
Artificial Cultivation Changes Foliar Endophytic Fungal Community of the Ornamental Plant Lirianthe delavayi


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Population and Sample Source
2.2. High-Throughput Sequencing and Clustering into Amplified Sequence Variants (ASVs)
2.3. Statistical Analysis
3. Results
3.1. Changes in Diversity of Endophytic Fungi in L. delavayi Leaves
3.2. Endophytic Fungal Species Composition in L. delavayi Leaves
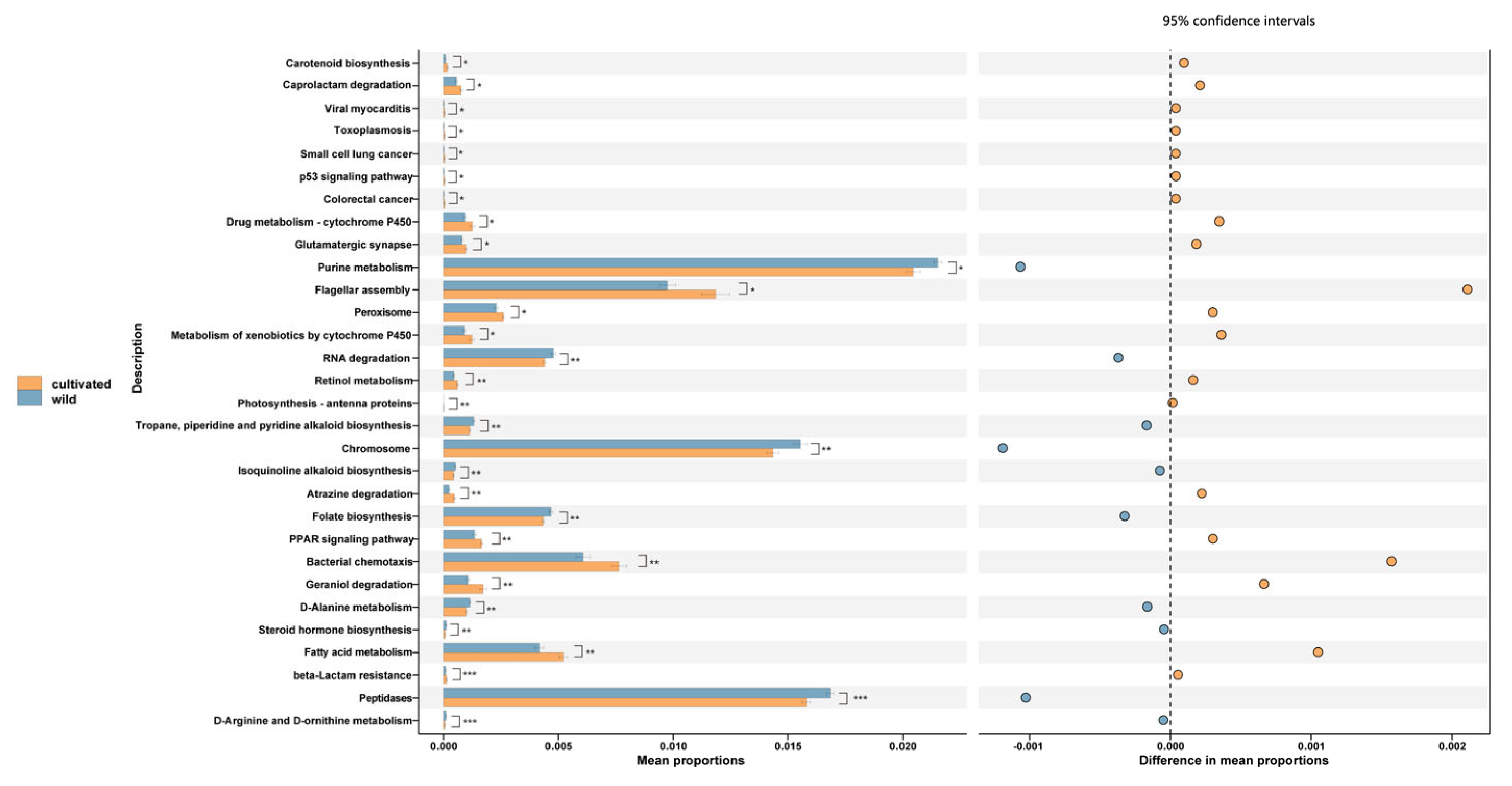
3.3. Functional Predictions of Endophytic Fungi in L. delavayi Leaves
4. Discussion
4.1. Impact of Artificial Cultivation on the Diversity of Endophytic Fungi in L. delavayi Leaves
4.2. Impact of Artificial Cultivation on the Species Composition of Endophytic Fungi in L. delavayi Leaves
4.3. Effect of Artificial Cultivation on the Functions of Endophytic Fungi Community in L. delavayi Leaves
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chen, M.; Arato, M.; Borghi, L.; Nouri, E.; Reinhardt, D. Beneficial Services of Arbuscular Mycorrhizal Fungi—From Ecology to Application. Front. Plant Sci. 2018, 9, 1270. [Google Scholar] [CrossRef] [PubMed]
- Martin, F.; Kohler, A.; Murat, C.; Veneault-Fourrey, C.; Hibbett, D.S. Unearthing the roots of ectomycorrhizal symbioses. Nat. Rev. Microbiol. 2016, 14, 760–773. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, R.J.; White, J.F., Jr.; Arnold, A.E.; Redman, A.R.A. Fungal endophytes: Diversity and functional roles. New Phytol. 2009, 182, 314–330. [Google Scholar] [CrossRef] [PubMed]
- Yan, L.; Zhu, J.; Zhao, X.; Shi, J.; Jiang, C.; Shao, D. Beneficial effects of endophytic fungi colonization on plants. Appl. Microbiol. Biotechnol. 2019, 103, 3327–3340. [Google Scholar] [CrossRef]
- Rana, K.L.; Kour, D.; Sheikh, I.; Yadav, N.; Yadav, A.N.; Kumar, V.; Singh, B.P.; Dhaliwal, H.S.; Saxena, A.K. Biodiversity of Endophytic Fungi from Diverse Niches and Their Biotechnological Applications. In Advances in Endophytic Fungal Research: Present Status and Future Challenges; Singh, B.P., Ed.; Springer International Publishing: Cham, Switzerland, 2019; pp. 105–144. ISBN 978-3-030-03589-1. [Google Scholar] [CrossRef]
- He, M. Research progress of plant endophytic fungi as biocontrol factors. Plant Prot. 2005, 31, 10–14. [Google Scholar]
- Wang, Y.-L.; Gao, C.; Chen, L.; Ji, N.-N.; Wu, B.-W.; Lü, P.-P.; Li, X.-C.; Qian, X.; Maitra, P.; Babalola, B.J.; et al. Community Assembly of Endophytic Fungi in Ectomycorrhizae of Betulaceae Plants at a Regional Scale. Front. Microbio. 2020, 10, 3105. [Google Scholar] [CrossRef]
- Cox, F.; Barsoum, N.; Lilleskov, E.A.; Bidartondo, M.I. Nitrogen availability is a primary determinant of conifer mycorrhizas across complex environmental gradients. Ecol. Lett. 2010, 13, 1103–1113. [Google Scholar] [CrossRef]
- Mcguire, K.L.; Bent, E.; Borneman, J.; Majumder, A.; Allison, S.D.; Treseder, K.K. Functional diversity in resource use by fungi. Ecology 2010, 91, 2324–2332. [Google Scholar] [CrossRef]
- Mundra, S.; Bahram, M.; Eidesen, P.B. Alpine bistort (Bistorta vivipara) in edge habitat associates with fewer but distinct ectomycorrhizal fungal species: A comparative study of three contrasting soil environments in Svalbard. Mycorrhiza 2016, 26, 809–818. [Google Scholar] [CrossRef]
- Barnes, C.J.; Van Der Gast, C.J.; Mcnamara, N.P.; Rowe, R.; Bending, G.D. Extreme rainfall affects assembly of the root-associated fungal community. New Phytol. 2018, 220, 1172–1184. [Google Scholar] [CrossRef] [Green Version]
- Miyamoto, Y.; Terashima, Y.; Nara, K. Temperature niche position and breadth of ectomycorrhizal fungi: Reduced diversity under warming predicted by a nested community structure. Glob. Chang. Biol. 2018, 24, 5724–5737. [Google Scholar] [CrossRef]
- Gooden, B.; Thompson, E.R.; French, K. Do native plant associations with arbuscular mycorrhizal fungi and dark septate endophytes differ between reconstructed and remnant coastal dunes? Plant Ecol. 2020, 221, 757–771. [Google Scholar] [CrossRef]
- Sikes, B.A.; Hawkes, C.V.; Fukami, T. Plant and root endophyte assembly history: Interactive effects on native and exotic plants. Ecology 2016, 97, 484–493. [Google Scholar] [CrossRef] [Green Version]
- Song, M.Y. The Application and Researches of Six Kinds Plants in Korla; Xinjiang Agricultural University: Urumqi, China, 2014. [Google Scholar]
- Pérez-Jaramillo, J.E.; Mendes, R.; Raaijmakers, J.M. Impact of plant domestication on rhizosphere microbiome assembly and functions. Plant Mol. Biol. 2016, 90, 635–644. [Google Scholar] [CrossRef] [Green Version]
- Qin, H.; Bai, J.F.; Xu, Q.F. Effects of arbuscular mycorrhizal fungal hyphae on soil microbial community composition and polychlorinated biphenyls degradation. Soils 2015, 47, 704–710. [Google Scholar] [CrossRef]
- Cao, Y.F.; Shen, Z.Z.; Liu, S.S. Evaluation of effect of Trichoderma controlling fusarium wilt disease and its influencing factors with meta-analysis in China. Acta Pedol. Sin. 2019, 56, 716–727. [Google Scholar] [CrossRef]
- Adrien, T.; Bellanger, J.-M.; Carcaillet, C.; Hugot, L.; Kjellberg, F.; Lecanda, A.; Lesne, A.; Moreau, P.-A.; Scharmann, K.; Leidel, S.; et al. Diversity of foliar endophytic ascomycetes in the endemic Corsican pine forests. Fungal Ecol. 2018, 36, 128–140. [Google Scholar]
- Christian, N.; Whitaker, B.K.; Clay, K. Microbiomes: Unifying animal and plant systems through the lens of community ecology theory. Front. Microbiol. 2015, 6, 869. [Google Scholar] [CrossRef] [Green Version]
- Hardoim, P.R.; van Overbeek, L.S.; Berg, G.; Pirttil€, A.M.; Compant, S.; Campisano, A.; Doring, M.; Sessitsch, A. The hidden world within plants: Ecological and evolutionary considerations for defining functioning of microbial endophytes. Microbiol. Mol. Biol. Rev. 2015, 79, 293–320. [Google Scholar] [CrossRef] [Green Version]
- Sapkota, R.; Knorr, K.; Jørgensen, L.N.; O’Hanlon, K.A.; Nicolaisen, M. Host genotype is an important determinant of the cereal phyllosphere mycobiome. New Phytol. 2015, 207, 1134–1144. [Google Scholar] [CrossRef]
- Flora of China Committee. Flora of China, Volume 30. Part 1; Science Press: Beijing, China, 1990; p. 112. [Google Scholar]
- Zhang, X. Application of Lirianthe Delavayi in Landscaping; Beijing Agriculture: Beijing, China, 2014; pp. 81–82. [Google Scholar]
- Natalie, C.; Courtney, S.; Noelle, D.V.; Keith, C. Plant Host and Geographic Location Drive Endophyte Community Composition in the Face of Perturbation. Microb. Ecol. 2016, 72, 621–632. [Google Scholar] [CrossRef]
- Gao, S.; Liu, X.C.; Dong, Z.; Liu, M.Y.; Dai, L.Y. Advance of phyllosphere microorganisms and their interaction with the outside environment. Plant Sci. J. 2016, 34, 654–661. [Google Scholar]
- Ihrmark, K.; Bödeker, I.; Cruz-Martinez, K.; Friberg, H.; Kubartova, A.; Schenck, J. New primers to amplify the fungal ITS2 region—Evaluation by 454-sequencing of artificial and natural communities. FEMS Microb. Ecol. 2012, 82, 666–677. [Google Scholar] [CrossRef] [PubMed]
- Benjamin, J.C.; Paul, J.M.; Michael, J.R. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [Green Version]
- Bolyen, E.; Jai, R.R.; Matthew, R.D. Author Correction: Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 1091. [Google Scholar] [CrossRef]
- Ricotta, C.; Podani, J. On some properties of the Bray-Curtis dissimilarity and their ecological meaning. Ecol. Complex. 2017, 31, 201–205. [Google Scholar] [CrossRef]
- Koljalg, U.; Nilsson, R.H.; Abarenkov, K. Towards a unified paradigm for sequence-basedidentification of fungi. Mol. Ecol. 2013, 22, 5271–5277. [Google Scholar] [CrossRef] [Green Version]
- Langille, M.G.I.; Zaneveld, J.; Caporaso, J.G.; McDonald, D.; Knights, D.; Reyes, J.A. Predictive functional profiling of microbial communities using 16S rRNA marker gene sequences. Nat. Biotechnol. 2013, 31, 814–821. [Google Scholar] [CrossRef]
- Chanumolu, S.K.; Mustafa, A.; Handan, C.; Otu, H.H. KEGG2Net: Deducing gene interaction networks and acyclic graphs from KEGG pathways. EMBnet J. 2021, 26, e949. [Google Scholar] [CrossRef]
- Kanehisa, M.; Sato, Y.; Kawashima, M. KEGG mapping tools for uncovering hidden features in biological data. Protein Sci. 2022, 31, 47–53. [Google Scholar] [CrossRef]
- Hassani, M.A.; Özkurt, E.; Franzenburg, S.; Stukenbrock, E.H. Ecological Assembly Processes of the Bacterial and Fungal Microbiota of Wild and Domesticated Wheat Species. Phytobiomes J. 2020, 4, 217–224. [Google Scholar] [CrossRef] [Green Version]
- Coleman-Derr, D.; Desgarennes, D.; Fonseca-Garcia, C.; Gross, S.; Clingenpeel, S.; Woyke, T.; North, G.; Visel, A.; Partida-Martinez, L.P.; Tringe, S.G. Plant compartment and biogeography affect microbiome composition in cultivated and native Agave species. New Phytol. 2016, 209, 798–811. [Google Scholar] [CrossRef] [Green Version]
- Xing, X.; Koch, A.; Jones, A.; Ragone, D.; Murch, S.; Hart, M. Mutualism breakdown in breadfruit domestication. Proc. R. Soc. B 2012, 279, 1122–1130. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Jaramillo, J.; de Hollander, M.; Ramírez, C.; Mendes, R.; Raaijmakers, J. Deciphering rhizosphere microbiome assembly of wild and modern common bean (Phaseolus vulgaris) in native and agricultural soils from Colombia. Microbiome 2019, 7, 114. [Google Scholar] [CrossRef] [Green Version]
- Xie, Y.; Jia, P.; Zheng, X.; Li, J.; Buwen, S.; Wang, Y. Effects and pathways of domestication on crop microbiome diversity and community structure. Chin. J. Plant Ecol. 2022, 46, 1–18. [Google Scholar] [CrossRef]
- Lee, J.; Teitzel, G.M.; Munkvold, K.; Del Pozo, O.; Martin, G.B.; Michelmore, R.W.; Greenberg, J.T. Type III secretion and effectors shape the survival and growth pattern of Pseudomonas syringae on leaf surfaces. Plant Physiol. 2012, 158, 1803–1818. [Google Scholar] [CrossRef] [Green Version]
- Dai, D.; Han, L.; Jin, X. Species Identification and Diversity Investigation of Bamboo Ascomycetes in Yunnan. J. Qujing Normal Univ. 2022, 41, 16–28. [Google Scholar]
- Ning, Y.; Li, Y.L.; Zhou, G.Y.; Yang, L.C.; Xu, W.H. Community composition and diversity of endophytic fungi from roots of Sinopodophyllum hexandrum in forest of Upper-north mountain of Qinghai province. China J. Chin. Mater. Med. 2016, 41, 1227–1234. [Google Scholar] [CrossRef]
- Deng, Z.S.; Liu, X.D.; Zhang, B.C.; Jiao, S.; Qi, X.Y.; Sun, Z.H. Root-Associated Endophytic Bacterial Community Composition of Pennisetum sinese from Four Representative Provinces in China. Microorganisms 2019, 7, 47. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Dong, C.B.; Chen, W.H.; Liang, J.D.; Han, Y.F.; Liang, Z.Q. Community composition and ecological functional structural analysis of endophytic fungi in bark of Eucommia ulmoides in different areas. China J. Chin. Mater. Med. 2019, 44, 1126–1134. [Google Scholar] [CrossRef]
- Li, Z.; PUBU, C.; Lyu, M.; Wang, M.; Liu, X. Species diversity of zygomycotan fungi in the Tibet Autonomous Region. Microbiol. China 2018, 45, 1250–1261. [Google Scholar] [CrossRef]
- Cai, X.B.; Peng, Y.L. Geographical distribution of arbuscular mycorrhizal fungi in the grassland ecosystems of the Tibetan plateau. Acta Ecol. Sin. 2016, 36, 2807–2818. [Google Scholar] [CrossRef]
- Zhu, G.L.; Li, J.; Wei, X.H. Longitudinal patterns of productivity and plant diversity in Tibetan alpine grasslands. J. Nat. Resour. 2017, 32, 210–222. [Google Scholar]
- Muhammad, F. Phylogeny of Alternaria spp. from Medicinal Plants in Southwest of China. Master’s Thesis, Guizhou University, Guiyang, China, 2021; pp. 77–82. [Google Scholar] [CrossRef]
- Yu, M.; Liu, Z.; Chen, S.; Chen, F. Expression of P. mume PGIP gene in transgenic Dendranthema morifolium increased tolerance to disease resistance. Acta Bot. Boreal. Occident. Sin. 2010, 30, 1111–1116. [Google Scholar]
- Yang, X.; He, X.; Chen, Q.; Zhang, J.; Fan, J.; Liu, P.; Hu, H. Screening of Pyrus pyrifolia genotypes for resistance to Alternaria alternata. Sci. Hortic. 2020, 259, 108838. [Google Scholar] [CrossRef]
- He, W.; Wang, H.; Zhang, Q.; Du, S.; Zhang, X.; Luo, B. Current Situation and Prospect of Research on Major Medicinal Plant Diseases in Sichuan. J. Chin. Med. Mater. 2022, 45, 2793. [Google Scholar] [CrossRef]
- Hervé, F.A.; Christian, S.; David, P.M.; Themis, J.M. Sensitivity and cross-resistance patterns to demethylation-inhibiting fungicides in California populations of Alternaria alternata pathogenic on pistachio. Crop Prot. 2016, 88, 72–78. [Google Scholar] [CrossRef]
- Yang, G.; Cui, S.; Ma, N.; Song, Y.; Ma, J.; Huang, W.; Zhang, Y.; Xua, J. Genetic Structure and Triazole Antifungal Susceptibilities of Alternaria alternata from Greenhouses in Kunming, China. Microbiol. Spectr. 2022, 10, e00382-22. [Google Scholar] [CrossRef]
- Ainsworth, G.C.; Sparrow, F.K.; Sussman, A.S. The Fungi: An Advanced Treatise. Volume IV A, a Taxonomic Review with Keys: Ascomycetes and Fungi Imperfecti; Academic Press: New York, NY, USA; London, UK, 1973; pp. 1–621. [Google Scholar] [CrossRef]
- Braun, U. A monograph of the Erysiphales (powdery mildews). Beih. Nova Hedwig. 1987, 89, 1–700. [Google Scholar]
- Amano, K. Host Range and Geographical Distribution of the Powdery Mildew Fungi, 2nd ed.; Taylor & Francis, Ltd.: Abingdon, UK, 1986; pp. 1–741. [Google Scholar] [CrossRef]
- An, Y.Y. Molecular Systematics of Mycosphaerellaceae Plant Pathogenic Fungi in Southwest China. Master’s Thesis, Guizhou University, Guiyang, China, 2021; pp. 87–89. [Google Scholar] [CrossRef]
- Braun, U.; Cook, R.T.A. Taxonomic Manual of the Erysiphales (Powdery Mildews); Fungal Biodiversity Centre, CBS Biodiversity Series No. 11; Centraalbureau voor Schimmelcultures: Utrecht, The Netherlands, 2012; pp. 1–707. [Google Scholar]
- Pourhosseini, L.; Kermani, M.J.; Habashi, A.A.; Khalighi, A. Efficiency of direct and indirect shoot organogenesis in different genotypes of Rosa hybrida. Plant Cell Tissue Organ Cult. 2013, 112, 101–108. [Google Scholar] [CrossRef]
- Kiss, L.; Jankovics, T.; Kovács, G.M.; Daughtrey, M.L. Oidium longipes, a new powdery mildew fungus on petunia in the USA: A potential threat to ornamental and vegetable solanaceous crops. Plant Dis. 2008, 92, 818–825. [Google Scholar] [CrossRef] [Green Version]
- Videira, S.; Groenewald, J.; Nakashima, C.; Braun, U.; Barreto, R.; de Wit, P.; Crous, P. Mycosphaerellaceae—Chaos or clarity? Stud. Mycol. 2017, 87, 257–421. [Google Scholar] [CrossRef]
- Crous, P.W.; Kang, J.C.; Braun, U. A phylogenetic redefifinition of anamorph genera in Mycosphaerella based on ITS rDNA sequence and morpholo. Mycologia 2001, 93, 1081–1101. [Google Scholar] [CrossRef]
- Murtza, T.; You, M.P.; Barbetti, M.J. Synergistic/antagonistic interactions between Neopseudocercosporella, Alternaria, Leptosphaeria, and Hyaloperonospora determine aggregate foliar disease severity in rapeseed. Plant Pathol. 2022, 71, 523–534. [Google Scholar] [CrossRef]
- Miguel, A.M.; Tino, K. The effect of bacterial chemotaxis on host infection and pathogenicity. FEMS Microbiol. Rev. 2018, 42, 40–67. [Google Scholar] [CrossRef] [Green Version]
- Walley, J.W.; Kliebenstein, D.J.; Bostock, R.M.; Dehesh, K. Fatty acids and early detection of pathogens. Curr. Opin. Plant Biol. 2013, 16, 520–526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.; Chin, C.; Chen, A. Expression of the yeast delta-9 desaturase gene in tomato enhances its resistance to powdery mildew. Physiol. Mol. Plant Pathol. 1998, 52, 371–383. [Google Scholar] [CrossRef]
- Xing, J.; Chin, C.K. Modification of fatty acids in eggplant affects its resistance to Verticillium dahliae. Physiol. Mol. Plant Pathol. 2000, 56, 217–225. [Google Scholar] [CrossRef]
- Madi, L.; Wang, X.; Kobiler, I.; Lichter, A.; Prusky, D. Stress on avocado fruits regulates 9-stearoyl ACP desaturase expression, fatty acid composition, antifungal diene level and resistance to Colletotrichum gloeosporioides attack. Physiol. Mol. Plant Pathol. 2003, 62, 277–283. [Google Scholar] [CrossRef]
- Yaeno, T.; Matsuda, O.; Iba, K. Role of chloroplast trienoic fatty acids in plant disease defense responses:Chloroplast TAs in defense responses. Plant J. 2004, 40, 931–941. [Google Scholar] [CrossRef]
- Ongena, M.; Duby, F.; Rossignol, F.; Fauconnier, M.L.; Dommes, J.; Thonart, P. Stimulation of the lipox ygenase pathway is associated with systemic resistance induced in bean by a nonpathogenic Pseudomonas strain. Mol. Plant-Microb. Interact. 2004, 17, 1009–1018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rita, Z.; Mark, S.; Uwe, S.; Ralf, B. Pyrimidine and Purine Biosynthesis and Degradation in Plants. Plant Biol 2006, 57, 805–836. [Google Scholar] [CrossRef]
- Abi, S.G.; Huang, S.; Monika, W.M. The peptidases involved in plant mitochondrial protein import. J. Exp. Bot. 2019, 70, 6005–6018. [Google Scholar] [CrossRef]
- Feng, Y.L.; Lei, Y.B.; Wang, R.F.; Callaway, R.M.; Valiente-Banuet, A.; Inderjit; Li, Y.P.; Zheng, Y.L. Evolutionary tradeoffs for nitrogen allocation to photosynthesis versus cell walls in an invasive plant. Proc. Natl. Acad. Sci. USA 2009, 106, 1853–1856. [Google Scholar] [CrossRef] [Green Version]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, N.-N.; Zeng, Z.-Y.; Xu, Q.-B.; Zhang, H.-B.; Xu, T. Artificial Cultivation Changes Foliar Endophytic Fungal Community of the Ornamental Plant Lirianthe delavayi. Microorganisms 2023, 11, 775. https://doi.org/10.3390/microorganisms11030775
Wu N-N, Zeng Z-Y, Xu Q-B, Zhang H-B, Xu T. Artificial Cultivation Changes Foliar Endophytic Fungal Community of the Ornamental Plant Lirianthe delavayi. Microorganisms. 2023; 11(3):775. https://doi.org/10.3390/microorganisms11030775
Chicago/Turabian StyleWu, Nan-Nan, Zhao-Ying Zeng, Qin-Bin Xu, Han-Bo Zhang, and Tao Xu. 2023. "Artificial Cultivation Changes Foliar Endophytic Fungal Community of the Ornamental Plant Lirianthe delavayi" Microorganisms 11, no. 3: 775. https://doi.org/10.3390/microorganisms11030775