
Genomic Diversity, Antimicrobial Susceptibility, and Biofilm Formation of Clinical Acinetobacter baumannii Isolates from Horses
 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Isolates
2.2. Whole-Genome Sequencing
2.3. Assignment of MLST Analysis and International Clones
2.4. Antimicrobial Susceptibility Testing
2.5. Resistance Genes and Biofilm-Associated Genes
2.6. Biofilm Assay
| SBF ≤ SBFT; |
| SBFT < SBF ≤ 2× SBFT; |
| 2× SBFT < SBF ≤ 4× SBFT; |
| 4× SBFT < SBF. |
2.7. Statistical Analysis
3. Results
3.1. Phylogenetic Analysis
3.2. Antimicrobial Susceptibility and AMR Genes
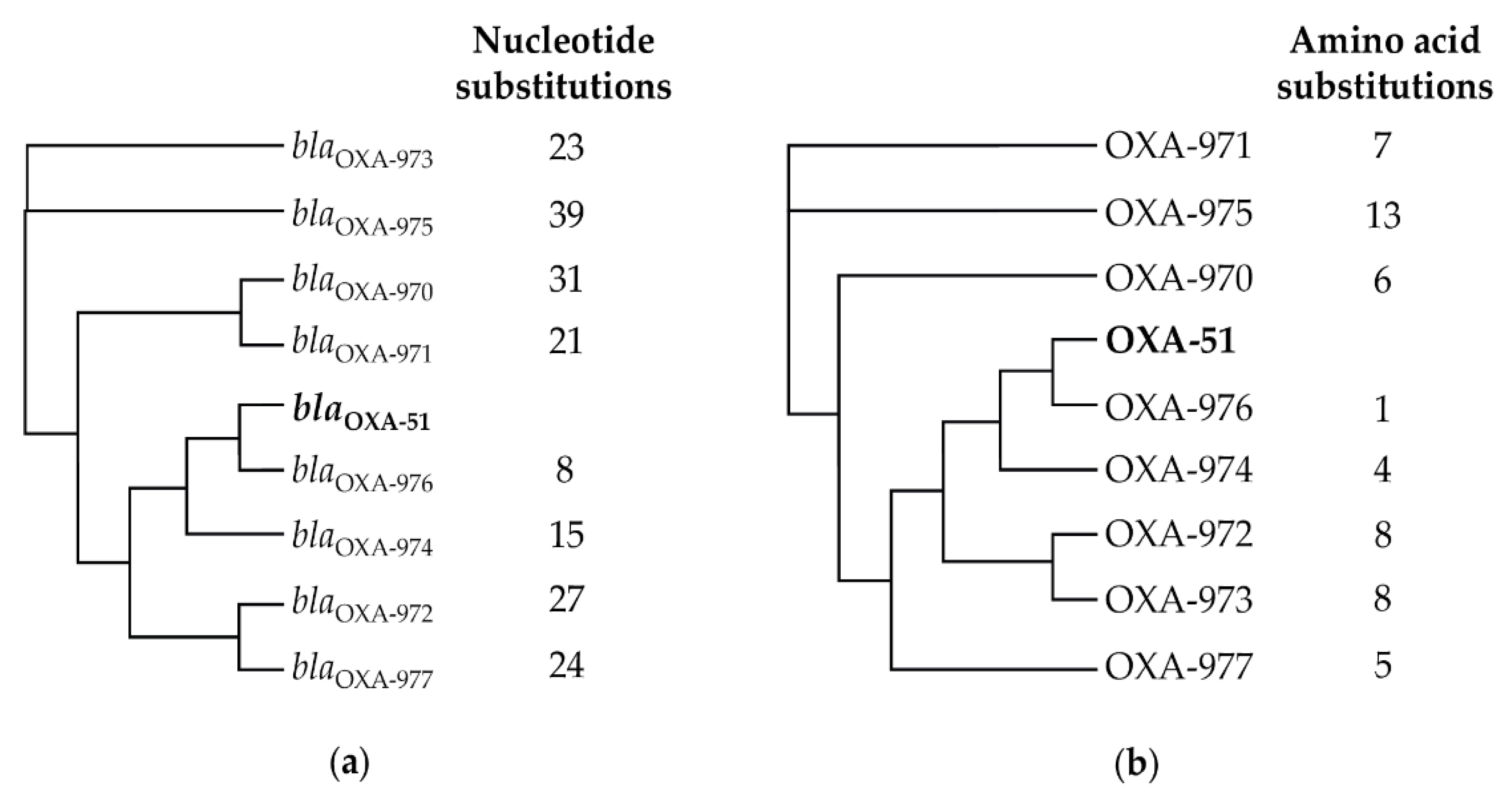
3.3. Instrinsic Oxacillinase Variants
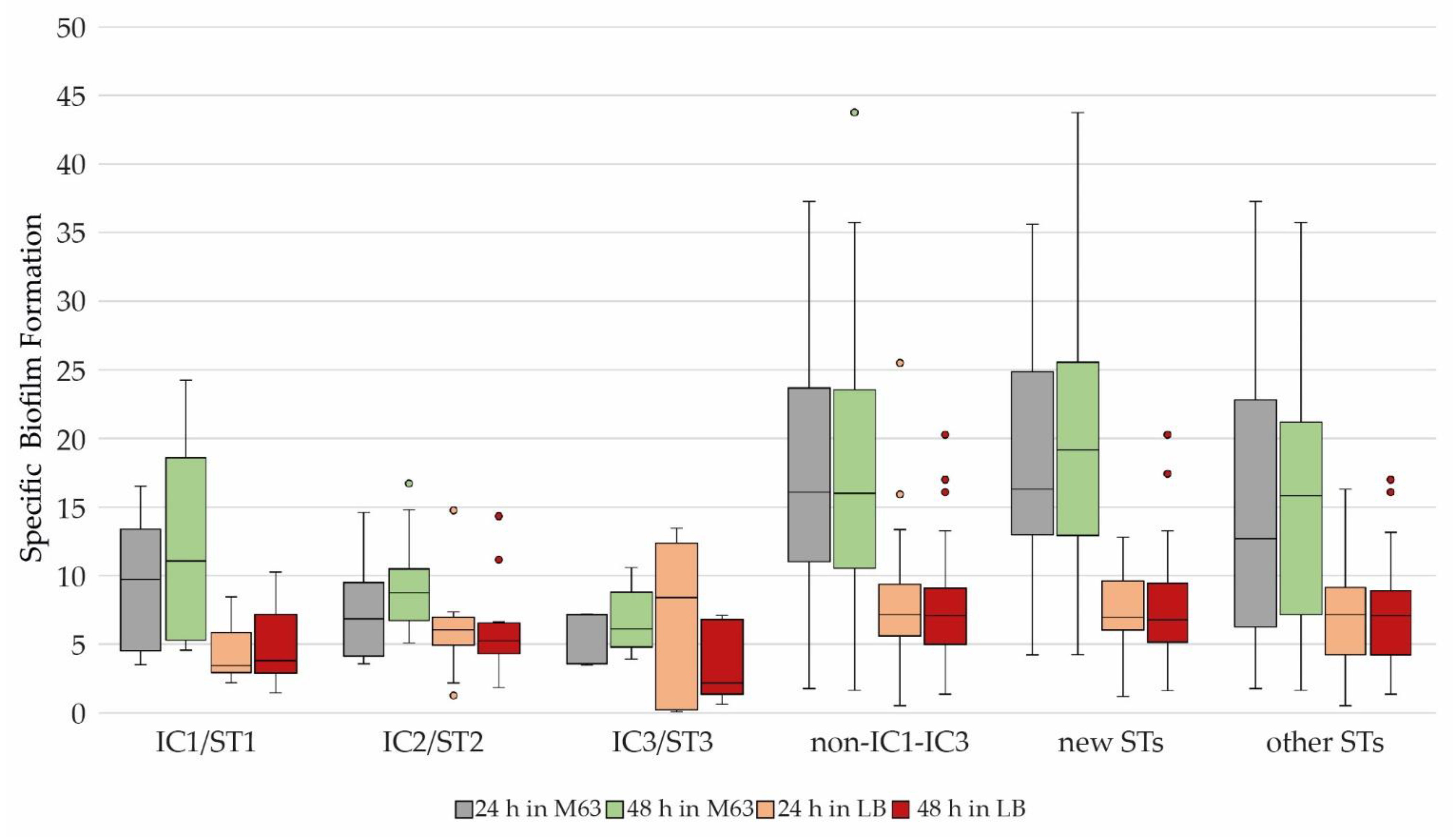
3.4. Biofilm Formation and Biofilm-Associated Genes (BAGs)
3.5. Correlation of Biofilm Formation with ICs/STs, Antimicrobial Susceptibility, and Resistance Genes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Batt, C.A.; Tortorello, M.L. (Eds.) Encyclopedia of Food Microbiology, 2nd ed.; Elsevier: Amsterdam, The Netherlands, 2014; ISBN 9780123847300. [Google Scholar]
- Almasaudi, S.B. Acinetobacter spp. as nosocomial pathogens: Epidemiology and resistance features. Saudi J. Biol. Sci. 2018, 25, 586–596. [Google Scholar] [CrossRef] [Green Version]
- Antunes, L.C.S.; Visca, P.; Towner, K.J. Acinetobacter baumannii: Evolution of a global pathogen. Pathog. Dis. 2014, 71, 292–301. [Google Scholar] [CrossRef] [Green Version]
- Wareth, G.; Neubauer, H.; Sprague, L.D. Acinetobacter baumannii—A neglected pathogen in veterinary and environmental health in Germany. Vet. Res. Commun. 2019, 43, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Rice, L.B. Federal funding for the study of antimicrobial resistance in nosocomial pathogens: No ESKAPE. J. Infect. Dis. 2008, 197, 1079–1081. [Google Scholar] [CrossRef]
- Nocera, F.P.; Attili, A.-R.; de Martino, L. Acinetobacter baumannii: Its Clinical Significance in Human and Veterinary Medicine. Pathogens 2021, 10, 127. [Google Scholar] [CrossRef]
- Higgins, P.G.; Dammhayn, C.; Hackel, M.; Seifert, H. Global spread of carbapenem-resistant Acinetobacter baumannii. J. Antimicrob. Chemother. 2010, 65, 233–238. [Google Scholar] [CrossRef] [Green Version]
- Endimiani, A.; Hujer, K.M.; Hujer, A.M.; Bertschy, I.; Rossano, A.; Koch, C.; Gerber, V.; Francey, T.; Bonomo, R.A.; Perreten, V. Acinetobacter baumannii isolates from pets and horses in Switzerland: Molecular characterization and clinical data. J. Antimicrob. Chemother. 2011, 66, 2248–2254. [Google Scholar] [CrossRef]
- Zordan, S.; Prenger-Berninghoff, E.; Weiss, R.; van der Reijden, T.; van den Broek, P.; Baljer, G.; Dijkshoorn, L. Multidrug-resistant Acinetobacter baumannii in veterinary clinics, Germany. Emerg. Infect. Dis. 2011, 17, 1751–1754. [Google Scholar] [CrossRef]
- Pour, N.K.; Dusane, D.H.; Dhakephalkar, P.K.; Zamin, F.R.; Zinjarde, S.S.; Chopade, B.A. Biofilm formation by Acinetobacter baumannii strains isolated from urinary tract infection and urinary catheters. FEMS Immunol. Med. Microbiol. 2011, 62, 328–338. [Google Scholar] [CrossRef] [Green Version]
- Longo, F.; Vuotto, C.; Donelli, G. Biofilm formation in Acinetobacter baumannii. New Microbiol. 2014, 37, 119–127. [Google Scholar]
- Tomaras, A.P.; Dorsey, C.W.; Edelmann, R.E.; Actis, L.A. Attachment to and biofilm formation on abiotic surfaces by Acinetobacter baumannii: Involvement of a novel chaperone-usher pili assembly system. Microbiology (Reading) 2003, 149, 3473–3484. [Google Scholar] [CrossRef] [Green Version]
- Eze, E.C.; Chenia, H.Y.; El Zowalaty, M.E. Acinetobacter baumannii biofilms: Effects of physicochemical factors, virulence, antibiotic resistance determinants, gene regulation, and future antimicrobial treatments. Infect. Drug Resist. 2018, 11, 2277–2299. [Google Scholar] [CrossRef] [Green Version]
- Espinal, P.; Martí, S.; Vila, J. Effect of biofilm formation on the survival of Acinetobacter baumannii on dry surfaces. J. Hosp. Infect. 2012, 80, 56–60. [Google Scholar] [CrossRef]
- Lin, F.; Xu, Y.; Chang, Y.; Liu, C.; Jia, X.; Ling, B. Molecular Characterization of Reduced Susceptibility to Biocides in Clinical Isolates of Acinetobacter baumannii. Front. Microbiol. 2017, 8, 1836. [Google Scholar] [CrossRef]
- Sarshar, M.; Behzadi, P.; Scribano, D.; Palamara, A.T.; Ambrosi, C. Acinetobacter baumannii: An Ancient Commensal with Weapons of a Pathogen. Pathogens 2021, 10, 387. [Google Scholar] [CrossRef]
- Choi, A.H.K.; Slamti, L.; Avci, F.Y.; Pier, G.B.; Maira-Litrán, T. The pgaABCD locus of Acinetobacter baumannii encodes the production of poly-beta-1-6-N-acetylglucosamine, which is critical for biofilm formation. J. Bacteriol. 2009, 191, 5953–5963. [Google Scholar] [CrossRef] [Green Version]
- Pakharukova, N.; Tuittila, M.; Paavilainen, S.; Malmi, H.; Parilova, O.; Teneberg, S.; Knight, S.D.; Zavialov, A.V. Structural basis for Acinetobacter baumannii biofilm formation. Proc. Natl. Acad. Sci. USA 2018, 115, 5558–5563. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.-J.; Kim, N.-Y.; Ko, S.-Y.; Park, S.-Y.; Oh, M.-H.; Shin, M.-S.; Lee, Y.-C.; Lee, J.-C. Complementary Regulation of BfmRS Two-Component and AbaIR Quorum Sensing Systems to Express Virulence-Associated Genes in Acinetobacter baumannii. Int. J. Mol. Sci. 2022, 23, 13136. [Google Scholar] [CrossRef]
- Roy, S.; Chowdhury, G.; Mukhopadhyay, A.K.; Dutta, S.; Basu, S. Convergence of Biofilm Formation and Antibiotic Resistance in Acinetobacter baumannii Infection. Front. Med. 2022, 9, 793615. [Google Scholar] [CrossRef]
- Larsen, M.V.; Cosentino, S.; Rasmussen, S.; Friis, C.; Hasman, H.; Marvig, R.L.; Jelsbak, L.; Sicheritz-Pontén, T.; Ussery, D.W.; Aarestrup, F.M.; et al. Multilocus sequence typing of total-genome-sequenced bacteria. J. Clin. Microbiol. 2012, 50, 1355–1361. [Google Scholar] [CrossRef] [Green Version]
- Center for Genomic Epidemiology. MLST. Available online: https://cge.food.dtu.dk/services/MLST/ (accessed on 8 September 2022).
- Turton, J.F.; Gabriel, S.N.; Valderrey, C.; Kaufmann, M.E.; Pitt, T.L. Use of sequence-based typing and multiplex PCR to identify clonal lineages of outbreak strains of Acinetobacter baumannii. Clin. Microbiol. Infect. 2007, 13, 807–815. [Google Scholar] [CrossRef] [Green Version]
- Feng, Y.; Zou, S.; Chen, H.; Yu, Y.; Ruan, Z. BacWGSTdb 2.0: A one-stop repository for bacterial whole-genome sequence typing and source tracking. Nucleic Acids Res. 2021, 49, 644–650. [Google Scholar] [CrossRef]
- Clinical and Laboratory Standards Institute. Performance Standards for Antimicrobial Susceptibility Testing, 32nd ed.; CLSI supplement M100; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2022; ISBN 978-1-68440-134-5. [Google Scholar]
- European Committee on Antimicrobial Susceptibility Testing. Breakpoint Tables for Interpretation of MICs and Zone Diameters, Version 12.0. 2022. Available online: http://www.eucast.org (accessed on 30 December 2022).
- Bortolaia, V.; Kaas, R.S.; Ruppe, E.; Roberts, M.C.; Schwarz, S.; Cattoir, V.; Philippon, A.; Allesoe, R.L.; Rebelo, A.R.; Florensa, A.F.; et al. ResFinder 4.0 for predictions of phenotypes from genotypes. J. Antimicrob. Chemother. 2020, 75, 3491–3500. [Google Scholar] [CrossRef]
- Zankari, E.; Allesøe, R.; Joensen, K.G.; Cavaco, L.M.; Lund, O.; Aarestrup, F.M. PointFinder: A novel web tool for WGS-based detection of antimicrobial resistance associated with chromosomal point mutations in bacterial pathogens. J. Antimicrob. Chemother. 2017, 72, 2764–2768. [Google Scholar] [CrossRef] [Green Version]
- Center for Genomic Epidemiology. Services. Available online: http://www.genomicepidemiology.org/services/ (accessed on 8 September 2022).
- National Library of Medicine (US); National Center for Biotechnology Information. BLAST: Basic Local Alignment Search Tool. Available online: https://blast.ncbi.nlm.nih.gov/Blast.cgi (accessed on 14 February 2022).
- BacWGSTdb. Multiple Genome Analysis. Available online: http://bacdb.cn/BacWGSTdb/analysis_multiple.php (accessed on 8 September 2022).
- O’Toole, G.A. Microtiter Dish Biofilm Formation Assay. J. Vis. Exp. 2011, 47, 2437–2438. [Google Scholar] [CrossRef]
- Qi, L.; Li, H.; Zhang, C.; Liang, B.; Li, J.; Wang, L.; Du, X.; Liu, X.; Qiu, S.; Song, H. Relationship between Antibiotic Resistance, Biofilm Formation, and Biofilm-Specific Resistance in Acinetobacter baumannii. Front. Microbiol. 2016, 7, 483. [Google Scholar] [CrossRef] [Green Version]
- Malaka de Silva, P.; Chong, P.; Fernando, D.M.; Westmacott, G.; Kumar, A. Effect of Incubation Temperature on Antibiotic Resistance and Virulence Factors of Acinetobacter baumannii ATCC 17978. Antimicrob. Agents Chemother. 2018, 62, e01514–e01517. [Google Scholar] [CrossRef] [Green Version]
- Stepanović, S.; Vuković, D.; Dakić, I.; Savić, B.; Švabić-Vlahović, M. A modified microtiter-plate test for quantification of staphylococcal biofilm formation. J. Microbiol. Methods 2000, 40, 175–179. [Google Scholar] [CrossRef]
- Magiorakos, A.-P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Harbarth, S.; Hindler, J.F.; Kahlmeter, G.; Olsson-Liljequist, B.; et al. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: An international expert proposal for interim standard definitions for acquired resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef] [Green Version]
- Evans, B.A.; Amyes, S.G.B. OXA β-lactamases. Clin. Microbiol. Rev. 2014, 27, 241–263. [Google Scholar] [CrossRef] [Green Version]
- Vaneechoutte, M.; Devriese, L.A.; Dijkshoorn, L.; Lamote, B.; Deprez, P.; Verschraegen, G.; Haesebrouck, F. Acinetobacter baumannii-infected vascular catheters collected from horses in an equine clinic. J. Clin. Microbiol. 2000, 38, 4280–4281. [Google Scholar] [CrossRef] [Green Version]
- van der Kolk, J.H.; Endimiani, A.; Graubner, C.; Gerber, V.; Perreten, V. Acinetobacter in veterinary medicine, with an emphasis on Acinetobacter baumannii. J. Glob. Antimicrob. Resist. 2019, 16, 59–71. [Google Scholar] [CrossRef]
- Walther, B.; Klein, K.-S.; Barton, A.-K.; Semmler, T.; Huber, C.; Wolf, S.A.; Tedin, K.; Merle, R.; Mitrach, F.; Guenther, S.; et al. Extended-spectrum beta-lactamase (ESBL)-producing Escherichia coli and Acinetobacter baumannii among horses entering a veterinary teaching hospital: The contemporary “Trojan Horse”. PLoS ONE 2018, 13, e0191873. [Google Scholar] [CrossRef] [Green Version]
- Müller, S.; Janssen, T.; Wieler, L.H. Multidrug resistant Acinetobacter baumannii in veterinary medicine--emergence of an underestimated pathogen? Berl. Munch. Tierarztl. Wochenschr. 2014, 127, 435–446. [Google Scholar]
- Jacobmeyer, L.; Semmler, T.; Stamm, I.; Ewers, C. Genomic Analysis of Acinetobacter baumannii Isolates Carrying OXA-23 and OXA-58 Genes from Animals Reveals ST1 and ST25 as Major Clonal Lineages. Antibiotics 2022, 11, 1045. [Google Scholar] [CrossRef]
- Lupo, A.; Châtre, P.; Ponsin, C.; Saras, E.; Boulouis, H.-J.; Keck, N.; Haenni, M.; Madec, J.-Y. Clonal Spread of Acinetobacter baumannii Sequence Type 25 Carrying blaOXA-23 in Companion Animals in France. Antimicrob. Agents Chemother. 2017, 61, e01881-16. [Google Scholar] [CrossRef] [Green Version]
- Hérivaux, A.; Pailhoriès, H.; Quinqueneau, C.; Lemarié, C.; Joly-Guillou, M.-L.; Ruvoen, N.; Eveillard, M.; Kempf, M. First report of carbapenemase-producing Acinetobacter baumannii carriage in pets from the community in France. Int. J. Antimicrob. Agents 2016, 48, 220–221. [Google Scholar] [CrossRef]
- Göttig, S.; Gruber, T.M.; Higgins, P.G.; Wachsmuth, M.; Seifert, H.; Kempf, V.A.J. Detection of pan drug-resistant Acinetobacter baumannii in Germany. J. Antimicrob. Chemother. 2014, 69, 2578–2579. [Google Scholar] [CrossRef]
- van Spijk, J.N.; Schmitt, S.; Fürst, A.; Schoster, A. A retrospective analysis of antimicrobial resistance in bacterial pathogens in an equine hospital (2012–2015). Schweiz Arch Tierheilkd 2016, 158, 433–442. [Google Scholar] [CrossRef] [Green Version]
- Karalewitz, A.P.-A.; Miller, S.I. Multidrug-Resistant Acinetobacter baumannii Chloramphenicol Resistance Requires an Inner Membrane Permease. Antimicrob. Agents Chemother. 2018, 62, e00513–e00518. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.-H.; Kuo, S.-C.; Lee, Y.-T.; Chang, I.C.Y.; Yang, S.-P.; Chen, T.-L.; Fung, C.-P. Amino acid substitutions of quinolone resistance determining regions in GyrA and ParC associated with quinolone resistance in Acinetobacter baumannii and Acinetobacter genomic species 13TU. J. Microbiol. Immunol. Infect 2012, 45, 108–112. [Google Scholar] [CrossRef] [Green Version]
- Turton, J.F.; Ward, M.E.; Woodford, N.; Kaufmann, M.E.; Pike, R.; Livermore, D.M.; Pitt, T.L. The role of ISAba1 in expression of OXA carbapenemase genes in Acinetobacter baumannii. FEMS Microbiol. Lett. 2006, 258, 72–77. [Google Scholar] [CrossRef] [Green Version]
- Ewers, C.; Klotz, P.; Leidner, U.; Stamm, I.; Prenger-Berninghoff, E.; Göttig, S.; Semmler, T.; Scheufen, S. OXA-23 and ISAba1-OXA-66 class D β-lactamases in Acinetobacter baumannii isolates from companion animals. Int. J. Antimicrob. Agents 2017, 49, 37–44. [Google Scholar] [CrossRef]
- Jacobmeyer, L.; Stamm, I.; Semmler, T.; Ewers, C. First report of NDM-1 in an Acinetobacter baumannii strain from a pet animal in Europe. J. Glob. Antimicrob. Resist. 2021, 26, 128–129. [Google Scholar] [CrossRef]
- Rafei, R.; Hamze, M.; Pailhoriès, H.; Eveillard, M.; Marsollier, L.; Joly-Guillou, M.-L.; Dabboussi, F.; Kempf, M. Extrahuman epidemiology of Acinetobacter baumannii in Lebanon. Appl. Environ. Microbiol. 2015, 81, 2359–2367. [Google Scholar] [CrossRef] [Green Version]
- Smet, A.; Boyen, F.; Pasmans, F.; Butaye, P.; Martens, A.; Nemec, A.; Deschaght, P.; Vaneechoutte, M.; Haesebrouck, F. OXA-23-producing Acinetobacter species from horses: A public health hazard? J. Antimicrob. Chemother. 2012, 67, 3009–3010. [Google Scholar] [CrossRef] [Green Version]
- Rahimi, S.; Farshadzadeh, Z.; Taheri, B.; Mohammadi, M.; Haghighi, M.-A.; Bahador, A. The Relationship Between Antibiotic Resistance Phenotypes and Biofilm Formation Capacity in Clinical Isolates of Acinetobacter baumannii. Jundishapur J. Microbiol. 2018, 11, e74315. [Google Scholar] [CrossRef] [Green Version]
- Yang, C.-H.; Su, P.-W.; Moi, S.-H.; Chuang, L.-Y. Biofilm Formation in Acinetobacter Baumannii: Genotype-Phenotype Correlation. Molecules 2019, 24, 1849. [Google Scholar] [CrossRef] [Green Version]
- de Gregorio, E.; Del Franco, M.; Martinucci, M.; Roscetto, E.; Zarrilli, R.; Di Nocera, P.P. Biofilm-associated proteins: News from Acinetobacter. BMC Genom. 2015, 16, 933. [Google Scholar] [CrossRef] [Green Version]
- Gaddy, J.A.; Tomaras, A.P.; Actis, L.A. The Acinetobacter baumannii 19606 OmpA protein plays a role in biofilm formation on abiotic surfaces and in the interaction of this pathogen with eukaryotic cells. Infect. Immun. 2009, 77, 3150–3160. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Clinical Origin | IC1 | IC2 | IC3 | Non-IC1-IC3 |
|---|---|---|---|---|
| Abscess | 0 | 1 | 0 | 1 |
| Eye | 1 | 1 | 1 | 0 |
| Feces | 1 | 2 | 0 | 0 |
| Gastrointestinal tract | 0 | 1 | 0 | 1 |
| Genital tract | 0 | 1 | 1 | 18 |
| Organs | 0 | 1 | 0 | 3 |
| Respiratory tract | 1 | 1 | 1 | 11 |
| Skin, hair, hoof | 0 | 0 | 0 | 6 |
| Urinary tract | 0 | 0 | 1 | 0 |
| Wound | 5 | 6 | 1 | 5 |
| Others | 0 | 2 | 0 | 4 |
| Total number of isolates | 8 | 16 | 5 | 49 |
| Phenotypic Resistance | Genotypic Resistance | |||
|---|---|---|---|---|
| Antibiotic Class | Antibiotic | R (%) | AMR Genes | Positive Isolates (%) |
| β-lactams | * Ampicillin * Amoxicillin–Clavulanate * Cefalexin Ceftiofur Cefpodoxime Imipenem | 100 100 100 98.7 94.9 0 | blaADC-25 blaOXA-51-like blaTEM-1D | 98.7 100 30.8 |
| Aminoglycosides | Gentamicin Amikacin | 38.5 1.3 | aadA1 aa(3)-Ia aph(3’)-VIa aph(3′)-Ia aph(3‘)-Ic aph(6)-Id | 29.5 32.1 5.1 23.1 30.8 23.1 |
| Phenicols | * Chloramphenicol | 100 | catA1 ABUW_0982 | 26.9 100 |
| Sulfonamides | Trimethoprim–sulfamethoxazole | 37.2 | sul1 sul2 | 35.9 2.6 |
| Tetracyclines | Tetracycline | 33.3 | tet(A) tet(B) tet(39) | 10.3 1.3 1.3 |
| Fluoroquinolones | Enrofloxacin Marbofloxacin | 37.2 35.9 | none # | ─ |
| none # | ─ | |||
| Nitrofuran derivates | Nitrofurantoin | 100 | none | ─ |
| Culture Medium | Incubation Time (h) | SBF | ||
|---|---|---|---|---|
| Min | Max | Median | ||
| LB | 24 | 0.09 | 25.49 | 6.61 |
| 48 | 0.64 | 20.27 | 6.01 | |
| M63 | 24 | 1.75 | 37.28 | 12.15 |
| 48 | 1.64 | 43.76 | 12.92 | |
| BAGs | Position in Reference Genome | Product | Function | Positive Isolates (%) | ||||
|---|---|---|---|---|---|---|---|---|
| All (n = 78) | IC1 (n = 8) | IC2 (n = 16) | IC3 (n = 5) | Non-IC1-IC3 (n = 49) | ||||
| abaI | 1396054-1396605 | N-acyl-L-homoserine lactone synthetase | Quorum-sensing system: regulates biofilm formation and surface motility | 93.59 | 100 | 100 | 100 | 93.59 |
| abaR | 1394083-1394799 | DNA-binding HTH-domain-containing protein | 98.72 | 100 | 100 | 100 | 98.72 | |
| bfmR | 2304914-2305630 | Biofilm-controlling response regulator | Quorum sensing regulated two-component system involved in biofilm formation | 100 | 100 | 100 | 100 | 100 |
| bfmS | 2305663-2307312 | Signal transduction histidine kinase | 98.72 | 100 | 100 | 80.00 | 100 | |
| bap | 536313-541547 | Biofilm-associated protein (Bap) | Biofilm formation | 55.13 | 0 | 100 | 0 | 24.36 |
| blp1 | 869025-879134 | Bap-like protein 1 | 0 | 0 | 0 | 0 | 0 | |
| blp2 | 1099370-1101556 | Bap-like protein 2 | 75.64 | 100 | 100 | 100 | 78.72 | |
| csuA | 3998748-3999194 | Csu pilus subunit A | Biofilm formation | 93.59 | 100 | 100 | 100 | 93.59 |
| csuA/B | 3999371-3999907 | Csu pilus major pilin subunit CsuA/B | 97.44 | 100 | 100 | 100 | 97.44 | |
| csuB | 3998224-3998742 | Csu pilus subunit B | 92.31 | 100 | 100 | 60.00 | 92.31 | |
| csuC | 3997397-3998230 | Csu pilus chaperone protein | 98.72 | 100 | 100 | 100 | 98.72 | |
| csuD | 3994902-3997400 | Csu pilus usher protein | 98.72 | 100 | 100 | 100 | 98.72 | |
| csuE | 3993886-3994905 | Csu pilus tip adhesin | 98.72 | 100 | 100 | 100 | 98.72 | |
| ompA | 703280-704350 | Outer membrane protein A | Antibiotic and serum resistance, biofilm formation, host interaction, cytotoxicity, interference with autophagy and apoptosis | 55.13 | 0 | 100 | 0 | 57.69 |
| pgaA | 3941315-3942688 | poly-beta-1,6 N-acetyl-D-glucosamine export porin | Biofilm formation | 100 | 100 | 100 | 100 | 100 |
| pgaB | 3938457-3940286 | poly-beta-1,6-N-acetyl-D-glucosamine N-deacetylase | 98.72 | 100 | 100 | 100 | 98.72 | |
| pgaC | 3937210-3938457 | poly-beta-1,6 N-acetyl-D-glucosamine synthase | 97.44 | 100 | 100 | 100 | 97.44 | |
| pgaD | 3936749-3937213 | poly-beta-1,6-N-acetyl-D-glucosamine biosynthesis protein | 97.44 | 100 | 100 | 100 | 93.59 | |
| Medium | Classification | No. of Isolates | Acquired Phenotypic Resistance to CTU, GEN, ENR, TET, and SXT (0 to 5 Antibiotics) * | |||||
|---|---|---|---|---|---|---|---|---|
| 0 | 1 | 2 | 3 | 4 | 5 | |||
| LB 24 h | non-biofilm | 29 | 0% | 44.8% | 0% | 3.4% | 10.3% | 41.4% |
| weak | 43 | 0% | 72.1% | 0% | 0% | 4.7% | 23.3% | |
| moderate | 4 | 0% | 25.0% | 0% | 0% | 75.0% | 0% | |
| strong | 2 | 50.0% | 50.0% | 0% | 0% | 0% | 0% | |
| LB 48 h | non-biofilm | 47 | 0% | 46.8% | 0% | 2.1% | 8.5% | 42.6% |
| weak | 26 | 0% | 80.8% | 0% | 0% | 11.5% | 7.7% | |
| moderate | 5 | 20.0% | 60.0% | 0% | 0% | 20.0% | 0% | |
| strong | 0 | - | - | - | - | - | ||
| M63 24 h | non-biofilm | 41 | 2.4% | 34.1% | 0% | 2.4% | 4.9% | 46.3% |
| weak | 27 | 0% | 81.5% | 0% | 0% | 7.4% | 11.1% | |
| moderate | 10 | 0% | 100% | 0% | 0% | 0% | 0% | |
| strong | 0 | - | - | - | - | - | ||
| M63 48 h | non-biofilm | 44 | 2.3% | 38.6% | 0% | 2.3% | 13.6% | 43.2% |
| weak | 30 | 0% | 83.3% | 0% | 0% | 6.7% | 10.0% | |
| moderate | 4 | 0% | 100% | 0% | 0% | 0% | 0% | |
| strong | 0 | - | - | - | - | - | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rühl-Teichner, J.; Jacobmeyer, L.; Leidner, U.; Semmler, T.; Ewers, C. Genomic Diversity, Antimicrobial Susceptibility, and Biofilm Formation of Clinical Acinetobacter baumannii Isolates from Horses. Microorganisms 2023, 11, 556. https://doi.org/10.3390/microorganisms11030556
Rühl-Teichner J, Jacobmeyer L, Leidner U, Semmler T, Ewers C. Genomic Diversity, Antimicrobial Susceptibility, and Biofilm Formation of Clinical Acinetobacter baumannii Isolates from Horses. Microorganisms. 2023; 11(3):556. https://doi.org/10.3390/microorganisms11030556
Chicago/Turabian StyleRühl-Teichner, Johanna, Lisa Jacobmeyer, Ursula Leidner, Torsten Semmler, and Christa Ewers. 2023. "Genomic Diversity, Antimicrobial Susceptibility, and Biofilm Formation of Clinical Acinetobacter baumannii Isolates from Horses" Microorganisms 11, no. 3: 556. https://doi.org/10.3390/microorganisms11030556