
Microbial Composition on Abandoned and Reclaimed Mining Sites in the Komi Republic (North Russia)
 , ,
, ,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling
2.2. Agrochemical Analysis and DNA Extraction
2.3. Bioinformatic Analysis
3. Results
3.1. General Soil Characteristics
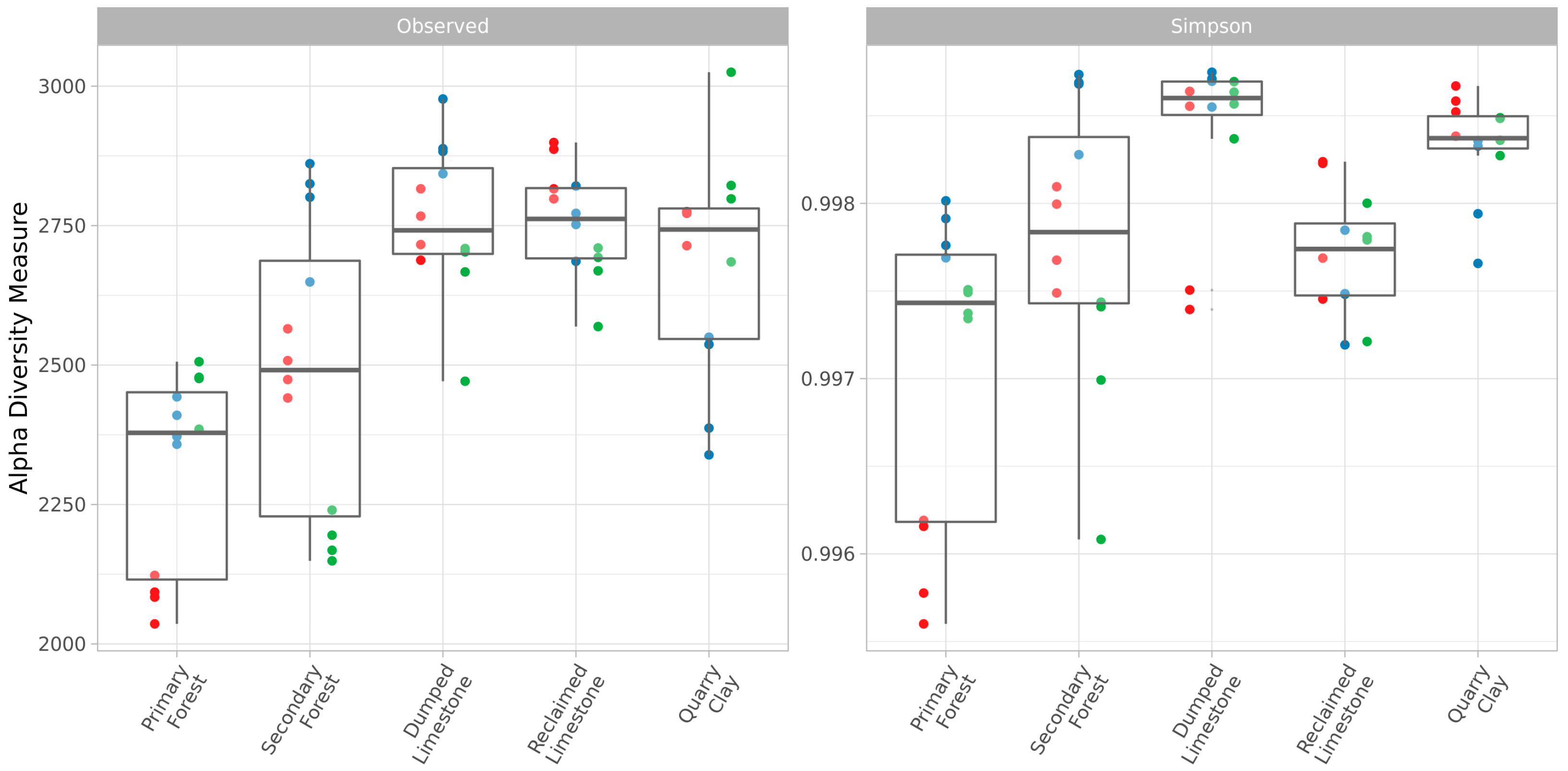
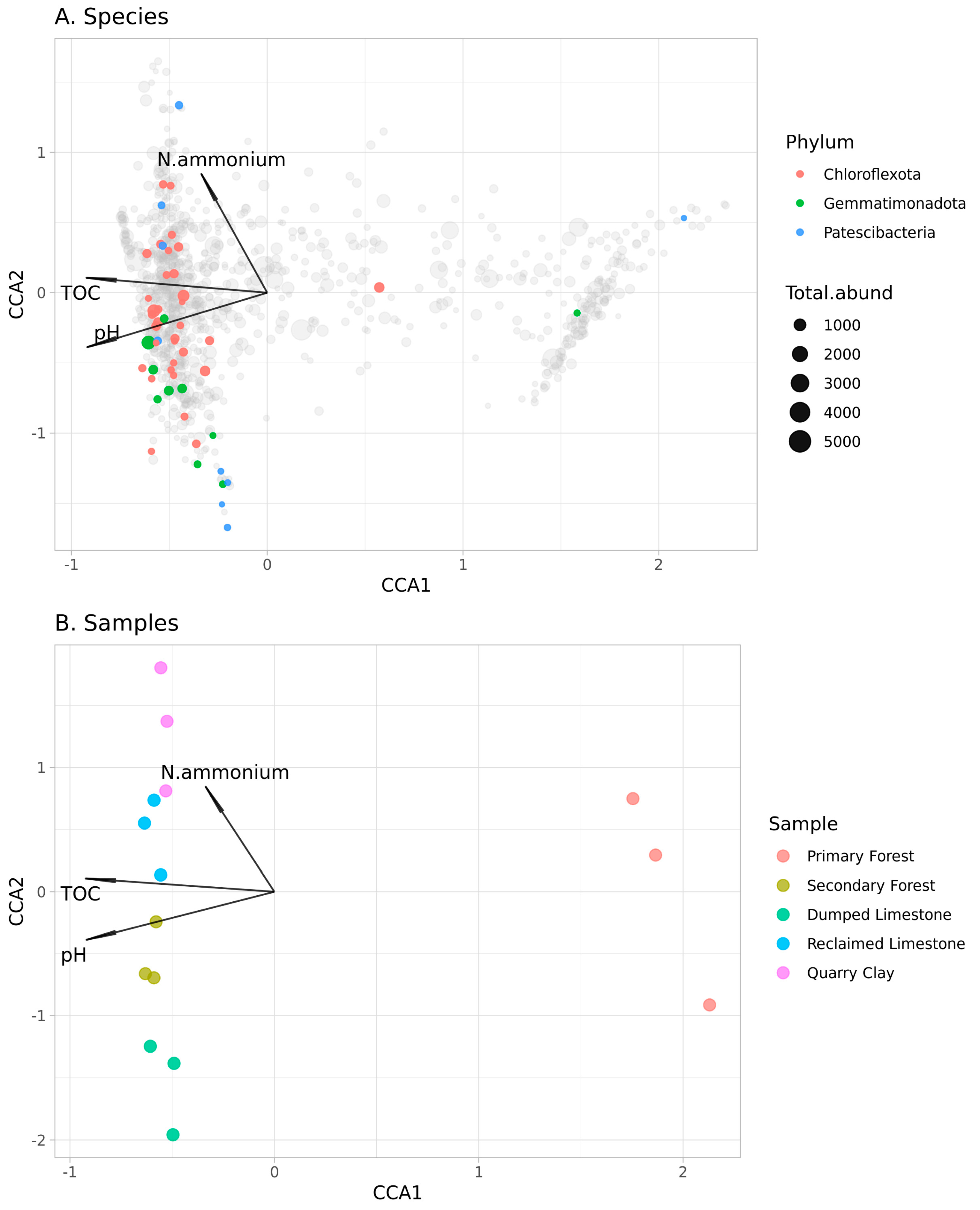
3.2. Soil Microbiota
4. Discussion
4.1. Primary Forest Soil Was Distinct from All Other Samples
4.2. Secondary Forest Samples Were More Diverse Than the Primary Ones
4.3. Abandoned Quarries Were Poor, but Different, Due to Their Soil Properties
4.4. Limestone Recultivated Soils Had Their Unique Microbiome
4.5. pH, TOC and Ammonium Concentration Were Significant Abiotic Factors
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Evgeny, L. Soil Atlas of the Komi Republic; LC “Komi Republician Publishing House”: Syktyvkar, Russia, 2010. [Google Scholar]
- Iliushin, V.; Kirtsideli, I. Dynamics of complexes of microfungi in the process of overgrowing spoil tips of coal mines in the southern tundra zone (komi republic). Micol. Phytopatology 2021, 55, 129–137. [Google Scholar] [CrossRef]
- Pengerud, A.; Dignac, M.-F.; Certini, G.; Strand, L.; Forte, C.; Rasse, D. Soil Organic Matter Molecular Composition and State of Decomposition in Three Locations of the European Arctic. Biogeochemistry 2017, 135, 277–292. [Google Scholar] [CrossRef]
- Pastukhov, A.; Marchenko-Vagapova, T.; Loiko, S.; Kaverin, D. Vulnerability of the Ancient Peat Plateaus in Western Siberia. Plants 2021, 10, 2813. [Google Scholar] [CrossRef]
- Walker, T.; Young, S.; Crittenden, P.; Zhang, H. Anthropogenic Metal Enrichment of Snow and Soil in North-Eastern European Russia. Environ. Pollut. Barking Essex 1987 2003, 121, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Zemlyanskiy, V.; Kurta, I.; Pasynkov, A. Technological Researches of Coal Mining Waste with Its Processing and Utilization to Build-up Production of Constructional Concrete in the North. In Proceedings of the XVIII International Coal Preparation Congress, Saint-Petersburg, Russia, 28 June–1 July 2016; pp. 477–482, ISBN 978-3-319-40942-9. [Google Scholar]
- Sheoran, V.; Sheoran, A.; Poonia, P. Soil Reclamation of Abandoned Mine Land by Revegetation: A Review. Int. J. Soil Sediment Water 2010, 3, 13. [Google Scholar]
- Zverev, A.; Kimeklis, A.; Kichko, A.; Gladkov, G.; Andronov, E.; Abakumov, E. Microbial Features of Mature and Abandoned Soils in Refractory Clay Deposits. BMC Microbiol. 2022, 22, 237. [Google Scholar] [CrossRef] [PubMed]
- Abakumov, E.; Maksimova, E.; Lagoda, A.; Koptseva, E. Soil Formation in the Quarries for Limestone and Clay Production in the Ukhta Region. Eurasian Soil Sci. 2011, 44, 380–385. [Google Scholar] [CrossRef]
- Archegova, I. Thermal Regime of Tundra Soils under Reclamation and Restoration of Natural Vegetation. Eurasian Soil Sci. 2007, 40, 854–859. [Google Scholar] [CrossRef]
- Likhanova, I.; Archegova, I. Soil Formation under Progressive Accelerated Succession of Forest Ecosystems in the North of Russia. Contemp. Probl. Ecol. 2009, 2, 66–71. [Google Scholar] [CrossRef]
- Pershina, E.; Ivanova, E.; Kineklis, A.; Zverev, A.; Kichko, A.; Aksenova, T.; Evgeny, A.; Abakumov, E. Soil Microbiomes of Reclaimed and Abandoned Mines of the Yamal Region. Pol. Polar Res. 2020, 41, 95–114. [Google Scholar] [CrossRef]
- Abakumov, E.; Zverev, A.; Morgun, E.; Alekseev, I. Microbiome of Abandoned Agricultural and Mature Tundra Soils in Southern Yamal Region, Russian Arctic. Open Agric. 2020, 5, 335–344. [Google Scholar] [CrossRef]
- Zverev, A.; Petrov, A.; Kimeklis, A.; Kichko, A.; Andronov, E.; Petrov, A.; Abakumov, E. Microbiomes of the Initial Soils of Mining Areas of Yakutsk City (Eastern Siberia, Russia). Czech Polar Rep. 2020, 10, 69–82. [Google Scholar] [CrossRef]
- World Reference Base for Soil Resources. International Soil Classification System for Naming Soils and Creating Legends for Soil Maps, 4th ed.; International Union of Soil Sciences: Vienna, Austria, 2022. [Google Scholar]
- Walkley, A.; Black, I.A. An Examination of the Degtjareff Method for Determining Soil Organic Matter, and A Proposed Modification of the Chromic Acid Titration Method. Soil Sci. 1934, 37, 29–38. [Google Scholar] [CrossRef]
- FAQ. Standard Operating Procedure for Soil PH Determination; FAQ: Rome, Italy, 2021. [Google Scholar]
- FAQ. Standard Operating Procedure for Soil Available Phosphorus-Olsen Method; FAQ: Rome, Italy, 2021. [Google Scholar]
- Caporaso, J.; Lauber, C.; Walters, W.; Berg-Lyons, D.; Lozupone, C.; Turnbaugh, P.; Fierer, N.; Knight, R. Global Patterns of 16S RRNA Diversity at a Depth of Millions of Sequences per Sample. Proc. Natl. Acad. Sci. USA 2011, 108 (Suppl. S1), 4516–4522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Callahan, B.; Mcmurdie, P.; Rosen, M.; Han, A.; Johnson, A.J.; Holmes, S. DADA2: High-Resolution Sample Inference from Illumina Amplicon Data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [Green Version]
- Mcmurdie, P.; Holmes, S. Phyloseq: An R Package for Reproducible Interactive Analysis and Graphics of Microbiome Census Data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef] [Green Version]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F. The SILVA Ribosomal RNA Gene Database Project: Improved Data Processing and Web-Based Tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef]
- Parte, A.C.; Sardà Carbasse, J.; Meier-Kolthoff, J.P.; Reimer, L.C.; Göker, M. List of Prokaryotic Names with Standing in Nomenclature (LPSN) Moves to the DSMZ. Int. J. Syst. Evol. Microbiol. 2020, 70, 5607–5612. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.; O’Hara, B.; Simpson, G.; Solymos, P.; Stevens, H.; Wagner, H. Vegan: Community Ecology Package. R Package Version 22-1 2015, 2, 1–2. [Google Scholar]
- Love, M.; Huber, W.; Anders, S. Moderated Estimation of Fold Change and Dispersion for RNA-Seq Data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, Z.; Zhang, Y.; Yang, C.; Wang, S. Land-Use Type Strongly Shapes Community Composition, but Not Always Diversity of Soil Microbes in Tropical China. Catena 2018, 165, 369–380. [Google Scholar] [CrossRef]
- Cho, H.; Ahn, J.-H.; Weon, H.-Y.; Joa, J.-H.; Hong, S.-B.; Seok, S.-J.; Kim, J.-S.; Kwon, S.-W.; Kim, S.-J. Parafilimonas Rhizosphaerae Sp. Nov., Isolated from the Rhizosphere of Tomato Plant (Solanum lycopersicum L.). Int. J. Syst. Evol. Microbiol. 2017, 67, 2279–2283. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.-J.; Park, J.-H.; Lim, J.-M.; Ahn, J.-H.; Anandham, R.; Weon, H.-Y.; Kwon, S.-W. Parafilimonas Terrae Gen. Nov., Sp Nov., Isolated from Greenhouse Soil. Int. J. Syst. Evol. Microbiol. 2014, 64, 3040–3045. [Google Scholar] [CrossRef]
- Methé, B.A.; Nelson, K.E.; Eisen, J.A.; Paulsen, I.T.; Nelson, W.; Heidelberg, J.F.; Wu, D.; Wu, M.; Ward, N.; Beanan, M.J.; et al. Genome of Geobacter Sulfurreducens: Metal Reduction in Subsurface Environments. Science 2003, 302, 1967–1969. [Google Scholar] [CrossRef] [Green Version]
- Lv, Y.; Gao, Z.; Xia, F.; Maman, A.; Qiu, L. Puia Dinghuensis Gen. Nov., Sp. Nov., Isolated from Monsoon Evergreen Broad-Leaved Forest Soil. Int. J. Syst. Evol. Microbiol. 2017, 67, 4639–4645. [Google Scholar] [CrossRef] [PubMed]
- Spyridonov, I.; Krafft, L.; Schöning, I.; Schrumpf, M.; Nacke, H. The Ubiquitous Soil Verrucomicrobial Clade ‘Candidatus Udaeobacter’ Shows Preferences for Acidic PH. Environ. Microbiol. Rep. 2021, 13, 878–883. [Google Scholar] [CrossRef]
- Liang, Y.; Fujing, P.; Ma, J.; Yang, Z.; Yan, P. Long-Term Forest Restoration Influences Succession Patterns of Soil Bacterial Communities. Environ. Sci. Pollut. Res. 2021, 28, 20598–20607. [Google Scholar] [CrossRef] [PubMed]
- Akter, S.; Park, J.-H.; Rahman, M.M.; Huq, M.A. Niastella Soli Sp. Nov., Isolated from Rhizospheric Soil of a Persimmon Tree. Int. J. Syst. Evol. Microbiol. 2021, 71, 004870. [Google Scholar] [CrossRef]
- Guo, Q.; Zhou, Z.; Zhang, L.; Zhang, C.; Chen, M.; Wang, B.; Lin, M.; Wang, W.; Zhang, W.; Li, X. Skermanella Pratensis Sp. Nov., Isolated from Meadow Soil, and Emended Description of the Genus Skermanella. Int. J. Syst. Evol. Microbiol. 2020, 70, 1605–1609. [Google Scholar] [CrossRef]
- Carvalho, F.M.; Souza, R.C.; Barcellos, F.G.; Hungria, M.; Vasconcelos, A.T.R. Genomic and Evolutionary Comparisons of Diazotrophic and Pathogenic Bacteria of the Order Rhizobiales. BMC Microbiol. 2010, 10, 37. [Google Scholar] [CrossRef] [Green Version]
- DeBruyn, J.M.; Nixon, L.T.; Fawaz, M.N.; Johnson, A.M.; Radosevich, M. Global Biogeography and Quantitative Seasonal Dynamics of Gemmatimonadetes in Soil. Appl. Environ. Microbiol. 2011, 77, 6295–6300. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Vegetation | Replica Details | Replica | Soil Profile | Location Details | Location |
|---|---|---|---|---|---|
| Clover, coltsfoot, cereals, St. John’s wort, moss | Bottom of the quarry uphill from water | I | W * C | Abandoned clay quarry near a road. Herbs overgrown on the bottom of the quarry | Quarry Clay N 63.54852, E 53.757625 |
| Willow, sparse herbs, litter | Bottom of the quarry near the slope | II | |||
| Grass cover, dotted spruce and pine | Bottom of the quarry near the water pond | III | |||
| Pine, birch, rowan, moss, blueberry, wild rose, lingonberry, oxalis, legumes | Forest, forest litter | I | AY * E BF C | Primary pine forest with well-developed underwood | Primary Forest N 63.542693, E 53.776319 |
| Pine, birch, club moss | Forest, forest litter and moss | II | |||
| Birch, rowan | Forest, forest litter and grass | III | |||
| Sedge, cereals, coltsfoot, legumes | Bottom of the quarry | I | W * AC Cc | Bottom of abandoned limestone quarry, broken stones with herbs | Dumped Limestone N 63.599297, E 53.776415 |
| Birch, pine, willow, spruce, coltsfoot, Ivan-tea, crushed stone on the surface, moss | Bottom of the quarry, young forest | II | |||
| Burdock, cereals, legumes, coltsfoot | Top of the local hill in the quarry | III | |||
| Willow, pine, birch, cereals, legumes, Ivan-tea | Slope of dump | I | W * ACca Cg | Bottom of limestone quarry recultivated by topsoil cover, broken stones | Reclaimed Limestone N 63.589972, E 53.796547 |
| Ivan-tea, moss, peas, willow, pine, birch | Slope with rocks | II | |||
| Aspen, undergrowth of larch and pine, birch | Edge of quarry | III | |||
| Undergrowth of pine, birch, willow. herbs, coltsfoot, clover, A moss, legumes, Ivan-tea | Edge of the forest | I | AU * AC Cca | Young secondary forest (pine, birch, willow) | Secondary Forest N 63.596806, E 53.782191 |
| Birch, pine, moss, clover, legumes, dandelion, coltsfoot | Forest | II | |||
| Pine, birch, willow, clover, legumes, Ivan-tea | Forest | III |
| NO3− mg/100 g | NH4+ mg/100 g | K, mg/100 g | P, mg/100 g | pH | TOC % | Replica | Location |
|---|---|---|---|---|---|---|---|
| 15.2 | 98.6 | 369 | 42 | 6.8 | 1.45 | I | Quarry Clay |
| 1.59 | 44.3 | 337 | 7.3 | 7 | 1.56 | II | |
| 12.4 | 93 | 885 | 36 | 6.9 | 1.21 | III | |
| 2.5 | 7.62 | 59 | 46 | 6.4 | 0.66 | I | Primary Forest |
| 2.24 | 10.3 | 188 | 52 | 5.9 | 0.87 | II | |
| 0.77 | 19.2 | 296 | 67 | 5.5 | 0.55 | III | |
| 0.73 | 3.76 | 785 | 4.7 | 7.3 | 1.22 | I | Dumped Limestone |
| 1.38 | 9.36 | 118 | 6.8 | 7.5 | 1.34 | II | |
| 4.65 | 4.26 | 240 | 2.1 | 7.4 | 1.12 | III | |
| 10.6 | 13.4 | 205 | 3.4 | 7.3 | 1.45 | I | Secondary Forest |
| 1.08 | 17.5 | 188 | 1.3 | 7.2 | 1.65 | II | |
| 1.08 | 15.7 | 181 | 3.4 | 7.2 | 1.45 | III | |
| 8.52 | 19 | 77 | 0.9 | 7.2 | 1.65 | I | Reclaimed Limestone |
| 7.86 | 44.3 | 146 | 4.7 | 7.1 | 1.66 | II | |
| 5.24 | 37.9 | 216 | 3.4 | 7.1 | 1.7 | III |
| Quarry Clay | Reclaimed Limestone | Dumped Limestone | Secondary Forest | Primary Forest | |
|---|---|---|---|---|---|
| <0.01 | <0.01 | <0.01 | 0.137 | Primary Forest | |
| 0.086 | 0.06 | 0.05 | 0.034 | Secondary Forest | |
| 0.822 | 1 | 0.094 | <00.1 | Dumped Limestone | |
| 0.852 | 0.038 | 0.995 | 0.087 | Reclaimed Limestone | |
| 0.086 | 0.997 | 0.193 | <0.01 | Quarry Clay |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zverev, A.O.; Gladkov, G.V.; Kimeklis, A.K.; Kichko, A.A.; Andronov, E.E.; Abakumov, E.V. Microbial Composition on Abandoned and Reclaimed Mining Sites in the Komi Republic (North Russia). Microorganisms 2023, 11, 720. https://doi.org/10.3390/microorganisms11030720
Zverev AO, Gladkov GV, Kimeklis AK, Kichko AA, Andronov EE, Abakumov EV. Microbial Composition on Abandoned and Reclaimed Mining Sites in the Komi Republic (North Russia). Microorganisms. 2023; 11(3):720. https://doi.org/10.3390/microorganisms11030720
Chicago/Turabian StyleZverev, Aleksei O., Grigory V. Gladkov, Anastasiia K. Kimeklis, Arina A. Kichko, Evgeny E. Andronov, and Evgeny V. Abakumov. 2023. "Microbial Composition on Abandoned and Reclaimed Mining Sites in the Komi Republic (North Russia)" Microorganisms 11, no. 3: 720. https://doi.org/10.3390/microorganisms11030720