
Risk of Exposure to Coccidioides spp. in the Temblor Special Recreation Management Area (SRMA), Kern County, CA
Abstract
:1. Introduction
2. Material and Methods
2.1. Sampling Plan
2.2. Sampling and Sample Storage
2.3. Safety
2.4. Soil Parameters
2.5. DNA Extraction and PCR
2.6. Statistics
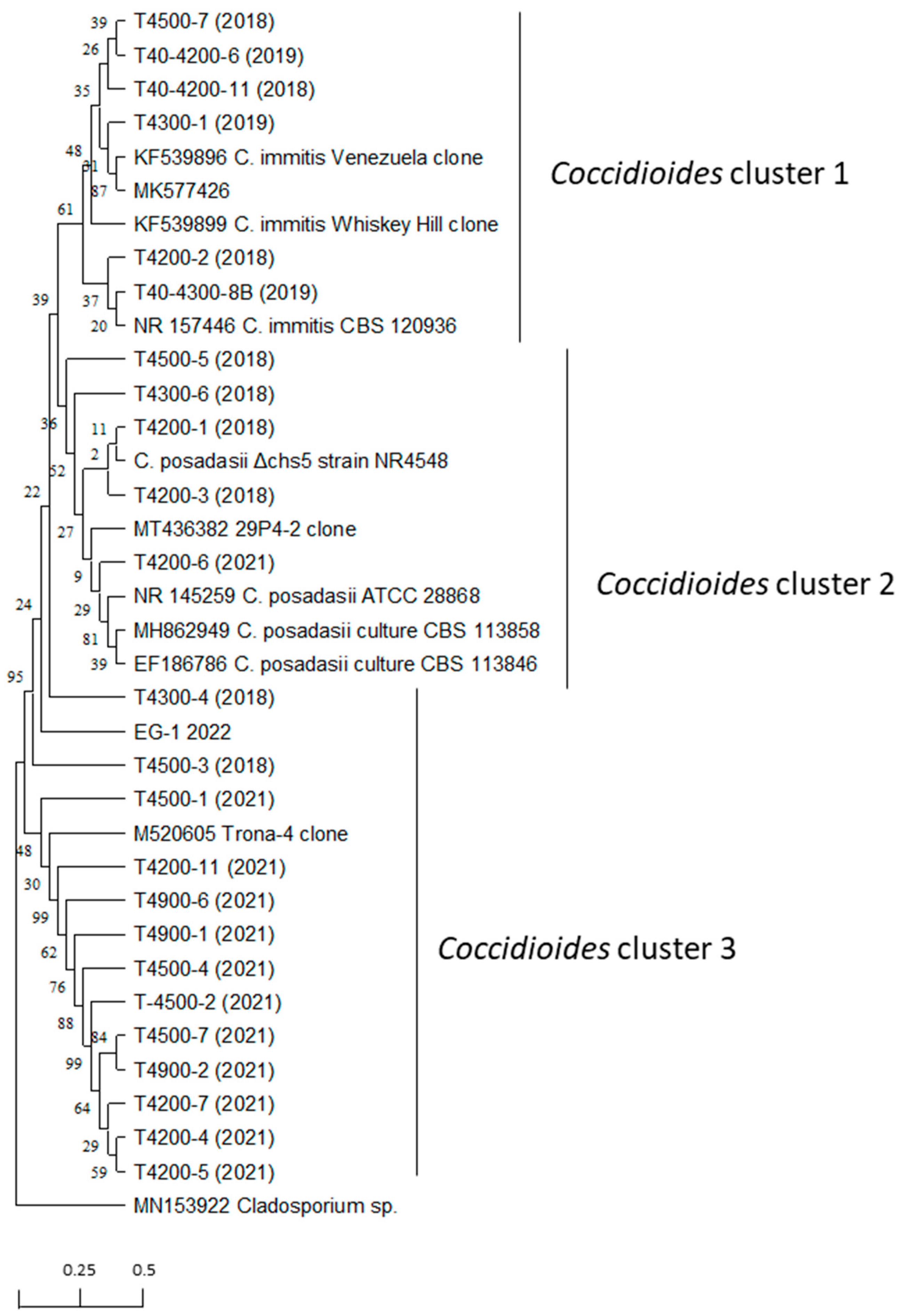
2.7. Evolutionary Relationships of Taxa
3. Results
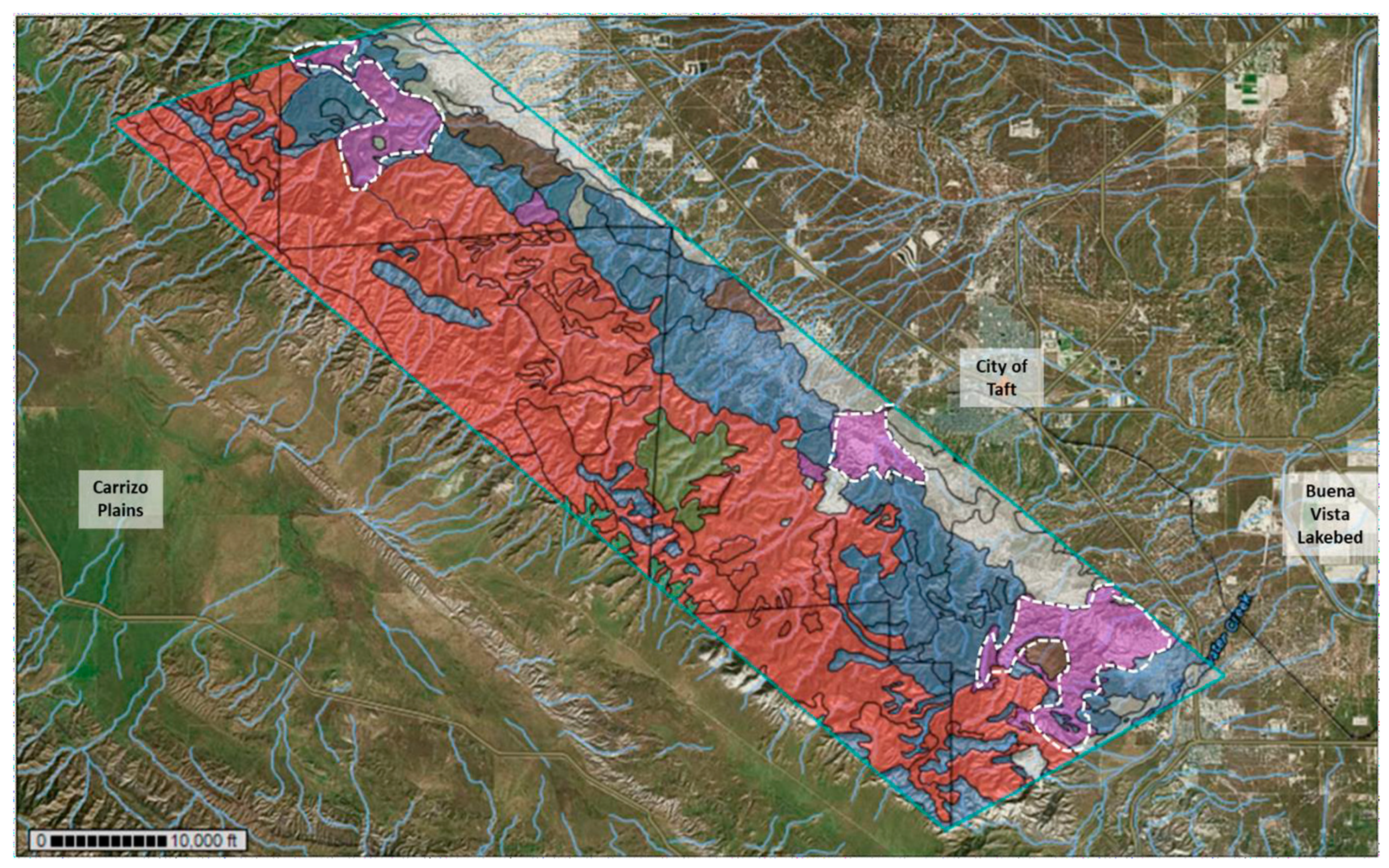
3.1. The Temblor SRMA: Soil Types and Landscape
3.2. Sampling
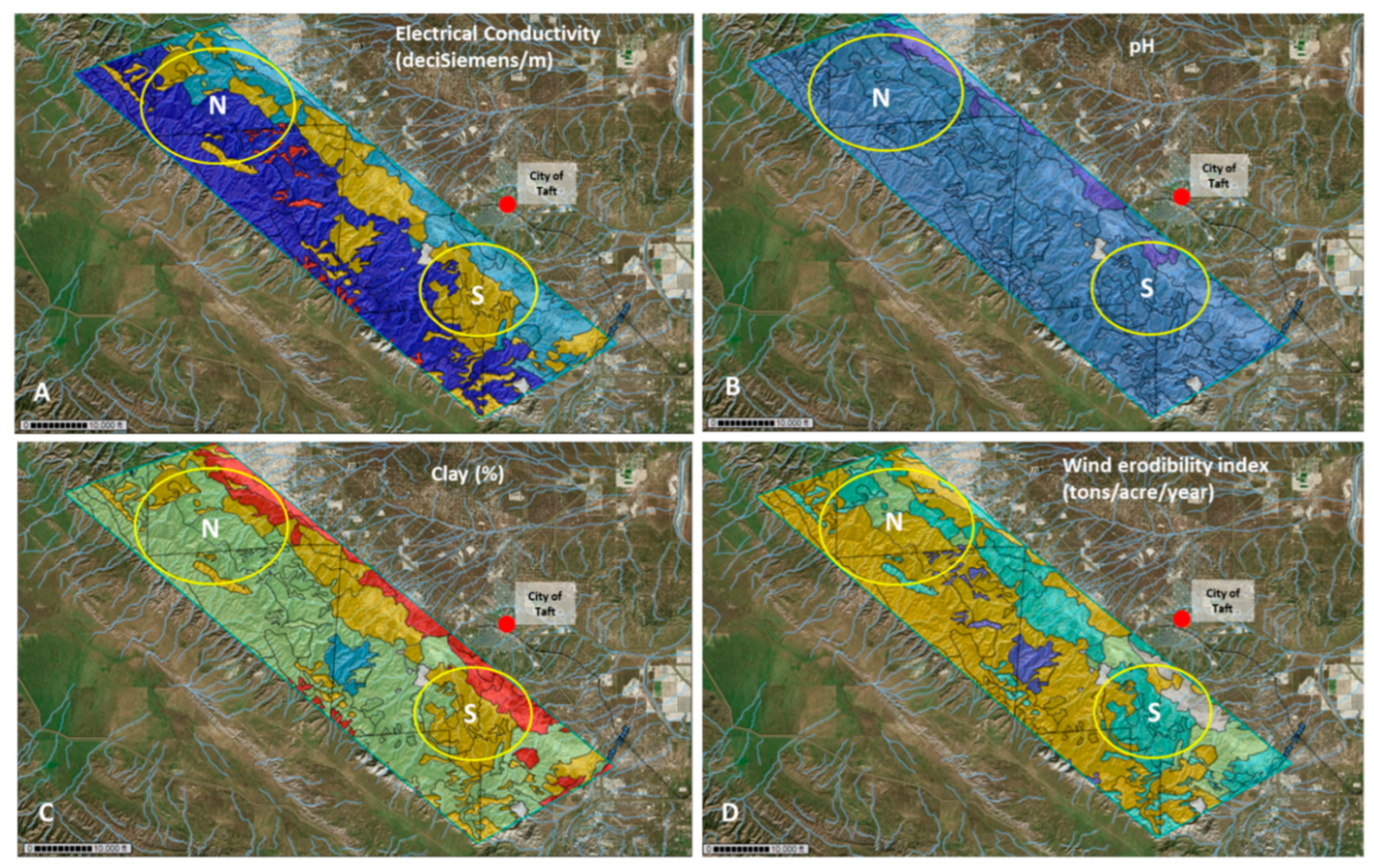
3.3. Environmental Parameters
3.4. Detection of Coccidioides
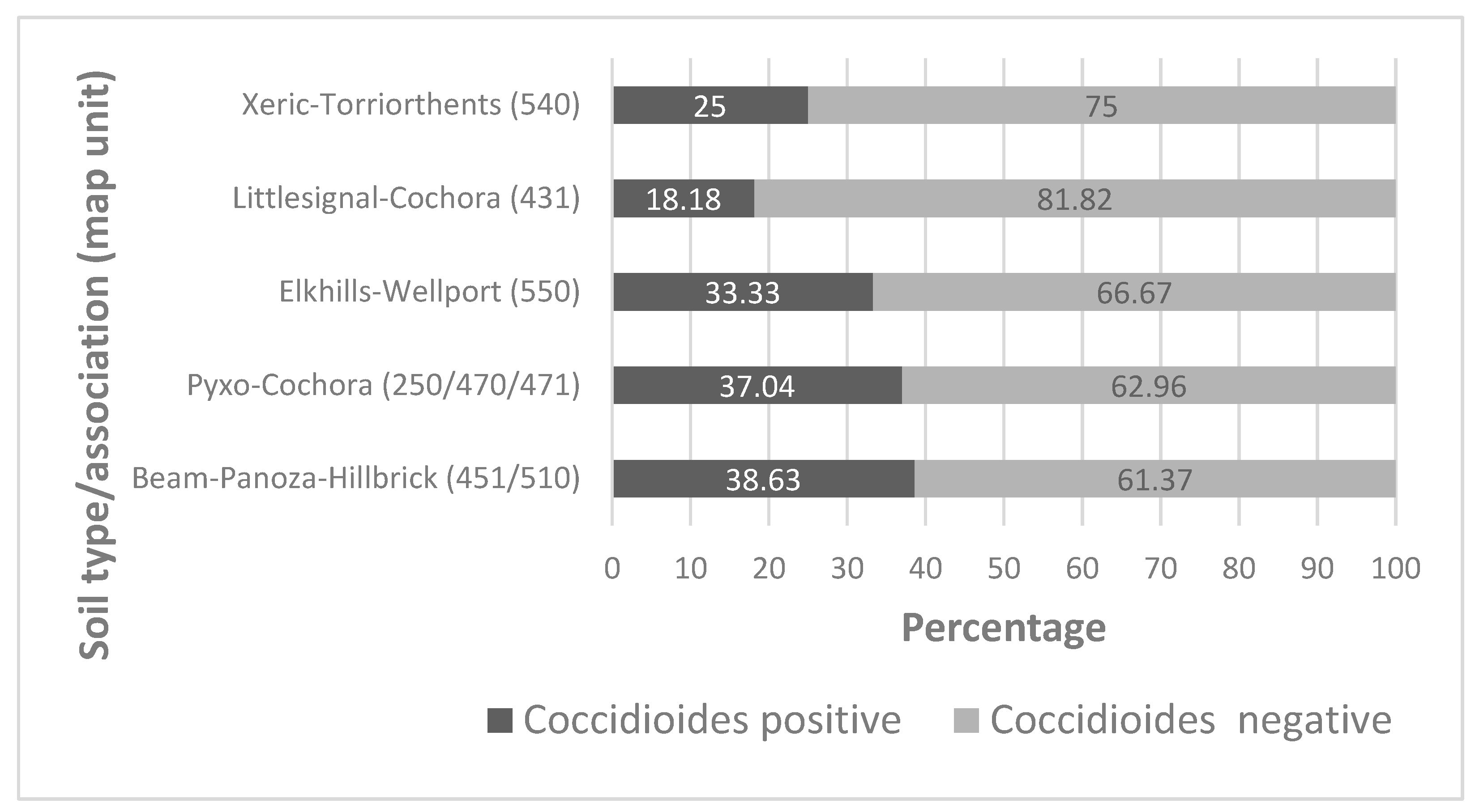
3.5. Environmental Parameters and Coccidioides
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- National Cooperative Soil Survey. Temblor Series. 1999. Available online: https://soilseries.sc.egov.usda.gov/OSD_Docs/T/TEMBLOR.html (accessed on 10 November 2022).
- Fisher, M.C.; Koenig, G.; White, T.; San-Blas, G.; Negroni, R.; Alvarez, I.G.; Wanke, B.; Taylor, J.W. Biogeographic range expansion into South America by Coccidioides immitis mirrors New World patterns of human migration. Proc. Natl. Acad. Sci. USA 2001, 98, 4558–4562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fisher, M.C.; Koenig, G.L.; White, T.J.; Taylor, J.W. Molecular and phenotypic description of Coccidioides posadasii sp. nov., previously recognized as the non-California population of Coccidioides immitis. Mycologia 2002, 94, 73–84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cooksey Sondermeyer, G.; Nguyen, A.; Knutson, K.; Tabnak, F.; Benedict, K.; McCotter, O.; Jain, S.; Vugia, D. Notes from the field: Increase in coccidioidomycosis—California, 2016. Morb. Mortal. Wkly. Rep. 2017, 66, 833. [Google Scholar] [CrossRef] [Green Version]
- Williams, S.L.; Chiller, T. Update on the epidemiology, diagnosis, and treatment of coccidioidomycosis. J. Fungi 2022, 8, 666. [Google Scholar] [CrossRef] [PubMed]
- Galgiani, J.N.; Ampel, N.M.; Blair, J.E.; Catanzaro, A.; Geertsma, F.; Hoover, S.E.; Johnson, R.H.; Kusne, S.; Lisse, J.; MacDonald, J.D.; et al. 2016 Infectious Diseases Society of America (IDSA) Clinical Practice Guideline for the Treatment of Coccidioidomycosis. Clin. Infect. Dis. 2016, 63, e112–e146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hector, R.F.; Rutherford, G.W.; Tsang, C.A.; Erhart, L.M.; McCotter, O.; Anderson, S.M.; Komatsu, K.; Tabnak, F.; Vugia, D.J.; Yang, Y.; et al. The public health impact of coccidioidomycosis in Arizona and California. Int. J. Environ. Res. Public Health 2011, 8, 1150–1173. [Google Scholar] [CrossRef]
- Wilson, L.; Ting, J.; Lin, H.; Shah, R.; MacLean, M.; Peterson, M.W.; Stockamp, N.; Lipke, R.; Brown, P. The rise of valley fever: Prevalence and cost burden of coccidioidomycosis infection in California. Int. J. Environ. Res. Public Health 2019, 16, 1113. [Google Scholar] [CrossRef] [Green Version]
- Benedict, K.; Jackson, B.R.; Chiller, T.; Beer, K.D. Estimation of direct healthcare costs of fungal diseases in the United States. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2019, 68, 1791–1797. [Google Scholar] [CrossRef] [Green Version]
- Benedict, K.; Whitham, H.K.; Jackson, B.R. Economic Burden of Fungal Diseases in the United States. Open Forum Infect. Dis. 2022, 9, ofac097. [Google Scholar] [CrossRef]
- Lauer, A.; Baal, J.D.; Baal, J.C.; Verma, M.; Chen, J.M. Detection of Coccidioides immitis in Kern County, California, by multiplex PCR. Mycologia 2012, 104, 62–69. [Google Scholar] [CrossRef]
- Dobos, R.R. Mapping Soi Habitat for Coccidioides. Natl. Coop. Soil Surv. (NCSS) Newsl. 2014, 66, 9–12. [Google Scholar]
- Sigler, L.; Flis, A.L.; Carmicheal, J.W. The genus Uncinocarpus (Onygenaceae) and its synonym Brunneospora: New concepts, combinations, and connections to anamorphs in Chrysosporium, and further evidence of relationship with Coccidioides immitis. Can. J. Bot. 1998, 76, 1624–1636. [Google Scholar]
- Kollath, D.R.; Teixeira, M.M.; Funke, A.; Miller, K.J.; Barker, B.M. Investigating the role of animal burrows on the ecology and distribution of Coccidioides spp. in Arizona soils. Mycopathologia 2020, 185, 145–159. [Google Scholar] [CrossRef]
- Wollum, A.G. Soil sampling for microbiological analysis. In Methods of Soil Analysis: Part 2 Microbiological and Biochemical Properties, 1st ed.; Weaver, R.W., Bottomley, P., Bezdicek, D., Smith, S., Tabatabai, A., Wollom, A.G., Eds.; Soil Science Society of America, Inc.: Madison WI, USA, 1994; Volume 1, pp. 1–4. [Google Scholar] [CrossRef]
- USEPA/USGS Sample Collection Protocol for Bacterial Pathogens in Surface Soil|Science Inventory; U.S. Environmental Protection Agency (EPA), Washington, DC, USA. 2014. Available online: https://cfpub.epa.gov/si/si_public_record_report.cfm?Lab=NHSRC&dirEntryId=285571 (accessed on 11 November 2022).
- Soil Survey Staff, Natural Resources Conservation Service, United States Department of Agriculture. Web Soil Survey. Available online: https://websoilsurvey.sc.egov.usda.gov/App/HomePage.htm (accessed on 2 November 2022).
- Baptista-Rosas, R.C.; Catalan-Dibene, J.; Romero-Olivares, A.L.; Hinojosa, A.; Cavazos, T.; Riquelme, M. Molecular detection of Coccidioides spp. from environmental samples in Baja California: Linking Valley Fever to soil and climate conditions. Fungal Ecol. 2012, 5, 177–190. [Google Scholar] [CrossRef]
- Johnson, S.M.; Carlson, E.L.; Fisher, F.S.; Pappagianis, D. Demonstration of Coccidioides immitis and Coccidioides posadasii DNA in soil samples collected from Dinosaur National Monument, Utah. Sabouraudia 2014, 52, 610–617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benson, D.A.; Clark, K.; Karsch-Mizrachi, I.; Lipman, D.J.; Ostell, J.; Sayers, E.W. GenBank. Nucleic Acids Res. 2010, 42, D32–D37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013; Available online: http://www.R-project.org/ (accessed on 9 November 2022).
- Tamura, K.; Stecher, G.; Kumar, S. MEGA 11: Molecular Evolutionary Genetics Analysis Version 11; Molecular Biology and Evolution: Oxford, UK, 2021. [Google Scholar] [CrossRef]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [CrossRef]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Tamura, K.; Nei, M.; Kumar, S. Prospects for inferring very large phylogenies by using the neighbor-joining method. Proc. Natl. Acad. Sci. USA 2004, 101, 11030–11035. [Google Scholar] [CrossRef] [Green Version]
- Fisher, F.S.; Bultman, M.W.; Johnson, S.M.; Pappagianis, D.; Zaborsky, E. Coccidioides niches and habitat parameters in the southwestern United States. Ann. N. Y. Acad. Sci. 2007, 1111, 47–72. [Google Scholar] [CrossRef] [Green Version]
- Baptista-Rosas, R.C.; Hinojosa, A.; Riquelme, M. Ecological niche modeling of Coccidioides spp. in western North American deserts. Ann. N. Y. Acad. Sci. 2007, 1111, 35–46. [Google Scholar] [CrossRef] [PubMed]
- Lacy, G.H.; Swatek, F.E. Soil ecology of Coccidioides immitis at Amerindian middens in California. Appl. Microbiol. 1974, 27, 379–388. [Google Scholar] [CrossRef] [PubMed]
- Talamantes, J.; Behseta, S.; Zender, C.S. Statistical modeling of valley fever data in Kern County, California. Int. J. Biometeorol. 2007, 51, 307–313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kollath, D.R.; Mihaljevic, J.R.; Barker, B.M. PM10 and Other Climatic Variables Are Important Predictors of Seasonal Variability of Coccidioidomycosis in Arizona. Microbiol. Spectr. 2022, 10, e01483-21. [Google Scholar] [CrossRef]
- Vargas-Gastélum, L.; Romero-Olivares, A.L.; Escalante, A.E.; Rocha-Olivares, A.; Brizuela, C.; Riquelme, M. Impact of seasonal changes on fungal diversity of a semi-arid ecosystem revealed by 454 pyrosequencing. FEMS Microbiol. Ecol. 2015, 91, fiv044. [Google Scholar] [CrossRef] [Green Version]
- Chow, N.A.; Kangiser, D.; Gade, L.; McCotter, O.Z.; Hurst, S.; Salamone, A.; Wohrle, R.; Clifford, W.; Kim, S.; Salah, Z.; et al. Factors influencing distribution of Coccidioides immitis in soil, Washington State, 2016. Msphere 2021, 6, e00598-21. [Google Scholar] [CrossRef]
- Gade, L.; McCotter, O.Z.; Bowers, J.R.; Waddell, V.; Brady, S.; Carvajal, J.A.; Sunenshine, R.; Komatsu, K.K.; Engelthaler, D.M.; Chiller, T.; et al. The detection of Coccidioides from ambient air in Phoenix, Arizona: Evidence of uneven distribution and seasonality. Med. Mycol. 2020, 58, 552–559. [Google Scholar] [CrossRef] [Green Version]
- Dobos, R.R.; Benedict, K.; Jackson, B.R.; McCotter, O.Z. Using soil survey data to model potential Coccidioides soil habitat and inform Valley fever epidemiology. PLoS ONE 2021, 16, e0247263. [Google Scholar] [CrossRef]
- Pearson, D.; Ebisu, K.; Wu, X.; Basu, R. A Review of Coccidioidomycosis in California: Exploring the Intersection of Land-use, Population Movement, and Climate Change. Epidemiol. Rev. 2019, 41, 145–157. [Google Scholar] [CrossRef]
- Lauer, A.; Etyemezian, V.; Nikolich, G.; Kloock, C.; Arzate, A.F.; Sadiq Batcha, F.; Kaur, M.; Garcia, E.; Mander, J.; Kayes Passaglia, A. Valley fever: Environmental risk factors and exposure pathways deduced from field measurements in California. Int. J. Environ. Res. Public Health 2020, 17, 5285. [Google Scholar] [CrossRef]
- Goossens, D.; Buck, B.; McLaurin, B. Contributions to atmospheric dust production of natural and anthropogenic emissions in a recreational area designated for off-road vehicular activity (Nellis Dunes, Nevada, USA). J. Arid Environ. 2012, 78, 80–99. [Google Scholar] [CrossRef]
- Tong, D.Q.; Gorris, M.E.; Gill, T.E.; Ardon-Dryer, K.; Wang, J.; Ren, L. Dust storms, valley fever, and public awareness. GeoHealth 2022, 6, e2022GH000642. [Google Scholar] [CrossRef] [PubMed]
- Diab, S.; Johnson, S.M.; Garcia, J.; Carlson, E.L.; Pappagianis, D.; Smith, J.; Uzal, F.A. Case report: Abortion and disseminated infection by Coccidioides posadasii in an alpaca (Vicugna pacos) fetus in Southern California. Med. Mycol. Case Rep. 2013, 2, 159–162. [Google Scholar] [CrossRef] [PubMed]
- Hamm, P.S.; Hutchison, M.I.; Leonard, P.; Melman, S.; Natvig, D.O. First analysis of human Coccidioides isolates from New Mexico and the southwest Four Corners region: Implications for the distributions of C. posadasii and C. immitis and human groups at risk. J. Fungi 2019, 5, 74. [Google Scholar] [CrossRef] [Green Version]
- Chaturvedi, S.; Victor, T.R.; Marathe, A.; Sidamonidze, K.; Crucillo, K.L.; Chaturvedi, V. Real-time PCR assay for detection and differentiation of Coccidioides immitis and Coccidioides posadasii from culture and clinical specimens. PLoS Negl. Trop. Dis. 2021, 15, e0009765. [Google Scholar] [CrossRef]
- Muhs, D.R. Airborne dust fall on the California Channel Islands, USA. J. Arid Environ. 1983, 6, 223–238. [Google Scholar] [CrossRef]
- Muhs, D.R.; Budahn, J.R.; Johnson, D.L.; Reheis, M.; Beann, J.; Skipp, G.; Fisher, E.; Jones, J.A. Geochemical evidence for airborne dust additions to soils in Channel Islands National Park, California. Geol. Soc. Am. Bull. 2008, 120, 106–126. [Google Scholar] [CrossRef]
- Jardine, G.E.; Crocker, A.J.; Bailey, I.; Cooper, M.J.; Milton, J.A.; Wilson, P.A. The imprint of windblown dust from the North American Southwest on the California Channel Islands and Pacific Ocean sediments. Quat. Sci. Rev. 2021, 261, 106934. [Google Scholar] [CrossRef]
- Gorris, M.E.; Cat, L.A.; Zender, C.S.; Treseder, K.K.; Randerson, J.T. Coccidioidomycosis dynamics in relation to climate in the southwestern United States. GeoHealth 2018, 2, 6–24. [Google Scholar] [CrossRef]
- Huckabone, S.E.; Gulland, F.M.; Johnson, S.M.; Colegrove, K.M.; Dodd, E.M.; Pappagianis, D.; Dunkin, R.C.; Casper, D.; Carlson, E.L.; Sykes, J.E.; et al. Coccidioidomycosis and other systemic mycoses of marine mammals stranding along the central California, USA coast: 1998–2012. J. Wildl. Dis. 2015, 51, 295–308. [Google Scholar] [CrossRef]
- Lauer, A.; Palmer, L.; Kloock, C.; Liwanag, H.E.; Norris, T.; McDonald, B.; Muñoz, J.M.; Mulcahy, C.; Hannh, S.; Johnson, S. Survey for Coccidioides Antibodies in blood Sera from California Sea Lions and Northern Fur Seals that Stranded Along the Coast of California (2013–2015). West. Wildl. 2019, 6, 69–81. [Google Scholar]
- Gorris, M.E.; Cat, L.A.; Matlock, M.; Ogunseitan, O.A.; Treseder, K.K.; Randerson, J.T.; Zender, C.S. Coccidioidomycosis (Valley fever) case data for the southwestern United States. Open Health Data 2020, 7. [Google Scholar] [CrossRef] [Green Version]
- Duarte-Escalante, E.; Reyes-Montes, M.D.R.; Frías-De-León, M.G.; Meraz-Ríos, B. Effect of Climate Change on the Incidence and Geographical Distribution of Coccidioidomycosis. In The Impact of Climate Change on Fungal Diseases, 1st ed.; Springer: Cham, Switzerland, 2022; pp. 131–143. [Google Scholar] [CrossRef]
- Valley Fever, Tailgate Training Guide for California Construction Workers; California Department of Public Health (CDPH). 2020. Available online: https://ucanr.edu/sites/safety/files/333256.pdf (accessed on 11 November 2022).
- Acute Communicable Disease Control Program and Department of Environmental Health, Coccidioidomycosis (Valley Fever) Management Plan: Guidelines for Employers; County of Los Angeles Public Health. Published August 2019. Available online: http://publichealth.lacounty.gov/acd/docs/valleyfeverplan2019.pdf (accessed on 11 November 2022).
- Be Aware of Valley Fever; Centers for Disease Control and Prevention (CDC), Mission and Community Service Groups. Published September 2019. Available online: https://www.cdc.gov/fungal/diseases/coccidioidomycosis/pdf/be-aware-of-valley-fever-h.pdf (accessed on 11 November 2022).
- Valley Fever (Coccidioidomycosis) Statistics, CEDC 24/7: Saving Lives, Protecting People™, Centers for Disease Control and Prevention (CDC). 2022. Available online: https://www.cdc.gov/fungal/diseases/coccidioidomycosis/statistics.html (accessed on 11 November 2022).
- Valley Fever is on the Rise in California; California Department of Public Health (CDPH). 2022. Available online: https://www.cdph.ca.gov (accessed on 11 November 2022).
- Dusty Work Increases Valley Fever; California Department of Public Health (CDPH), Occupational Health Branch. Published September 2022. Available online: https://www.cdph.ca.gov (accessed on 11 November 2022).
- Valley Fever, Symptoms; California Department of Public Health (CDPH). August 2022. Available online: https://www.cdph.ca.gov (accessed on 11 November 2022).









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sampling, November 2018 | |||
|---|---|---|---|
| Soil Type and Soil Map Unit # | Number of Soil Samples n (North = 18, South = 23) | Detection of Coccidioides (n) | Detection of Coccidioides (%) |
| Elkhills-Welport association (550) | 1 (north), 3 (south) | 0 (north), 1 (south) | 25.00 |
| Pyxo-Cochora association (470, 471) | 3 (north), 6 (south) | 1 (north), 4 (south) | 55.55 |
| Beam-Panoza-Hillbrick association (510, 221, 223, 227, 451) | 8 (north), 7 (south) | 3 (north), 4 (south) | 46.67 |
| Littlesignal-Cochora association (431) | 2 (north), 3 (south) | 1 (north), 0 (south) | 20.00 |
| Elkhills-Pyxo association (440) | 1 (north), 0 (south) | 0 (north), 0 (south) | 0 |
| Pyxo-Kimberlina-Cochora association (472) | 1 (north), 0 (south) | 1 (north), 0 (south) | 100.00 |
| Xeric-Torrirothents-Badlands complex (249, 540) | 1 (north), 4 (south) | 0 (north), 2 (south) | 40.00 |
| Reward-Hillbrick association (580) | 1 (north), 0 (south) | 0 (north), 0 (south) | 0 |
| Sampling, June 2019 | |||
| Soil Type and Soil Map Unit # | Number of Soil Samples n (North = 19, South = 10) | Detection of Coccidioides (n) | Detection of Coccidioides (%) |
| Elkhills-Welport association (550) | 4 (north), 0 (south) | 1 (north), 0 (south) | 25.00 |
| Elkhills-Pyxo association (440) | 1 (north), 0 (south) | 0 (north), 0 (south) | 0 |
| Pyxo-Cochora association (470, 471) | 1 (north), 2 (south) | 0 (north), 0 (south) | 0 |
| Beam-Panoza-Hillbrick association (510, 221, 223, 227, 451) | 10 (north), 4 (south) | 3 (north), 1 (south) | 28.57 |
| Littlesignal-Cochora association (431) | 0 (north), 2 (south) | 0 (north), 0 (south) | 0 |
| Pyxo-Kimberlina-Cochora association (472) | 0 (north), 0 (south) | not re-sampled | 0 |
| Xeric-Torrirothents-Badlands complex (249, 540) | 0 (north), 0 (south) | not re-sampled | 0 |
| Reward-Hillbrick-association (580) | 0 (north), 0 (south) | not re-sampled | 0 |
| Guijarral gravelly sandy loam (193) | 0 (north), 1 (south) | 0 (north), 0 (south) | 0 |
| Xerorthents-Badlands complex (219) | 3 (north), 0 (south) | 0 (north), 0 (south) | 0 |
| Padres sandy loam (490) | 0 (north), 1 (south) | 0 (north), 0 (south) | 0 |
| Sampling March 2021 | |||
| Soil Type and Soil Map Unit # | Number of Soil Samples n (North = 20, South = 24) | Detection of Coccidioides (n) | Detection of Coccidioides (%) |
| Elkhills-Welport association (550) | 3 (north), 2 (south) | 0 (north), 2 (south) | 40.00 |
| Pyxo-Cochora association (470, 471) | 5 (north), 9 (south) | 0 (north), 5 (south) | 35.71 |
| Beam-Panoza-Hillbrick association (510, 221, 223, 227, 451) | 5 (north), 10 (south) | 0 (north), 5 (south) | 40.00 |
| Littlesignal-Cochora association (431) | 3 (north), 1 (south) | 0 (north), 0 (south) | 25.00 |
| Elkhills-Pyxo association (440) | 1 (north), 0 (south) | 0 (north), 0 (south) | 0 |
| Pyxo-Kimberlina-Cochora association (472) | 1 (north), 2 (south) | 0 (north), 0 (south) | 0 |
| Xeric-Torrirothents-Badlands complex (249, 540) | 1 (north), 0 (south) | 0 (north), 0 (south) | 0 |
| Polonio clay loam (170) | 1 (north), 0 (south) | 0 (north), 0 (south) | 0 |
| Xerorthents-Badlands complex (219) | 0 (north), 0 (south) | 0 (north), 0 (south) | 0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lauer, A.; Lopez, J.J.; Chabolla, M.; Kloock, C. Risk of Exposure to Coccidioides spp. in the Temblor Special Recreation Management Area (SRMA), Kern County, CA. Microorganisms 2023, 11, 518. https://doi.org/10.3390/microorganisms11020518
Lauer A, Lopez JJ, Chabolla M, Kloock C. Risk of Exposure to Coccidioides spp. in the Temblor Special Recreation Management Area (SRMA), Kern County, CA. Microorganisms. 2023; 11(2):518. https://doi.org/10.3390/microorganisms11020518
Chicago/Turabian StyleLauer, Antje, Jocelyne Jassiri Lopez, Michael Chabolla, and Carl Kloock. 2023. "Risk of Exposure to Coccidioides spp. in the Temblor Special Recreation Management Area (SRMA), Kern County, CA" Microorganisms 11, no. 2: 518. https://doi.org/10.3390/microorganisms11020518