

Succession Patterns of Microbial Composition and Activity following the Diesel Spill in an Urban River

{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Water Sampling Procedure
2.2. Microcosm Setup
2.3. Hydrocarbon Extraction and Analysis
2.4. Microbial Biomass Production
2.5. Microbial 14C-hydrocarbon Oxidation Rates
2.6. DNA Extraction and Cell Counting
2.7. Illumina Miseq Sequencing of 16S rRNA Gene Amplicons and Data Analysis
2.8. Statistical Analyses
3. Result
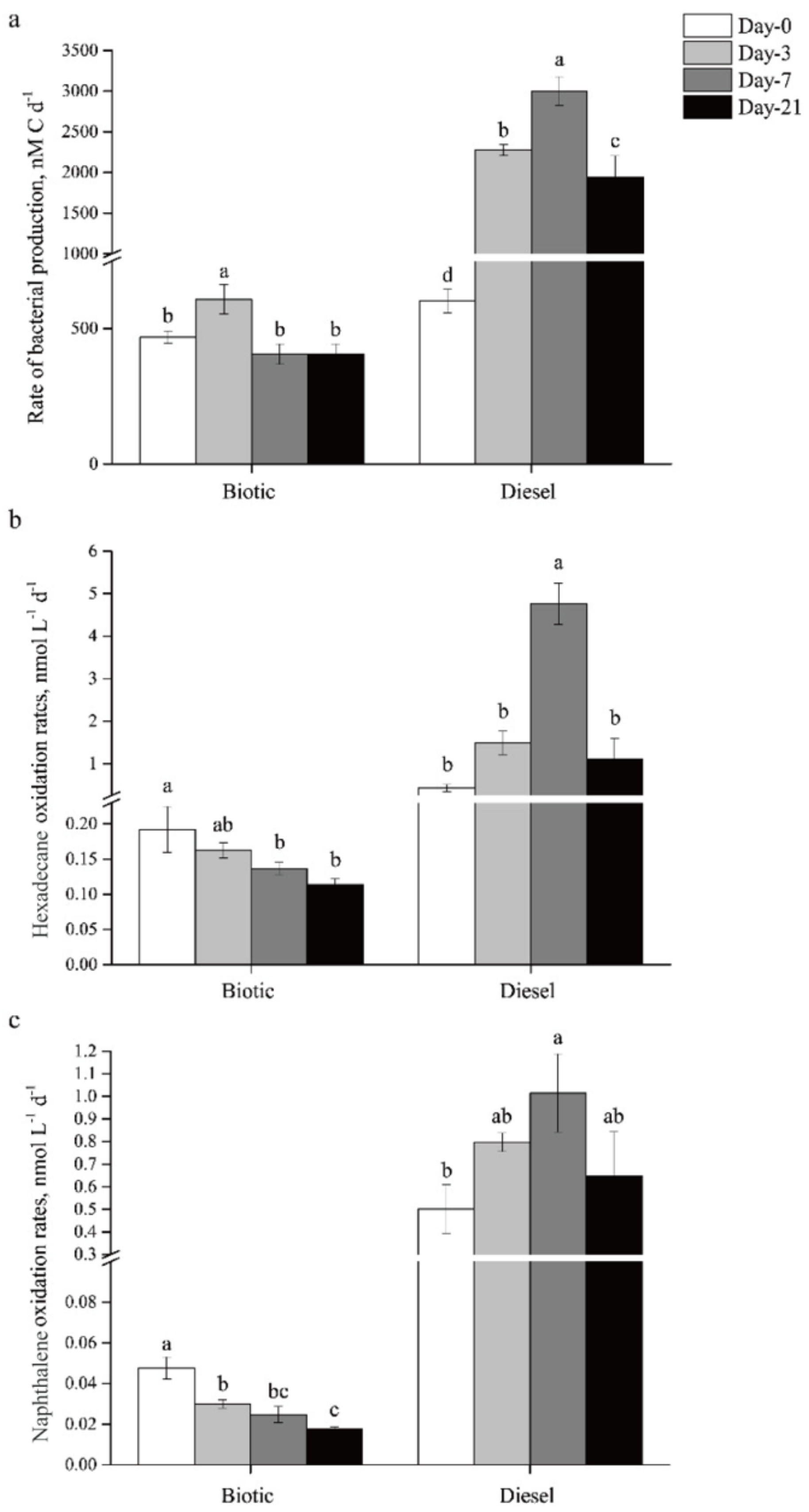
3.1. Response of Microbial Total Activities to Diesel Addition
3.2. Response of Oil-Degrading Microorganisms to Diesel Oil Addition
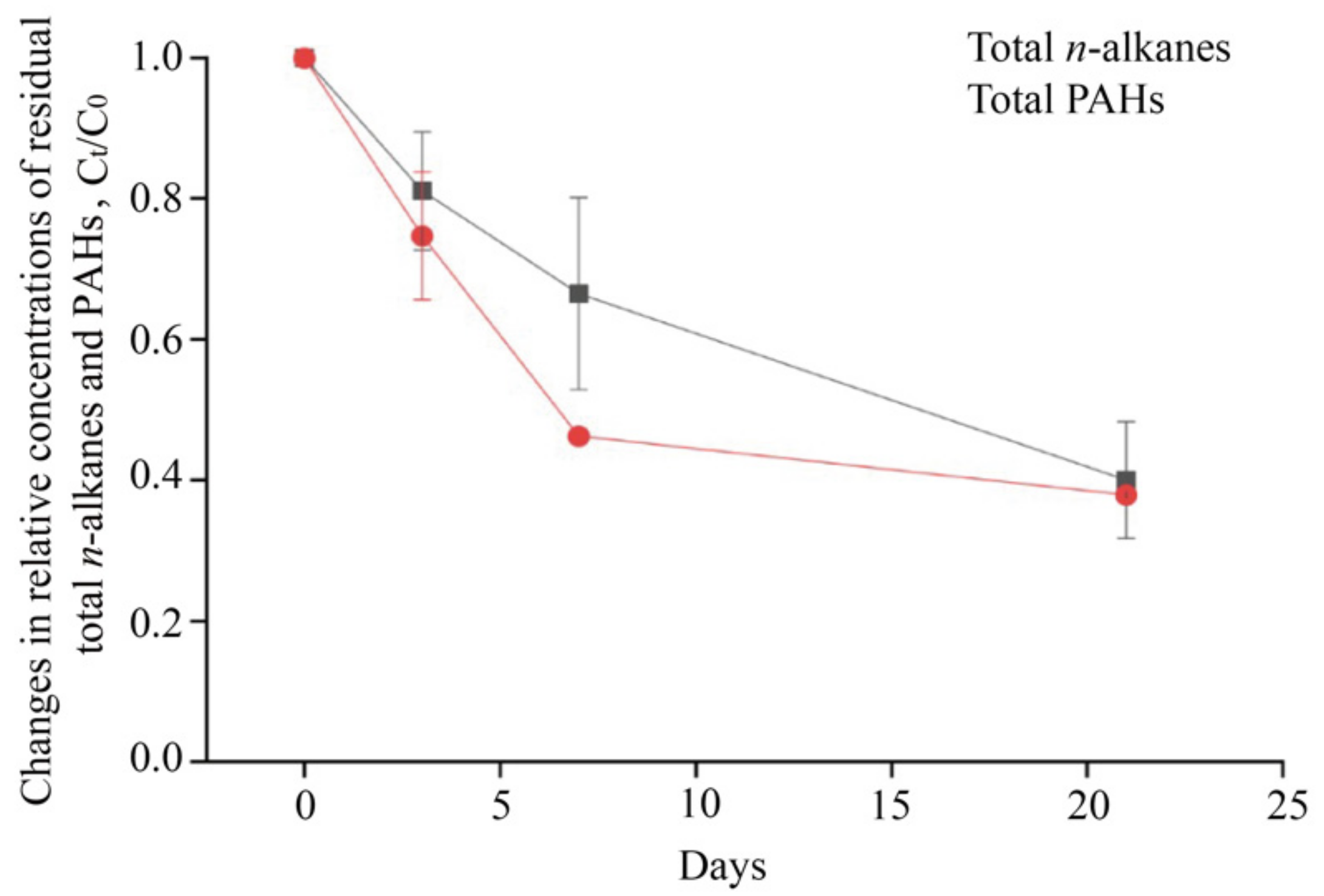
3.3. The Degradation of Total n-Alkanes and PAHs during the Incubation
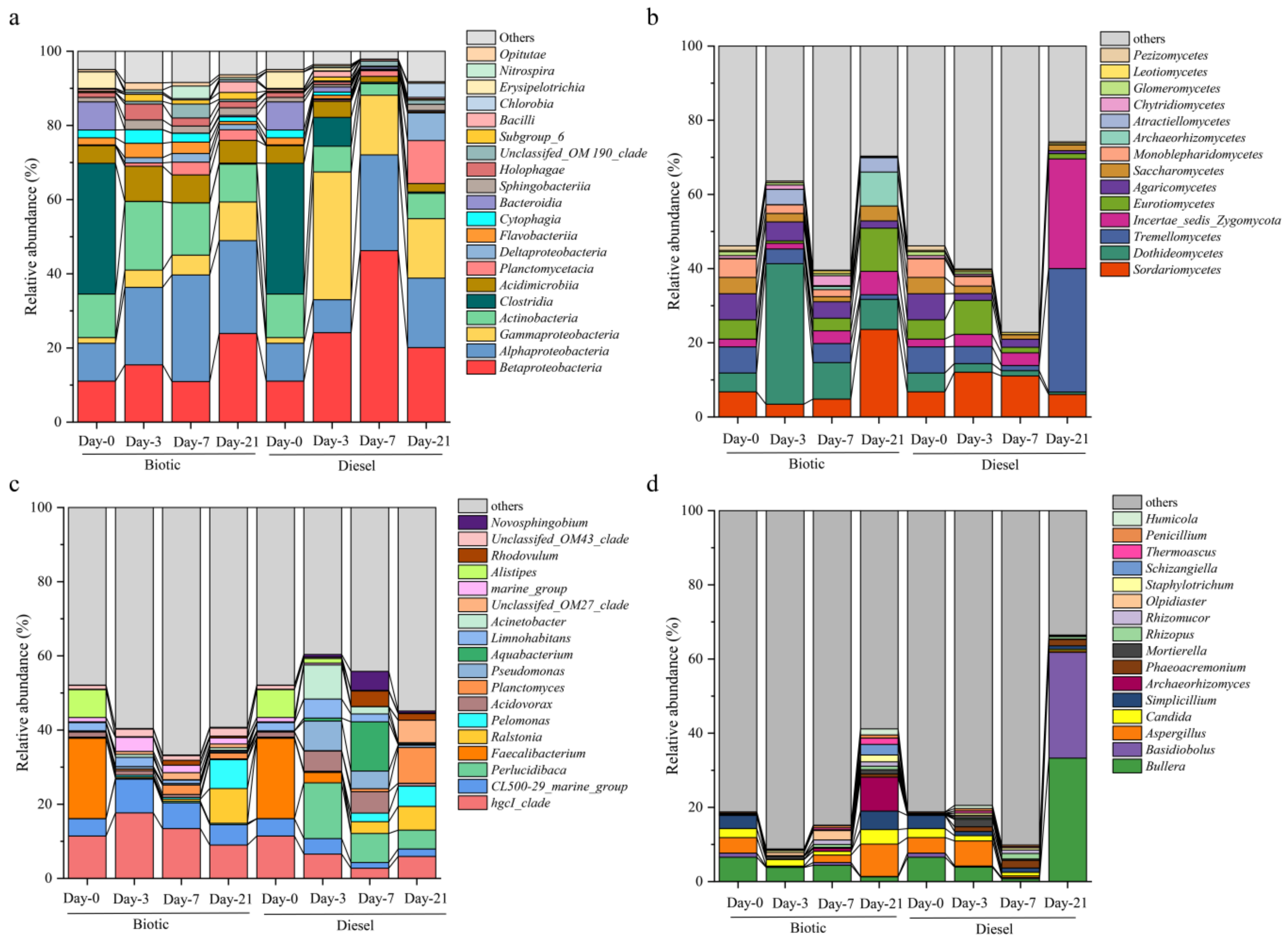
3.4. Shift in the Microbial Community Composition and Key Potential Oil-Degrading Bacteria during the Incubation
4. Discussion
4.1. Fast Responses of Microbial Activities and Diesel-Degrading Microorganisms to Diesel in the River Ecosystem
4.2. The Enrichment and Succession of Diesel-Degrading Bacteria and Fungi during the Incubation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Chang, S.E.; Stone, J.; Demes, K.; Piscitelli, M. Consequences of oil spills: A review and framework for informing planning. Ecol. Soc. 2014, 19, 26. [Google Scholar] [CrossRef] [Green Version]
- Michel, J.; Fingas, M. Oil Spills: Causes, consequences, prevention, and countermeasures. In Fossil Fuels: Current Status and Future Directions; Crawley, G.M., Ed.; World Scientific: Singapore, 2016; pp. 159–201. [Google Scholar]
- Jernelöv, A. The threats from oil spills: Now, then, and in the future. Ambio 2010, 39, 353–366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khalid, F.E.; Lim, Z.S.; Sabri, S.; Gomez-Fuentes, C.; Zulkharnain, A.; Ahmad, S.A. Bioremediation of diesel contaminated marine water by bacteria: A review and bibliometric analysis. J. Mar. Sci. Eng. 2021, 9, 155. [Google Scholar] [CrossRef]
- Duodu, G.O.; Ogogo, K.N.; Mummullage, S.; Harden, F.; Goonetilleke, A.; Ayoko, G.A. Source apportionment and risk assessment of PAHs in Brisbane river sediment, Australia. Ecol. Indic. 2017, 73, 784–799. [Google Scholar] [CrossRef]
- Shangyu, L.; Jian, D. Risk Assessment on Ship’s Oil Spill of Yangtze River Main Line Wharf. In IOP Conference Series: Earth and Environmental Science; IOP Publishing: London, UK, 2020; p. 012074. [Google Scholar]
- Bezmaternykh, D.; Puzanov, A.; Kotovshchikov, A.; Drobotov, A.; Tolomeev, A. Hydrochemical Indicators of Water Quality in the Norilsk–Pyasino Lake–River System after a Diesel Fuel Spill at Norilsk Heat and Power Plant 3 in 2020. Contemp. Probl. Ecol. 2021, 14, 323–334. [Google Scholar] [CrossRef]
- Glyaznetsova, Y.S.; Nemirovskaya, I.A.; Flint, M.V. Study of the Effects of an accidental diesel fuel spill in Norilsk. Dokl. Earth Sci. 2021, 501, 994–999. [Google Scholar] [CrossRef]
- Mapelli, F.; Scoma, A.; Michoud, G.; Aulenta, F.; Boon, N.; Borin, S.; Kalogerakis, N.; Daffonchio, D. Biotechnologies for marine oil spill cleanup: Indissoluble ties with microorganisms. Trends Biotechnol. 2017, 35, 860–870. [Google Scholar] [CrossRef]
- Head, I.M.; Jones, D.M.; Röling, W.F.M. Marine microorganisms make a meal of oil. Nat. Rev. Microbiol. 2006, 4, 173–182. [Google Scholar] [CrossRef]
- Dombrowski, N.; Donaho, J.A.; Gutierrez, T.; Seitz, K.W.; Teske, A.P.; Baker, B.J. Reconstructing metabolic pathways of hydrocarbon-degrading bacteria from the Deepwater Horizon oil spill. Nat. Microbiol. 2016, 1, 16057. [Google Scholar] [CrossRef]
- Varjani, S.J.; Upasani, V.N. A new look on factors affecting microbial degradation of petroleum hydrocarbon pollutants. Int. Biodeterior. Biodegrad. 2017, 120, 71–83. [Google Scholar] [CrossRef]
- Nzila, A. Update on the cometabolism of organic pollutants by bacteria. Environ. Pollut. 2013, 178, 474–482. [Google Scholar] [CrossRef] [PubMed]
- Hu, P.; Dubinsky, E.A.; Probst, A.J.; Wang, J.; Sieber, C.M.K.; Tom, L.M.; Gardinali, P.R.; Banfield, J.F.; Atlas, R.M.; Andersen, G.L. Simulation of Deepwater Horizon oil plume reveals substrate specialization within a complex community of hydrocarbon degraders. Proc. Natl. Acad. Sci. USA 2017, 114, 7432–7437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Onur, G.; Yilmaz, F.; Icgen, B. Diesel oil degradation potential of a bacterium inhabiting petroleum hydrocarbon contaminated surface waters and characterization of its emulsification ability. J. Surfactants Deterg. 2015, 18, 707–717. [Google Scholar] [CrossRef]
- Kayode-Isola, T.; Eniola, K.; Olayemi, A.; Igunnugbemi, O. Response of resident bacteria of a crude oil-polluted river to diesel oil. Eur. J. Agron. 2008, 1, 6–9. [Google Scholar]
- Icgen, B.; Yilmaz, F. Screening and in situ monitoring of potential petroleum hydrocarbon degraders in contaminated Surface water. CLEAN 2017, 45, 1. [Google Scholar] [CrossRef]
- Sreejina, K.; Sruthi, T.; Vineeth, T. Diesel oil utilization efficiency of selective bacterial isolates from automobile workshop and Thesjaswini river of Kerala. Pollution 2016, 2, 221–232. [Google Scholar] [CrossRef]
- Khan, A.H.A.; Tanveer, S.; Anees, M.; Muhammad, Y.S.; Iqbal, M.; Yousaf, S. Role of nutrients and illuminance in predicting the fate of fungal mediated petroleum hydrocarbon degradation and biomass production. J. Environ. Manag. 2016, 176, 54–60. [Google Scholar] [CrossRef]
- Harms, H.; Schlosser, D.; Wick, L.Y. Untapped potential: Exploiting fungi in bioremediation of hazardous chemicals. Nat. Rev. Microbiol. 2011, 9, 177–192. [Google Scholar] [CrossRef]
- Peng, C.; Tang, Y.; Yang, H.; He, Y.; Liu, Y.; Liu, D.; Qian, Y.; Lu, L. Time- and compound-dependent microbial community compositions and oil hydrocarbon degrading activities in seawater near the Chinese Zhoushan Archipelago. Mar. Pollut. Bull. 2020, 152, 110907. [Google Scholar] [CrossRef]
- Song, B.; Li, Z.; Li, S.; Zhang, Z.; Fu, Q.; Wang, S.; Li, L.; Qi, S. Functional metagenomic and enrichment metatranscriptomic analysis of marine microbial activities within a marine oil spill area. Environ. Pollut. 2021, 274, 116555. [Google Scholar] [CrossRef]
- Ribicic, D.; Netzer, R.; Hazen, T.C.; Techtmann, S.M.; Drabløs, F.; Brakstad, O.G. Microbial community and metagenome dynamics during biodegradation of dispersed oil reveals potential key-players in cold Norwegian seawater. Mar. Pollut. Bull. 2018, 129, 370–378. [Google Scholar] [CrossRef] [Green Version]
- Xu, F.; Jia, Y.; Wang, Y.; Zhang, F.; Li, L.; Li, Y.; Ren, L.; Wang, D.; Zhang, T. Does sand mining affect the remobilization of copper and zinc in sediments?—A case study of the Jialing River (China). Environ. Res. 2021, 200, 111416. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Chang, S.; Wang, X.; Wang, J.; Zhang, Z.; Pan, C.; Guo, Q. Emergency treatment technology and case study of sudden environmental pollution caused by oil spill in river basin. In IOP Conference Series: Earth and Environmental Science; IOP Publishing: London, UK, 2021; Volume 621, p. 012143. [Google Scholar]
- Liu, J.; Bacosa, H.P.; Liu, Z. Potential environmental factors affecting oil-degrading bacterial populations in deep and surface waters of the northern gulf of mexico. Front. Microbiol. 2016, 7, 2131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Zhang, J.; Li, H.; Yang, H.; Peng, C.; Peng, Z.; Lu, L. Shift in the microbial community composition of surface water and sediment along an urban river. Sci. Total Environ. 2018, 627, 600–612. [Google Scholar] [CrossRef] [PubMed]
- Kirchman, D. Measuring bacterial biomass production and growth rates from leucine incorporation in natural aquatic environments. Methods Microbiol. 2001, 30, 227–237. [Google Scholar] [CrossRef]
- Sibert, R.; Harrison, S.; Joye, S.B. Protocols for radiotracer estimation of primary hydrocarbon oxidation in oxygenated seawater. In Hydrocarbon and Lipid Microbiology Protocols; McGenity, T.J., Timmis, K.N., Nogales, B., Eds.; Springer: Berlin/Heidelberg, Germany, 2017; pp. 263–276. [Google Scholar]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581. [Google Scholar] [CrossRef] [Green Version]
- Callahan, B.J.; McMurdie, P.J.; Holmes, S.P. Exact sequence variants should replace operational taxonomic units in marker-gene data analysis. ISME J. 2017, 11, 2639. [Google Scholar] [CrossRef] [Green Version]
- Ziervogel, K.; D’souza, N.; Sweet, J.; Yan, B.; Passow, U. Natural oil slicks fuel surface water microbial activities in the northern Gulf of Mexico. Front. Microbiol. 2014, 5, 188. [Google Scholar] [CrossRef]
- Kleindienst, S.; Seidel, M.; Ziervogel, K.; Grim, S.; Loftis, K.; Harrison, S.; Malkin, S.Y.; Perkins, M.J.; Field, J.; Sogin, M.L. Chemical dispersants can suppress the activity of natural oil-degrading microorganisms. Proc. Natl. Acad. Sci. USA 2015, 112, 14900–14905. [Google Scholar] [CrossRef] [Green Version]
- Abuhamed, T.; Bayraktar, E.; Mehmetoğlu, T.; Mehmetoğlu, Ü. Kinetics model for growth of Pseudomonas putida F1 during benzene, toluene and phenol biodegradation. Process Biochem. 2004, 39, 983–988. [Google Scholar] [CrossRef]
- Joye, S.B.; Kleindienst, S.; Gilbert, J.A.; Handley, K.M.; Weisenhorn, P.; Overholt, W.A.; Kostka, J.E. Responses of microbial communities to hydrocarbon exposures. Oceanography 2016, 29, 136–149. [Google Scholar] [CrossRef] [Green Version]
- Pardee, A.B.; Prestidge, L.S. The initial kinetics of enzyme induction. Biochim. Biophys. Acta 1961, 49, 77–88. [Google Scholar] [CrossRef] [PubMed]
- Coulon, F.; McKew, B.A.; Osborn, A.M.; McGenity, T.J.; Timmis, K.N. Effects of temperature and biostimulation on oil-degrading microbial communities in temperate estuarine waters. Environ. Microbiol. 2007, 9, 177–186. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Nigro, L.M.; Gutierrez, T.; Joye, S.B.; Highsmith, R.; Teske, A. Pulsed blooms and persistent oil-degrading bacterial populations in the water column during and after the Deepwater Horizon blowout. Deep Sea Res. Part II Top. Stud. Oceanogr. 2016, 129, 282–291. [Google Scholar] [CrossRef] [Green Version]
- Bouchez-Naïtali, M.; Vandecasteele, J.-P. Biosurfactants, an help in the biodegradation of hexadecane? The case of Rhodococcus and Pseudomonas strains. World J. Microb. Biot. 2008, 24, 1901–1907. [Google Scholar] [CrossRef]
- Lee, D.W.; Lee, H.; Kwon, B.-O.; Khim, J.S.; Yim, U.H.; Kim, B.S.; Kim, J.-J. Biosurfactant-assisted bioremediation of crude oil by indigenous bacteria isolated from Taean beach sediment. Environ. Pollut. 2018, 241, 254–264. [Google Scholar] [CrossRef]
- Dobson, R.; Schroth, M.H.; Zeyer, J. Effect of water-table fluctuation on dissolution and biodegradation of a multi-component, light nonaqueous-phase liquid. J. Contam. Hydrol. 2007, 94, 235–248. [Google Scholar] [CrossRef]
- Kanaly, R.A.; Watanabe, K. Multiple mechanisms contribute to the biodegradation of benzo[a]pyrene by petroleum-derived multicomponent nonaqueous-phase liquids. Environ. Toxicol. Chem. 2004, 23, 850–856. [Google Scholar] [CrossRef]
- Kanaly, R.A.; Harayama, S.; Watanabe, K. Rhodanobacter sp. strain BPC1 in a benzo[a]pyrene-mineralizing bacterial consortium. Appl. Microbiol. Biotechnol. 2002, 68, 5826–5833. [Google Scholar] [CrossRef] [Green Version]
- Ron, E.Z.; Rosenberg, E. Enhanced bioremediation of oil spills in the sea. Curr. Opin. Biotech. 2014, 27, 191–194. [Google Scholar] [CrossRef] [PubMed]
- Hua, F.; Wang, H.Q. Uptake and trans-membrane transport of petroleum hydrocarbons by microorganisms. Biotechnol. Biotechnol. Equip. 2014, 28, 165–175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ulrici, W. Contaminated Soil Areas, Different Countries and Contaminants. In Biotechnology Set; Rehm, H.J., Reed, G., Eds.; Wiley-VCH: Weinheim, Germany, 2001; pp. 5–41. [Google Scholar]
- Lide, D.R. CRC Handbook of Chemistry and Physics; Haynes, W.M., Ed.; CRC Press: Boca Raton, FL, USA, 2004; Volume 85. [Google Scholar]
- Lee, R.F. Fate of petroleum components in estuarine waters of the southeastern United States. In International Oil Spill Conference; American Petroleum Institute: Washington, DC, USA, 1977; pp. 611–616. [Google Scholar]
- Lofthus, S.; Bakke, I.; Tremblay, J.; Greer, C.W.; Brakstad, O.G. Biodegradation of weathered crude oil in seawater with frazil ice. Mar. Pollut. Bull. 2020, 154, 111090. [Google Scholar] [CrossRef]
- Li, X.; Zhao, Q.; Wang, X.; Li, Y.; Zhou, Q. Surfactants selectively reallocated the bacterial distribution in soil bioelectrochemical remediation of petroleum hydrocarbons. J. Hazard. Mater. 2018, 344, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Nikolova, C.N.; Ijaz, U.Z.; Magill, C.; Kleindienst, S.; Joye, S.B.; Gutierrez, T. Response and oil degradation activities of a northeast Atlantic bacterial community to biogenic and synthetic surfactants. Microbiome 2021, 9, 191. [Google Scholar] [CrossRef] [PubMed]
- Jung, J.; Park, W. Acinetobacter species as model microorganisms in environmental microbiology: Current state and perspectives. Appl. Microbiol. Biotechnol. 2015, 99, 2533–2548. [Google Scholar] [CrossRef] [PubMed]
- Muthukumar, B.; Al Salhi, M.S.; Narenkumar, J.; Devanesan, S.; Rao, T.N.; Kim, W.; Rajasekar, A. Characterization of two novel strains of Pseudomonas aeruginosa on biodegradation of crude oil and its enzyme activities. Environ. Pollut. 2022, 304, 119223. [Google Scholar] [CrossRef]
- Liu, H.; Yang, G.; Jia, H.; Sun, B. Crude oil degradation by a novel strain Pseudomonas aeruginosa AQNU-1 isolated from an oil-contaminated lake wetland. Processes 2022, 10, 307. [Google Scholar] [CrossRef]
- Chauhan, A.; Pathak, A.; Ewida, A.Y.; Griffiths, Z.; Stothard, P. Whole genome sequence analysis of an Alachlor and Endosulfan degrading Pseudomonas strain W15Feb9B isolated from Ochlockonee River, Florida. Genom. Data 2016, 8, 134–138. [Google Scholar] [CrossRef] [Green Version]
- Masuda, H.; Shiwa, Y.; Yoshikawa, H.; Zylstra, G.J. Draft genome sequence of the versatile alkane-degrading bacterium Aquabacterium sp. strain NJ1. Genome Announc. 2014, 2, e01271-14. [Google Scholar] [CrossRef] [Green Version]
- Assil, Z.; Esegbue, O.; Mašek, O.; Gutierrez, T.; Free, A. Specific enrichment of hydrocarbonclastic bacteria from diesel-amended soil on biochar particles. Sci. Total Environ. 2021, 762, 143084. [Google Scholar] [CrossRef] [PubMed]
- Chaudhary, D.K.; Dahal, R.H.; Kim, D.-U.; Kim, J. Novosphingobium olei sp. nov., with the ability to degrade diesel oil, isolated from oil-contaminated soil and proposal to reclassify Novosphingobium stygium as a later heterotypic synonym of Novosphingobium aromaticivorans. Int. J. Syst. Evol. Microbiol. 2021, 71, 004628. [Google Scholar] [CrossRef] [PubMed]
- Segura, A.; Hernández-Sánchez, V.; Marqués, S.; Molina, L. Insights in the regulation of the degradation of PAHs in Novosphingobium sp. HR1a and utilization of this regulatory system as a tool for the detection of PAHs. Sci. Total Environ. 2017, 590, 381–393. [Google Scholar] [CrossRef]
- Brown, L.M.; Gunasekera, T.S.; Bowen, L.L.; Ruiz, O.N. Draft genome sequence of Rhodovulum sp. strain NI22, a naphthalene-degrading marine bacterium. Genome Announc. 2015, 3, e01475-14. [Google Scholar] [CrossRef]
- Orsi, W.D.; Smith, J.M.; Liu, S.; Liu, Z.; Sakamoto, C.M.; Wilken, S.; Poirier, C.; Richards, T.A.; Keeling, P.J.; Worden, A.Z. Diverse, uncultivated bacteria and archaea underlying the cycling of dissolved protein in the ocean. ISME J. 2016, 10, 2158–2173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, Y.; Feng, L.; Yang, G.; Yang, Q.; Zhang, X.; Mu, J. Intensification and microbial pathways of simultaneous nitrification–denitrification in a sequencing batch biofilm reactor for seawater-based saline wastewater treatment. J. Chem. Technol. Biotechnol. 2018, 93, 2766–2773. [Google Scholar] [CrossRef]
- Volodina, E.; Raberg, M.; Steinbüchel, A. Engineering the heterotrophic carbon sources utilization range of Ralstonia eutropha H16 for applications in biotechnology. Crit. Rev. Biotechnol. 2016, 36, 978–991. [Google Scholar] [CrossRef] [PubMed]
- McGenity, T.J.; Folwell, B.D.; McKew, B.A.; Sanni, G.O. Marine crude-oil biodegradation: A central role for interspecies interactions. Aquat. Sci. 2012, 8, 10. [Google Scholar] [CrossRef] [Green Version]
- Alharbi, N.K.; Alzaban, M.I.; Albarakaty, F.M.; Abd El-Aziz, A.R.; AlRokban, A.H.; Mahmoud, M.A. Transcriptome profiling reveals differential gene expression of laccase genes in Aspergillus terreus KC462061 during biodegradation of crude oil. Biology 2022, 11, 564. [Google Scholar] [CrossRef]
- Zhang, J.H.; Xue, Q.H.; Gao, H.; Ma, X.; Wang, P. Degradation of crude oil by fungal enzyme preparations from Aspergillus spp. for potential use in enhanced oil recovery. J. Chem. Technol. Biotechnol. 2016, 91, 865–875. [Google Scholar] [CrossRef]
- Steliga, T. Role of fungi in biodegradation of petroleum hydrocarbons in drill waste. Pol. J. Environ. Stud. 2012, 21, 471–479. [Google Scholar]
- Su, D.; Pu, Y.; Gong, C.; He, Z. Application of cold-adaptive Pseudomonas sp. SDR4 and Mortierella alpina JDR7 co-immobilized on maize cob in remediating PAH-contaminated freeze-thawed soil. Environ. Adv. 2021, 4, 100063. [Google Scholar] [CrossRef]
- Adetitun, D.; Sulaiman, R.; Oguntoye, M. Utilization of Decane, Heptane, Petrol and hexadecane by fungi isolated from engine oil contaminated soil in Ilorin. J. Environ. Manag. 2019, 23, 1671–1676. [Google Scholar] [CrossRef]
- Mitchell, C.; Millar, C.; Minter, D. Studies on decomposition of Scots pine needles. Trans. Brit. Mycol. Soc. 1978, 71, 343–348. [Google Scholar] [CrossRef]
- Doiphode, N.; Joshi, C.; Ghormade, V.; Deshpande, M. Biotechnological applications of dimorphic yeasts. In Yeast Biotechnology: Diversity and Applications; Satyanarayana, T., Kunze, G., Eds.; Springer: Berlin/Heidelberg, Germany, 2009; pp. 635–650. [Google Scholar]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, R.; Peng, C.; Ye, Y.; Tang, Y.; Lu, L. Succession Patterns of Microbial Composition and Activity following the Diesel Spill in an Urban River. Microorganisms 2023, 11, 698. https://doi.org/10.3390/microorganisms11030698
Yang R, Peng C, Ye Y, Tang Y, Lu L. Succession Patterns of Microbial Composition and Activity following the Diesel Spill in an Urban River. Microorganisms. 2023; 11(3):698. https://doi.org/10.3390/microorganisms11030698
Chicago/Turabian StyleYang, Ruiyu, Chao Peng, Yuqiu Ye, Yun Tang, and Lu Lu. 2023. "Succession Patterns of Microbial Composition and Activity following the Diesel Spill in an Urban River" Microorganisms 11, no. 3: 698. https://doi.org/10.3390/microorganisms11030698