
Assessing the Safety and Probiotic Characteristics of Lacticaseibacillus rhamnosus X253 via Complete Genome and Phenotype Analysis

Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Culture Conditions
2.2. Extraction of Nucleic Acids and Genome Sequencing
2.3. Genomic Annotation and Analysis
2.4. Evolutionary Position of L. rhamnosus X253
2.5. Safety Assessment of L. rhamnosus X253
2.5.1. Prediction of Safety-Related Genes in the Genome
2.5.2. Hemolytic Assay
2.5.3. Enzymatic Profiles by API ZYM
2.5.4. Antimicrobial Susceptibility Test
2.6. Assessment of Probiotic Properties of L. rhamnosus X253
2.6.1. Prediction of Probiotic-Related Genes in the Genome
2.6.2. Carbohydrate Utilization
2.6.3. Artificial Gastric and Bile Salt Tolerance Assay
2.6.4. Hydrophobicity and Auto-Aggregation
2.6.5. Adhesion Assay to CaCo-2 Cells
2.6.6. Tolerance to Hydrogen Peroxide
2.6.7. In Vitro Assessment of Antioxidant Activity
2.7. Statistical Analysis
3. Results and Discussion
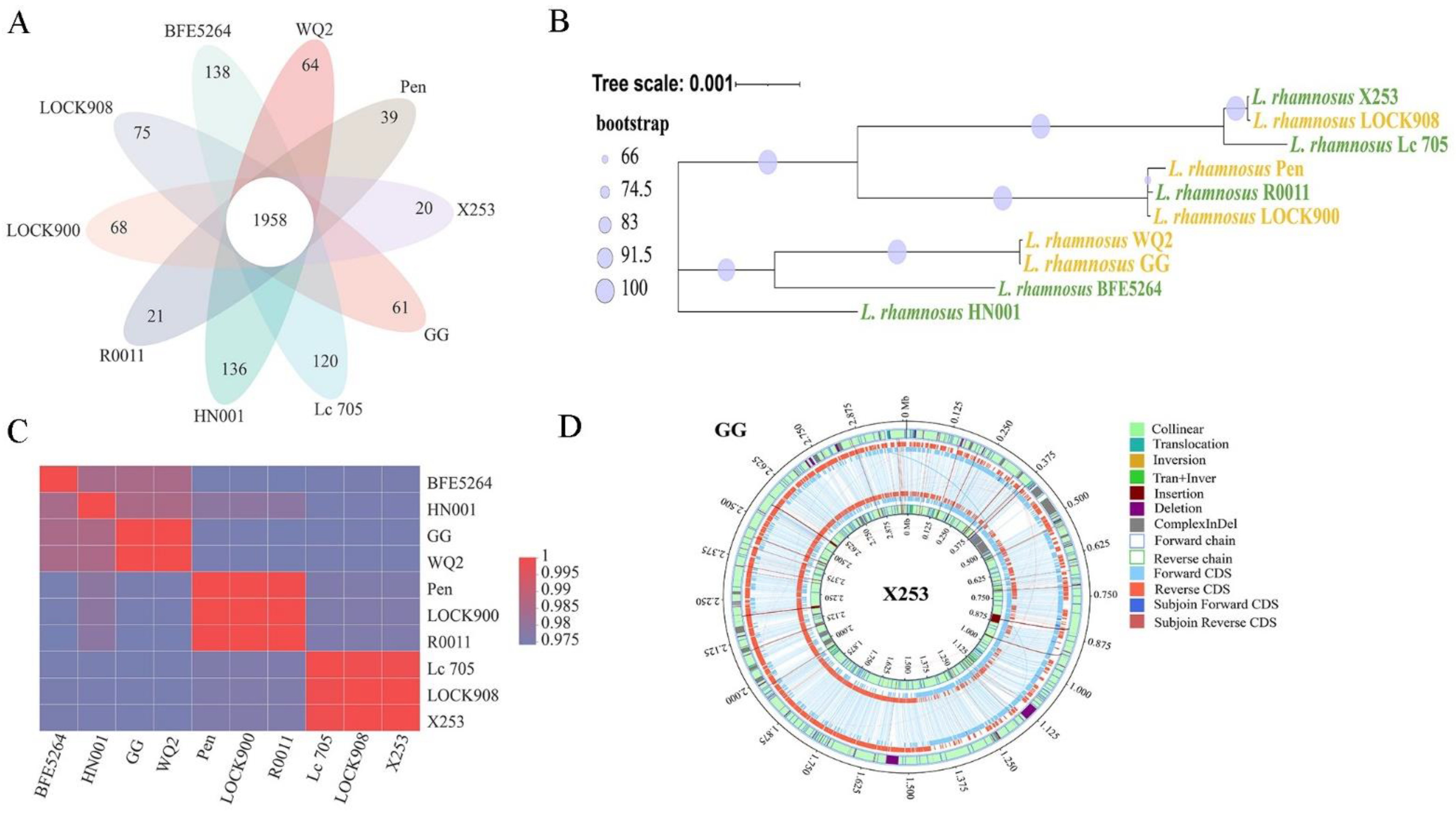
3.1. Genome Properties
3.2. Phylogenetic Analyses
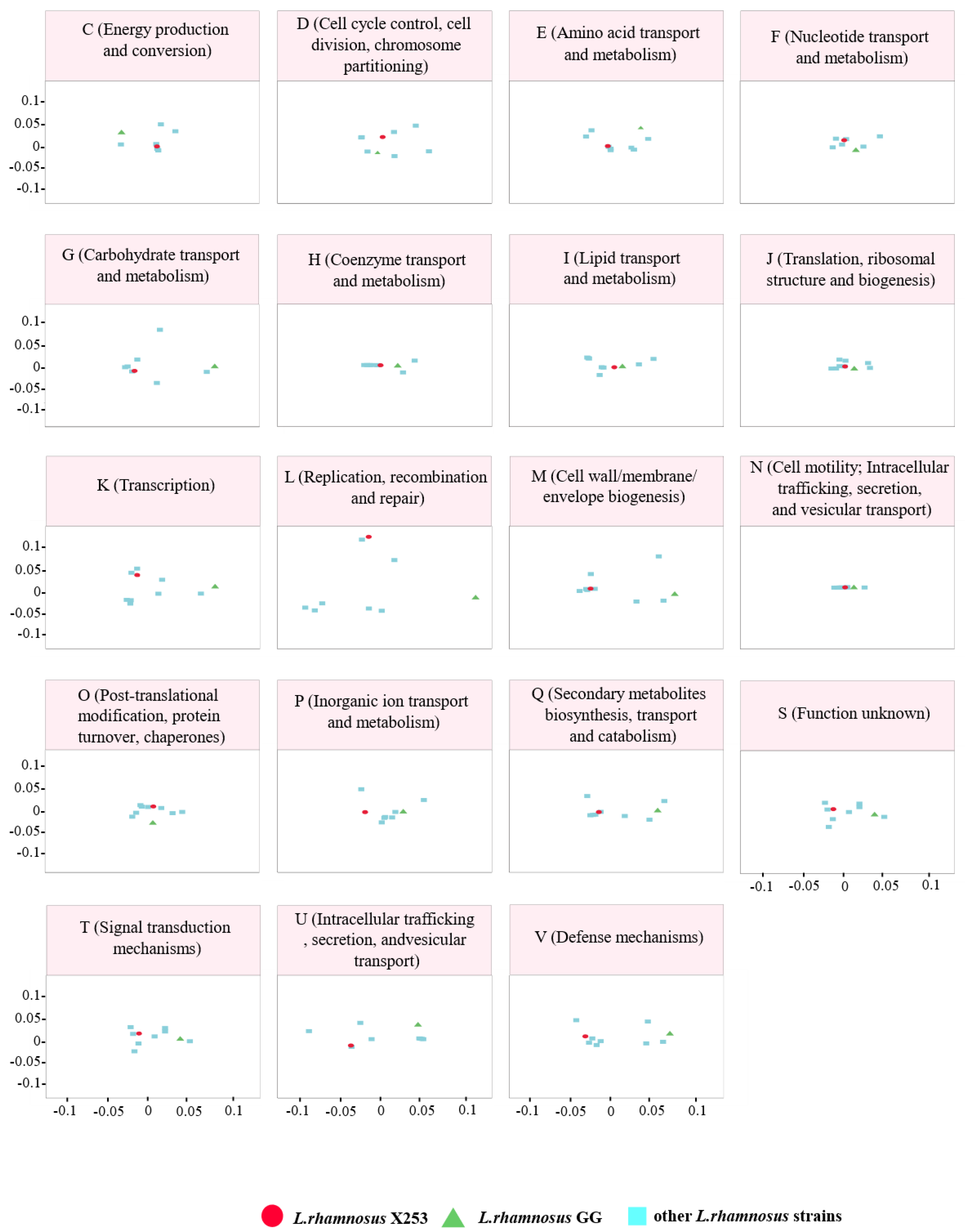
3.3. Comparative Functional Analysis between the X253 and GG Genomes
3.4. Safety Evaluation of L. rhamnosus X253
3.4.1. Antibiotic Resistance
3.4.2. Virulence Factor Genes and Toxin-Encoding Genes
3.4.3. Enzymatic Profile by API ZYM
3.5. Assessment of Probiotic Properties
3.5.1. Carbohydrate Fermentation Profiles
3.5.2. Artificial Gastric Fluid and Intestinal Fluid Tolerance Assays
3.5.3. Adhesion Ability
3.5.4. Hydrogen Peroxide Tolerance Assay
3.5.5. Antioxidant Activity In Vitro
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Huang, Y.Y.; Liu, D.M.; Jia, X.Z.; Liang, M.H.; Lu, Y.; Liu, J. Whole genome sequencing of Lactobacillus plantarum DMDL 9010 and its effect on growth phenotype under nitrite stress. LWT-Food Sci. Technol. 2021, 149, 111778. [Google Scholar] [CrossRef]
- Wu, Y.P.; Liu, D.M.; Zhao, S.; Huang, Y.Y.; Yu, J.J.; Zhou, Q.Y. Assessing the safety and probiotic characteristics of Bacillus coagulans 13002 based on complete genome and phenotype analysis. LWT-Food Sci. Technol. 2022, 155, 112847. [Google Scholar] [CrossRef]
- Damodharan, K.; Palaniyandi, S.A.; Yang, S.H.; Suh, J.W. Functional probiotic characterization and in vivo cholesterol-lowering activity of Lactobacillus helveticus isolated from fermented cow milk. J. Microbiol. Biotechnol. 2016, 26, 1675–1686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peirotén, Á.; Landete, J.M. Natural and engineered promoters for gene expression in Lactobacillus species. Appl. Microbiol. Biotechnol. 2020, 104, 3797–3805. [Google Scholar] [CrossRef] [PubMed]
- Stavropoulou, E.; Bezirtzoglou, E. Probiotics in medicine: A long debate. Front. Immunol. 2020, 11, 2192. [Google Scholar] [CrossRef] [PubMed]
- EFSA Panel on Biological Hazards (BIOHAZ); Koutsoumanis, K.; Allende, A.; Alvarez-Ordóñez, A.; Bolton, D.; Bover-Cid, S.; Chemaly, M.; Davies, R.; De Cesare, A.; Hilbert, F.; et al. Update of the list of QPS-recommended biological agents intentionally added to food or feed as notified to EFSA 15: Suitability of taxonomic units notified to EFSA until September 2021. EFSA J. 2022, 20, e07045. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.; Yang, B.; Chen, Y.; Stanton, C.; Ross, R.P.; Zhao, J.; Zhang, H.; Chen, W. Comparative genomic analyses of Lactobacillus rhamnosus isolated from Chinese subjects. Food Biosci. 2020, 36, 100659. [Google Scholar] [CrossRef]
- Shehata, H.R.; Newmaster, S.G. A validated real-time PCR method for the specific identification of probiotic strain Lactobacillus rhamnosus GG (ATCC 53103). J. AOAC Int. 2020, 103, 1604–1609. [Google Scholar] [CrossRef]
- Segers, M.E.; Lebeer, S. Towards a better understanding of Lactobacillus rhamnosus GG—Host interactions. Microb. Cell Fact. 2014, 13, S7. [Google Scholar] [CrossRef] [Green Version]
- Kankainen, M.; Paulin, L.; Tynkkynen, S.; von Ossowski, I.; Reunanen, J.; Partanen, P.; Satokari, R.; Vesterlund, S.; Hendrickx, A.P.; Lebeer, S.; et al. Comparative genomic analysis of Lactobacillus rhamnosus GG reveals pili containing a human-mucus binding protein. Proc. Natl. Acad. Sci. USA 2009, 106, 17193–17198. [Google Scholar] [CrossRef]
- Yau, Y.F.; El-Nezami, H.; Galano, J.M.; Kundi, Z.M.; Durand, T.; Lee, J.C.Y. Lactobacillus rhamnosus GG and oat beta-glucan regulated fatty acid profiles along the gut-liver-brain axis of mice fed with high fat diet and demonstrated antioxidant and anti-inflammatory potentials. Mol. Nutr. Food Res. 2020, 64, 2000566. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Yu, X.; Wang, D.; Gui, Y.; Wang, C.; Li, Q.; Wang, J.; Yin, B.; Pan, Z.; Gu, R. Rapid strain-specific identification of two Lactobacillus rhamnosus strains using PCR based on gene family analysis. LWT-Food Sci. Technol. 2021, 146, 111395. [Google Scholar] [CrossRef]
- Fu, X.; Lyu, L.; Wang, Y.; Zhang, Y.; Guo, X.; Chen, Q.; Liu, C. Safety assessment and probiotic characteristics of Enterococcus lactis JDM1. Microb. Pathog. 2022, 163, 105380. [Google Scholar] [CrossRef] [PubMed]
- Saroj, D.B.; Gupta, A.K. Genome based safety assessment for Bacillus coagulans strain LBSC (DSM 17654) for probiotic appli-cation. Int. J. Food Microbiol. 2020, 318, 108523. [Google Scholar] [CrossRef]
- Hussein, W.E.; Abdelhamid, A.G.; Rocha-Mendoza, D.; García-Cano, I.; Yousef, A.E. Assessment of safety and probiotic traits of Enterococcus durans OSY-EGY, isolated from Egyptian artisanal cheese, using comparative genomics and phenotypic analyses. Front. Microbiol. 2020, 11, 608314. [Google Scholar] [CrossRef] [PubMed]
- Emam, A.; Wu, X.; Xu, S.; Wang, L.; Liu, S.; Wang, B. Stalled replication fork protection limits cGAS–STING and P-body-dependent innate immune signalling. Nat. Cell Biol. 2022, 24, 1154–1164. [Google Scholar] [CrossRef] [PubMed]
- Xiang, Y.; Wang, Y.; Shen, H.; Wang, D. The draft genome sequence of Pseudomonas putida strain TGRB4, an aerobic bacterium capable of producing methylmercury. Curr. Microbiol. 2020, 77, 522–527. [Google Scholar] [CrossRef]
- Ardui, S.; Ameur, A.; Vermeesch, J.; Hestand, M. Single molecule real-time (SMRT) sequencing comes of age: Applications and utilities for medical diagnostics. Nucleic Acids Res. 2018, 46, 2159–2168. [Google Scholar] [CrossRef] [Green Version]
- Myers, E.W.; Sutton, G.G.; Delcher, A.L.; Dew, I.M.; Fasulo, D.P.; Flanigan, M.J.; Kravitz, S.A.; Mobarry, C.M.; Reinert, K.H.; Remington, K.A.; et al. A whole-genome assembly of Drosophila. Science 2000, 287, 2196–2204. [Google Scholar] [CrossRef]
- Besemer, J.; Lomsadze, A.; Borodovsky, M. GeneMarkS: A self-training method for prediction of gene starts in microbial genomes. Nucleic Acids Res. 2001, 29, 2607–2618. [Google Scholar] [CrossRef]
- Jin, Q.; Jiang, Q.; Zhao, L.; Su, C.; Li, S.; Si, F.; Li, S.; Zhou, C.; Mu, Y.; Xiao, M. Complete genome sequence of Bacillus velezensis S3-1, a potential biological pesticide with plant pathogen inhibiting and plant promoting capabilities. J. Biotechnol. 2017, 259, 199–203. [Google Scholar] [CrossRef] [PubMed]
- Juhl, J.L.; Philippe, J.; Michael, K.; Christian, V.M.; Jean, M.; Tobias, D.; Peer, B. eggNOG: Automated construction and anno-tation of orthologous groups of genes. Nucleic Acids Res. 2008, 36, D250–D254. [Google Scholar]
- Ogata, H.; Goto, S.; Sato, K.; Fujibuchi, W.; Bono, H.; Kanehisa, M. KEGG: Kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 1999, 27, 29–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harris, H.M.B.; Bourin, M.J.B.; Claesson, M.J.; O’Toole, P.W. Phylogenomics and comparative genomics of Lactobacillus sali-varius, a mammalian gut commensal. Microb. Genom. 2017, 3, e000115. [Google Scholar] [PubMed]
- Kelleher, P.; Bottacini, F.; Mahony, J.; Kilcawley, K.N.; van Sinderen, D. Comparative and functional genomics of the Lacto-coccus lactis taxon; insights into evolution and niche adaptation. BMC Genom. 2017, 18, 267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, L.T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. Interactive tree of life (iTOL) v3: An online tool for the display and annotation of phylogenetic and other trees. Nucleic Acids Res. 2016, 44, W242–W245. [Google Scholar] [CrossRef]
- Guo, Y.; Li, Y.; Su, L.; Chang, D.; Liu, W.; Wang, T.; Yuan, Y.; Fang, X.; Wang, J.; Li, T.; et al. Comparative genomic analysis of Klebsiella pneumonia (LCT-KP214) and a mutant strain (LCT-KP289) obtained after spaceflight. BMC Genom. 2014, 15, 589. [Google Scholar] [CrossRef] [Green Version]
- Petrova, M.I.; Macklaim, J.M.; Wuyts, S.; Verhoeven, T.; Vanderleyden, J.; Gloor, G.B.; Lebeer, S.; Reid, G. Comparative genomic and phenotypic analysis of the vaginal probiotic Lactobacillus rhamnosus GR-1. Front. Microbiol. 2018, 9, 1278. [Google Scholar] [CrossRef] [Green Version]
- Jeon, H.L.; Lee, N.K.; Yang, S.J.; Kim, W.S.; Paik, H.D. Probiotic characterization of Bacillus subtilis P223 isolated from kimchi. Food Sci. Biotechnol. 2017, 26, 1641–1648. [Google Scholar] [CrossRef]
- Jang, Y.J.; Gwon, H.M.; Jeong, W.S.; Yeo, S.H.; Kim, S.Y. Safety evaluation of Weissella cibaria JW15 by phenotypic and genotypic property analysis. Microorganisms 2021, 9, 2450. [Google Scholar] [CrossRef] [PubMed]
- EFSA Panel on Additives and Products or Substances Used in Animal Feed (FEEDAP). Guidance on the assessment of bacterial susceptibility to anti-microbials of human and veterinary importance. EFSA J. 2012, 10, 2740. [Google Scholar]
- Huang, Y.; Adams, M.C. In vitro assessment of the upper gastrointestinal tolerance of potential probiotic dairy propioni-bacteria. Int. J. Food Microbiol. 2004, 91, 253–260. [Google Scholar] [CrossRef] [PubMed]
- Sadishkumar, V.; Jeevaratnam, K. In vitro probiotic evaluation of potential antioxidant lactic acid bacteria isolated from idli batter fermented with Piper betle leaves. Int. J. Food Sci. Technol. 2017, 52, 329–340. [Google Scholar] [CrossRef]
- Todorov, S.D.; Holzapfel, W.; Nero, L.A. In vitro evaluation of beneficial properties of bacteriocinogenic Lactobacillus plantarum ST8Sh. Probiotics Antimicrob. Proteins 2017, 9, 194–203. [Google Scholar] [CrossRef] [Green Version]
- Tang, W.; Xing, Z.; Li, C.; Wang, J.; Wang, Y. Molecular mechanisms and in vitro antioxidant effects of Lactobacillus plantarum MA2. Food Chem. 2017, 221, 1642–1649. [Google Scholar] [CrossRef]
- Chen, P.; Zhang, Q.; Dang, H.; Liu, X.; Tian, F.; Zhao, J.; Chen, Y.; Zhang, H.; Chen, W. Screening for potential new probiotic based on probiotic properties and α-glucosidase inhibitory activity. Food Control 2014, 35, 65–72. [Google Scholar] [CrossRef]
- Lebeer, S.; Vanderleyden, J.; De Keersmaecker, S.C.J. Genes and molecules of lactobacilli supporting probiotic action. Microbiol. Mol. Biol. Rev. 2008, 72, 728–764. [Google Scholar] [CrossRef] [Green Version]
- Jeong, H.; Choi, S.; Park, G.S.; Ji, Y.; Park, S.; Holzapfel, W.; Mathara, J.; Kang, J. Comparative genomic analysis of Lactobacillus rhamnosus BFE5264, a probiotic strain isolated from traditional maasai fermented milk. Microbiol. Biotechnol. Lett. 2019, 47, 25–33. [Google Scholar] [CrossRef]
- Sun, Z.; Zhang, W.; Guo, C.; Yang, X.; Liu, W.; Wu, Y.; Song, Y.; Kwok, L.Y.; Cui, Y.; Menghe, B.; et al. Comparative genomic analysis of 45 type strains of the genus Bifidobacterium: A snapshot of its genetic diversity and evolution. PLoS ONE 2015, 10, e0117912. [Google Scholar] [CrossRef] [Green Version]
- Takashima, K.; Nagao, S.; Kizawa, A.; Suzuki, T.; Dohmae, N.; Hihara, Y. The role of transcriptional repressor activity of LexA in salt-stress responses of the cyanobacterium Synechocystis sp. PCC 6803. Sci. Rep. 2020, 10, 17393. [Google Scholar] [CrossRef] [PubMed]
- Wahyudi, H.; McAlpine, S.R. Predicting the unpredictable: Recent structure–activity studies on peptide-based macrocycles. Bioorg. Chem. 2015, 60, 74–97. [Google Scholar] [CrossRef] [PubMed]
- Selva, E.; Beretta, G.; Montanini, N.; Saddler, G.S.; Gastaldo, L.; Ferrari, P.; Lorenzetti, R.; Landini, P.; Ripamonti, F.; Goldstein, B.P.; et al. Antibiotic GE2270 a: A novel inhibitor of bacterial protein synthesis. I. Isolation and characterization. J. Antibiot. 1991, 44, 693–701. [Google Scholar] [CrossRef] [PubMed]
- Korhonen, J.M.; Van Hoek, A.H.; Saarela, M.; Huys, G.; Tosi, L.; Mayrhofer, S.; Wright, A.V. Antimicrobial susceptibility of Lactobacillus rhamnosus. Benefic. Microbes 2010, 1, 75–80. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, S.; Kehrenberg, C.; Doublet, B.; Cloeckaert, A. Molecular basis of bacterial resistance to chloramphenicol and florfenicol. FEMS Microbiol. Rev. 2004, 28, 519–542. [Google Scholar] [CrossRef] [Green Version]
- Heavey, P.M.; Rowland, I.R. Microbial-gut interactions in health and disease. Gastrointestinal cancer. Best Pract. Res. Clin. Gastroenterol. 2004, 18, 323–336. [Google Scholar] [CrossRef]
- Becerra, J.E.; Yebra, M.J.; Monedero, V. An L-fucose operon in the probiotic Lactobacillus rhamnosus GG is involved in adaptation to gastrointestinal conditions. Appl. Environ. Microbiol. 2015, 81, 3880–3888. [Google Scholar] [CrossRef] [Green Version]
- Hamon, E.; Horvatovich, P.; Marchioni, E.; Aoudé-Werner, D.; Ennahar, S. Investigation of potential markers of acid resistance in Lactobacillus plantarum by comparative proteomics. J. Appl. Microbiol. 2013, 116, 134–144. [Google Scholar] [CrossRef]
- Jia, F.F.; Zhang, L.J.; Pang, X.H.; Gu, X.X.; Abdelazez, A.; Liang, Y.; Sun, S.R.; Meng, X.C. Complete genome sequence of bac-teriocin-producing Lactobacillus plantarum KLDS1.0391, a probiotic strain with gastrointestinal tract resistance and adhesion to the intestinal epithelial cells. Genomics 2017, 109, 432–437. [Google Scholar] [CrossRef]
- Futai, M.; Kanazawa, H. Structure and function of proton-translocating adenosine triphosphatase (F0F1). Microbiol. Rev. 1983, 47, 285–312. [Google Scholar] [CrossRef]
- El Aila, N.A.; Tency, I.; Claeys, G.; Verstraelen, H.; Saerens, B.; Santiago, G.L.; De Backer, E.; Cools, P.; Temmerman, M.; Verhelst, R.; et al. Identification and genotyping of bacteria from paired vaginal and rectal samples from pregnant women indicates similarity between vaginal and rectal microflora. BMC Infect. Dis. 2009, 9, 167. [Google Scholar] [CrossRef] [Green Version]
- Riaz, M.S.; Mehwish, M.; Siddiq, M.; Haobin, Z.; Zhu, J.; Yan, L.; Shao, D.; Xu, X.; Shi, J. Identification, characterization, and probiotic potential of Lactobacillus rhamnosus isolated from human milk. LWT-Food Sci. Technol. 2017, 84, 271–280. [Google Scholar] [CrossRef]
- Ye, K.; Li, P.; Gu, Q. Complete genome sequence analysis of a strain Lactobacillus pentosus ZFM94 and its probiotic character-istics. Genomics 2020, 112, 3142–3149. [Google Scholar] [CrossRef] [PubMed]
- Pastoriza, S.; Delgado-Andrade, C.; Haro, A.; Rufián-Henares, J.A. A physiologic approach to test the global antioxidant response of foods. The GAR method. Food Chem. 2011, 129, 1926–1932. [Google Scholar] [CrossRef]
- Forsyth, C.B.; Farhadi, A.; Jakate, S.M.; Tang, Y.; Shaikh, M.; Keshavarzian, A. Lactobacillus GG treatment ameliorates alcohol-induced intestinal oxidative stress, gut leakiness, and liver injury in a rat model of alcoholic steatohepatitis. Alcohol 2009, 43, 163–172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martarelli, D.; Verdenelli, M.C.; Scuri, S.; Cocchioni, M.; Silvi, S.; Cecchini, C.; Pompei, P. Effect of a probiotic intake on oxidant and antioxidant parameters in plasma of athletes during intense exercise training. Curr. Microbiol. 2011, 62, 1689–1696. [Google Scholar] [CrossRef]
- Garigan, D.; Hsu, A.L.; Fraser, A.G.; Kamath, R.S.; Ahringer, J.; Kenyon, C. Genetic analysis of tissue aging in Caenorhabditis elegans: A role for heat-shock factor and bacterial proliferation. Genetics 2002, 161, 1101–1112. [Google Scholar] [CrossRef]
- Kaushik, J.K.; Kumar, A.; Duary, R.K.; Mohanty, A.K.; Grover, S.; Batish, V.K. Functional and probiotic attributes of an in-digenous isolate of Lactobacillus plantarum. PLoS ONE 2009, 4, e8099. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Strain | Assembly No. | Source |
|---|---|---|---|
| L. rhamnosus | X253 | GCA_018228745.1 | Dairy |
| BFE5264 | GCA_001988935.1 | ||
| Lc 705 | GCA_000026525.1 | ||
| R0011 | GCA_000235785.2 | ||
| HN001 | GCA_000173255.2 | ||
| Pen | GCA_002076955.1 | Human intestine | |
| GG | GCA_000026505.1 | ||
| LOCK908 | GCA_000418495.1 | ||
| LOCK900 | GCA_000418475.1 | ||
| WQ2 | GCA_002025085.1 |
| Strain | Size (Mb) | Plasmids | CDS | GC% | rRNAs | tRNAs | Source |
|---|---|---|---|---|---|---|---|
| X253 | 2.99 | 0 | 2649 | 46.8 | 15 | 62 | Dairy |
| BFE5264 | 3.11 | 1 | 2785 | 46.8 | 15 | 60 | |
| Lc 705 | 3.03 | 1 | 2652 | 46.6 | 15 | 61 | |
| R0011 | 2.90 | N/A | 2618 | 46.7 | 11 | 54 | |
| HN001 | 2.91 | 2 | 2642 | 46.7 | 3 | 50 | |
| Pen | 2.88 | 0 | 2569 | 46.8 | 15 | 59 | Human intestine |
| GG | 3.01 | 0 | 2703 | 46.7 | 15 | 57 | |
| LOCK908 | 2.99 | 0 | 2666 | 46.8 | 15 | 62 | |
| LOCK900 | 2.88 | 0 | 2586 | 46.8 | 15 | 59 | |
| WQ2 | 2.96 | 0 | 2653 | 46.7 | 2 | 45 |
| Antibiotic | MIC Cut-Off Values (μg/mL) | MIC Observed (μg/mL) | Results | ||
|---|---|---|---|---|---|
| X253 | GG | X253 | GG | ||
| Ampicillin | 4 | 0.5 | 0.5 | S | S |
| Chloramphenicol | 4 | 8 | 8 | R | R |
| Erythromycin | 1 | 0.5 | 0.5 | S | S |
| Gentamicin | 16 | 16 | 8 | S | S |
| Streptomycin | 32 | 8 | 16 | S | S |
| Tetracycline | 8 | 0.5 | 1 | S | S |
| Vancomycin | N/R | >256 | >256 | R | R |
| Substrate | X253 | GG |
|---|---|---|
| Alkaline phosphatase | + | + |
| Esterase (C4) | + | + |
| Esterase lipase (C8) | + | + |
| Lipase (C14) | − | − |
| Leucine arylamidase | + | + |
| Valine arylamidase | + | + |
| Cystine arylamidase | + | + |
| Trypsin | − | − |
| α-Chymotrypsin | + | + |
| Acid phosphatase | + | + |
| Naphthol-AS-BI-phosphohydrolase | + | + |
| α-Galactosidase | − | − |
| β-Galactosidase | + | + |
| β-Glucuronidase | − | − |
| α-Glucosidase | + | + |
| β-Glucosidase | + | + |
| N-acetyl-glucosaminidase | − | − |
| α-Mannosidase | − | − |
| β-Fucosidase | + | + |
| Substrate | X253 | GG | Substrate | X253 | GG |
|---|---|---|---|---|---|
| Glycerol | − | − | Esculine | + | + |
| Erythritol | − | − | Salicine | + | + |
| D-arabinose | − | + | D-cellobiose | + | + |
| L-arabinose | − | − | D-maltose | + | − |
| D-ribose | − | − | D-lactose | + | − |
| D-xylose | + | − | D-melibiose | − | − |
| L-xylose | − | − | D-saccharose | + | − |
| D-adonitol | − | − | D-trehalose | + | + |
| Methyl-β-D-xylopyranoside | − | − | Inulin | − | − |
| D-galactose | + | + | D-melezitose | + | + |
| D-glucose | + | + | D-raffinose | − | − |
| D-fructose | + | + | Starch | − | − |
| D-mannose | + | + | Glycogen | − | − |
| L-sorbose | + | − | Xylitol | − | − |
| L-rhamnose | + | − | Gentiobiose | − | + |
| Dulcitol | − | + | D-turanose | + | − |
| Inositol | − | − | D-lyxose | − | − |
| D-mannitol | + | + | D-tagatose | + | + |
| D-sorbitol | + | + | D-fucose | − | − |
| Methyl-α-D-mannopyranoside | − | − | L-fucose | − | + |
| Methyl-α-D-glucopyranoside | + | − | D-arabitol | − | − |
| N-acetylglucosamine | + | + | L-arabitol | − | − |
| Amygdaline | + | + | Potassium gluconate | − | − |
| Arbutine | + | + | Potassium 2-cetogluconate | − | − |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, L.; Zhang, Y.; Liu, Y.; Zhong, J.; Zhang, D. Assessing the Safety and Probiotic Characteristics of Lacticaseibacillus rhamnosus X253 via Complete Genome and Phenotype Analysis. Microorganisms 2023, 11, 140. https://doi.org/10.3390/microorganisms11010140
Zhao L, Zhang Y, Liu Y, Zhong J, Zhang D. Assessing the Safety and Probiotic Characteristics of Lacticaseibacillus rhamnosus X253 via Complete Genome and Phenotype Analysis. Microorganisms. 2023; 11(1):140. https://doi.org/10.3390/microorganisms11010140
Chicago/Turabian StyleZhao, Lei, Yinan Zhang, Yang Liu, Jiang Zhong, and Dong Zhang. 2023. "Assessing the Safety and Probiotic Characteristics of Lacticaseibacillus rhamnosus X253 via Complete Genome and Phenotype Analysis" Microorganisms 11, no. 1: 140. https://doi.org/10.3390/microorganisms11010140