | 380 + 90 | Pichia kudriavzevii |
| BB 3-3 | 620 | 340 + 210 + 70 | 300 + 220 + 90 | 410 + 210 | Rhodotorula mucilaginosa |
| Strains | At 37 °C | Survival Rate (%) | |
|---|---|---|---|
| DO600 nm | 0.3% Pepsin and pH 2.5 | 0.3% Bile Salts | |
| BB 3-7 | 2.58 ± 0.25 f | 96.96 ± 1.27 abc | 100.00 ± 0.00 a |
| Dr 1-8 | 1.68 ± 0.11 g | 96.30 ± 1.42 abc | 100.00 ± 00 a |
| BB 3-3 | 0.36 ± 0.01 h | 91.55 ± 4.97 cdef | 97.79 ± 1.07 b |
| BB 1-2 | 2.73 ± 0.17 f | 99.98 ± 0.00 a | 98.65 ± 1.29 ab |
| BB 2-3 | 3.32 ± 0.11 cd | 95.41 ± 6.97 abcde | 95.41 ± 2.74 c |
| BB 3-2 | 3.30 ± 0.05 d | 88.46 ± 3.34 ef | 99.25 ± 0.65 ab |
| BB 3-5 | 4.03 ± 0.10 ab | 91.22 ± 1.92 cdef | 99.36 ± 0.33 ab |
| Bf 2-4 | 3.69 ± 0.16 bc | 86.01 ± 1.98 f | 100.00 ± 0.00 a |
| Bf 2-6 | 2.87 ± 0.05 ef | 93.24 ± 5.41 abcde | 99.55 ± 0.56 ab |
| Bf 2-7 | 2.90 ± 0.04 ef | 95.60 ± 3.41 abcd | 95.56 ± 0.43 c |
| Dr 1-2 | 2.81 ± 0.12 f | 96.77 ± 1.94 abc | 98.73 ± 1.16 ab |
| Dr 1-5 | 2.25 ± 0.00 f | 89.03 ± 9.45 def | 99.53 ± 0.80 ab |
| Dd 3-4 | 3.19 ± 0.17 de | 92.36 ± 7.08 bcdef | 99.80 ± 0.33 ab |
| Fd 2-6 | 2.68 ± 0.03 f | 97.82 ± 1.54 abc | 99.51 ± 0.60 ab |
| Fd 2-8 | 3.36 ± 0.13 cd | 98.69 ± 1.37 ab | 100.00 ± 00 a |
| Ga 2-10 | 4.27 ± 0.05 a | 96.59 ± 1.75 abc | 100.00 ± 00 a |
| Ga 3-3 | 3.46 ± 0.08 cd | 96.95 ± 3.75 abc | 99.12 ± 1.51 ab |
| Strains | Auto-Aggregation (%) | Co-Aggregation (%) | Hydrophobicity (%) | |
|---|---|---|---|---|
| E. coli | S. enterica serovar Typhimurium | |||
| BB 3-7 | 81.25 ± 1.83 ab | 63.27 ±3.60 abc | 74.06 ± 7.94 a | 60.97 ± 2.20 abc |
| Dr 1-8 | 86.53 ± 4.82 a | 65.04 ± 7.56 abc | 67.44 ± 5.03 ab | 68.84 ± 5.23 a |
| BB 3-3 | 89.40 ± 2.30 a | 80.68 ± 9.53 a | 70.83 ± 5.05 a | 68.99 ± 5.37 a |
| BB 3-2 | 87.17 ± 2.86 a | 65.37 ± 4.35 abc | 69.38 ± 3.05 a | 51.99 ± 4.61 bcde |
| BB 1-2 | 70.20 ± 10.53 b | 50.15 ± 4.16 bcd | 66.38 ± 5.50 abc | 43.17 ± 5.07 e |
| BB 2-3 | 79.89 ± 7.08 ab | 56.16 ± 5.56 bcd | 57.9 ± 6.08 abcd | 63.09 ± 5.18 ab |
| BB 3-5 | 85.43 ± 5.25 a | 36.06 ± 3.45 de | 45.71 ± 4.35 bcd | 59.17 ± 4.88 abcd |
| Bf 2-4 | 84.38 ± 4.96 a | 24.92 ± 3.96 e | 40.89 ± 8.18 d | 60.11 ± 3.06 abc |
| Bf 2-6 | 89.11 ± 2.40 a | 49.91 ± 4.58 bcd | 58.82 ± 6.70 abcd | 67.97 ± 2.12 a |
| Bf 2-7 | 87.91 ± 3.47 a | 54.39 ± 4.90 bcd | 61.97 ± 2.71 abcd | 67.00 ± 2.45 a |
| Dr 1-2 | 89.13 ± 3.55 a | 72.74 ± 8.02 ab | 68.76 ± 5.00 a | 60.06 ± 2.52 abcd |
| Dr 1-5 | 81.48 ± 5.77 ab | 38.82 ± 10.8 de | 44.55 ± 7.09 cd | 64.53 ± 4.90 a |
| Dd 3-4 | 87.55 ± 2.28 a | 66.33 ± 5.56 abc | 61.72 ± 5.19 abcd | 48.23 ± 0.94 de |
| Fd 2-6 | 88.97 ± 1.54 a | 43.83 ± 7.03 cde | 59.74 ± 5.03 abcd | 70.73 ± 2.42 a |
| Fd 2-8 | 88.93 ± 1.74 a | 45.43 ± 13.15 cde | 61.21 ± 9.99 abcd | 65.11 ± 4.25 a |
| Ga 2-10 | 91.82 ± 1.96 a | 66.58 ± 7.02 abc | 72.78 ± 11.53 a | 63.04 ± 3.47 ab |
| Ga 3-3 | 84.34 ± 2.27 a | 65.49 ±4.6 abc | 66.16 ± 5.29 abc | 49.89 ± 3.41 cde |
| Strains | E. coli ATCC 8739 | S. aureus ATCC 33592 | L. monocytogenes ATCC 13932 | S. enteritidis ATCC 13076 | S. typhimurium ATCC 14028 |
|---|---|---|---|---|---|
| Fd 2-6 | 23.66 ± 1.15 a | 24.33 ± 0.57 a | 26.66 ± 2.88 a | 27.00 ± 2.64 a | 24.66 ± 0.57 a |
| Dd 3-4 | 23.33 ± 2.88 a | 21.33 ± 2.30 ab | 20.33 ± 0.57 bc | 26.00 ± 1.00 a | 25.00 ± 1.00 a |
| Ga 2-10 | 21.66 ± 2.88 ab | 21.66 ± 0.57 ab | 20.00 ± 0.00 bc | 24.33 ± 1.15 ab | 20.33 ± 0.57 bc |
| BB 3-2 | 20.00 ± 0.00 ab | 20.00 ± 0.00 abc | 30.00 ± 0.00 a | 25.00 ± 0.00 ab | 25.00 ± 0.00 a |
| Bf 2-6 | 20.00 ± 0.00 ab | 20.00 ± 0.00 abc | 21.33 ± 1.15 b | 24.66 ± 0.57 ab | 21.33 ± 1.15 ab |
| Fd 2-8 | 19.33 ± 1.15 abc | 20.00 ± 0.00 abc | 15.00 ± 0.00 de | 20.00 ± 0.00 cd | 16.33 ± 1.52 def |
| Ga 3-3 | 19.33 ± 1.15 abc | 19.33 ± 0.57 bcd | 15.00 ± 0.00 de | 18.66 ± 1.15 cd | 16.33 ± 0.57 def |
| Bf 2-4 | 17.33 ± 2.30 bcd | 20.00 ± 0.00 abc | 20.00 ± 0.00 bc | 21.33 ± 1.15 bc | 17.66 ± 2.08 bcde |
| BB 1-2 | 15.00 ± 0.00 cd | 16.66 ± 2.88 cde | 17.33 ± 2.51 cd | 20.00 ± 2.00 cd | 17.33 ± 2.51 cde |
| Dr 1-2 | 15.00 ± 0.00 cd | 16.66 ± 2.88 cde | 20.00 ± 0.00 bc | 17.33 ± 0.57 de | 17.66 ± 0.57 bcde |
| BB 3-5 | 13.66 ± 1.52 de | 14.33 ± 0.57 ef | 16.00 ± 1.73 d | 16.33 ± 0.57 de | 14.33 ± 0.57 efg |
| BB 2-3 | 13.33 ± 2.08 de | 14.33 ± 0.57 ef | 12.00 ± 1.73 e | 17.33 ± 1.15 de | 13.33 ± 1.15 fg |
| Bf 2-7 | 13.00 ± 1.00 de | 15.00 ± 0.00 def | 16.66 ± 0.57 cd | 17.66 ± 2.51 cd | 18.33 ± 0.57 bcd |
| Dr 1-5 | 10.00 ± 0.00 e | 11.00 ± 3.60 f | 14.33 ± 1.15 de | 13.66 ± 1.52 e | 10.66 ± 1.15 g |
| BB 3-7 | 5.33 ± 0.57 f | 6.33 ± 0.57 g | 5.33 ± 0.57 f | 5.33 ± 0.57 f | 4.33 ± 1.52 h |
| Dr 1-8 | 4.00 ± 1.73 fg | 5.00 ± 0.00 g | 6.00 ± 1.00 f | 5.00 ± 0.00 f | 4.00 ± 1.73 h |
| BB 3-3 | 0.00 ± 0.00 g | 0.00 ± 0.00 h | 0.00 ± 0.00 g | 0.00 ± 0.00 g | 0.00 ± 0.00 i |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mogmenga, I.; Somda, M.K.; Ouattara, C.A.T.; Keita, I.; Dabiré, Y.; Diguță, C.F.; Toma, R.C.; Ezeogu, L.I.; Ugwuanyi, J.O.; Ouattara, A.S.; et al. Promising Probiotic Properties of the Yeasts Isolated from Rabilé, a Traditionally Fermented Beer Produced in Burkina Faso. Microorganisms 2023, 11, 802. https://doi.org/10.3390/microorganisms11030802
Mogmenga I, Somda MK, Ouattara CAT, Keita I, Dabiré Y, Diguță CF, Toma RC, Ezeogu LI, Ugwuanyi JO, Ouattara AS, et al. Promising Probiotic Properties of the Yeasts Isolated from Rabilé, a Traditionally Fermented Beer Produced in Burkina Faso. Microorganisms. 2023; 11(3):802. https://doi.org/10.3390/microorganisms11030802
Chicago/Turabian StyleMogmenga, Iliassou, Marius Kounbèsiounè Somda, Cheik Amadou Tidiane Ouattara, Ibrahim Keita, Yérobessor Dabiré, Camelia Filofteia Diguță, Radu Cristian Toma, Lewis I. Ezeogu, Jerry O. Ugwuanyi, Aboubakar S. Ouattara, and et al. 2023. "Promising Probiotic Properties of the Yeasts Isolated from Rabilé, a Traditionally Fermented Beer Produced in Burkina Faso" Microorganisms 11, no. 3: 802. https://doi.org/10.3390/microorganisms11030802