
The Restriction–Modification Systems of Clostridium carboxidivorans P7

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains and Cultivation Conditions
2.2. Plasmid Construction, Sequencing and Transformation
2.3. Methylation Analysis
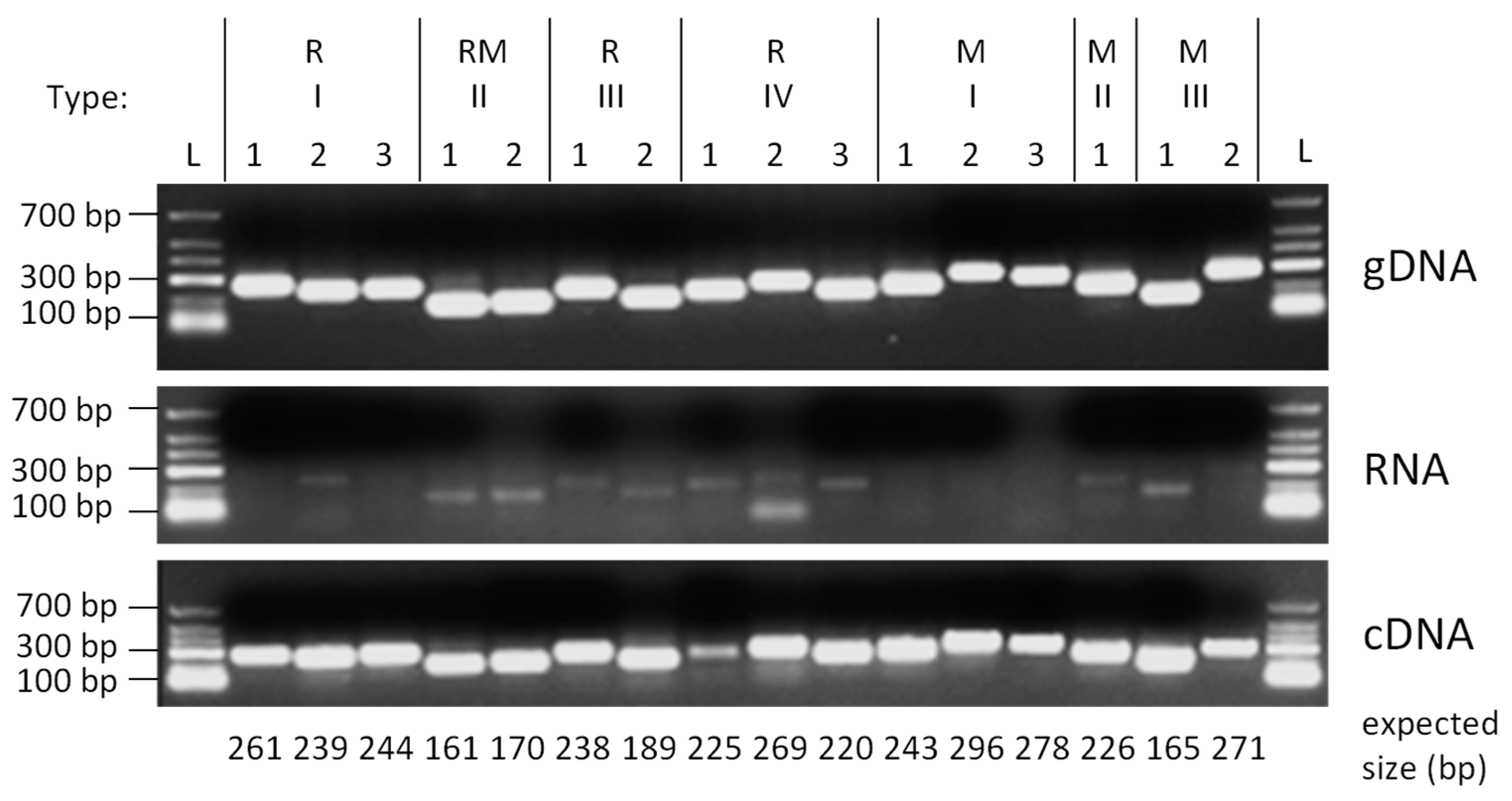
2.4. RNA Isolation and Semi Quantitative RT-PCR
3. Results and Discussion
3.1. Analysis of Methylation Activity in C. carboxidivorans
3.2. Expression of C. carboxidivorans MTases for In Vivo Methylation in E. coli
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain Name | Properties | |
|---|---|---|
| E. coli | NEB 5-alpha a | Strain for cloning of plasmids |
| E. coli | soloI.1 | Strain with MTase I.1 (S.CcaP7I + M.CcaP7I) |
| E. coli | soloI.2 | Strain with MTase I.2 (M.CcaP7II + S.CcaP7II) |
| E. coli | soloI.3 | Strain with MTase I.3 (S.CcaP7III + M.CcaP7III) |
| E. coli | soloII.1 | Strain with MTase II.1 (M.CcaP7ORF16660P) |
| E. coli | soloIIRM1 | Strain with MTase and fused restriction enzyme IIRM1 (CcaP7IV) |
| E. coli | soloIIRM2 | Strain with MTase and fused restriction enzyme IIRM2 (CcaP7ORF7120P) |
| E. coli | soloIII.1 | Strain with MTase III.1 (M.CcaP7ORF17830P) |
| E. coli | soloIII.2 | Strain with MTase III.1 (M.CcaP7ORF8705P) |
| Solo MTs | Primer Sequence (5′–3′) | Anneals |
|---|---|---|
| pMT1-3 fw (pCDF1-3 fw) | TTAACCTAGGCTGCTGCCACCGCTGAGCAATAACTAGC | pCDFDuet backbone |
| pCDF Duet rv neu | CATGGTATATCTCCTTATTAAAGTTAAACAAAATTATTTCTAC | pCDFDuet backbone |
| TypeIII MT1_fwd | TAATAAGGAGATATACCATGGCTAACTTAATTGAAAAC | C. carboxidivorans Type III M1 |
| TypeIII MT1_rev | GTGGCAGCAGCCTAGGTTAATATCCTATATACTCATAATATCTT TTATATCATTTC | C. carboxidivorans Type III M1 |
| TypeIII MT2_fwd | TGTTTAACTTTAATAAGGAGATATACCATGGAAAAAGTATATG CATTTG | C. carboxidivorans Type III M2 |
| TypeIII MT2_rev | TTGCTCAGCGGTGGCAGCAGCCTAGGTTAAATGCCTACCATG ACTTTAG | C. carboxidivorans Type III M2 |
| TypeII MT1_fwd | TGTTTAACTTTAATAAGGAGATATACCATTAGAAATAAAGTAA TAAATAAAGAGTGC | C. carboxidivorans Type II M1 |
| TypeII MT1_rev | TTGCTCAGCGGTGGCAGCAGCCTAGGTTAAGCTCTAAACAAA TTTAAGCTG | C. carboxidivorans Type II M1 |
| TypeII RM1_fwd | TGTTTAACTTTAATAAGGAGATATACCATGTATAAAGACGTTA AATTAGAAAAAAG | C. carboxidivorans Type II R+M 1 |
| TypeII RM1_rev | TTGCTCAGCGGTGGCAGCAGCCTAGGTTAATTCTTACAGCAGT TTATCTC | C. carboxidivorans Type II R+M 1 |
| TypeII RM2_fwd | TGTTTAACTTTAATAAGGAGATATACCATGGATAAGACTAAAG TAAAATCC | C. carboxidivorans Type II R+M 2 |
| TypeII RM2_rev | TTGCTCAGCGGTGGCAGCAGCCTAGGTTAAGTTAATTATATCT TAAAAAATAAATCCTTTTTAAC | C. carboxidivorans Type II R+M 2 |
| TypeI.1_fwd | TGTTTAACTTTAATAAGGAGATATACCATGTTAAACAGCGAGA CAAAAAG | C. carboxidivorans Type I M+S 1 |
| TypeI.1_rev | TTGCTCAGCGGTGGCAGCAGCCTAGGTTAATTAATTAAATAGT TCTCCTTTGAAAG | C. carboxidivorans Type I M+S 1 |
| TypeI.2_fwd | TGTTTAACTTTAATAAGGAGATATACCATGAATACACAAGAGA TAGTAAG | C. carboxidivorans Type I M+S 2 |
| TypeI.2_rev | TTGCTCAGCGGTGGCAGCAGCCTAGGTTAACTATATATCTTTAC TCAATATTTCCC | C. carboxidivorans Type I M+S 2 |
| TypeI.3_fwd | TGTTTAACTTTAATAAGGAGATATACCATGGAAAAAAACAAA AATAAACC | C. carboxidivorans Type I M+S 3 |
| TypeI.3_rev | TTGCTCAGCGGTGGCAGCAGCCTAGGTTAACTAAATTTTTACA CCTAAAATCTTCAATTG | C. carboxidivorans Type I M+S 3 |
| Primer | Primer Sequence (5′–3′) | Product Size (bp) | Anneals |
|---|---|---|---|
| RI.1_fw | AGTGAGCCTAGACAGGTTTG | 261 | C. carboxidivorans Type I R 1 |
| RI.1_rv | CCAGCTTGCCTCCATTAATC | C. carboxidivorans Type I R 1 | |
| RI.2_fw | AGCAACAGTACAGGCTATGG | 239 | C. carboxidivorans Type I R 2 |
| RI.2_rv | GGCTGGTGTAGCTGTAAGTG | C. carboxidivorans Type I R 2 | |
| RI.3_fw | AAGTGGCAGTTACGTTTAGC | 244 | C. carboxidivorans Type I R 3 |
| RI.3_rv | AGTTCTGGTGCATCAAATCC | C. carboxidivorans Type I R 3 | |
| RMII.1_fw | AGAAAGAATAAGCAGTGCAAAG | 161 | C. carboxidivorans Type II R+M 1 |
| RMII.1_rv | AGTCTATCAGGCAGTACAAATC | C. carboxidivorans Type II R+M 1 | |
| RMII.2_fw | GACATAGGAGCAAGGTATTGTC | 170 | C. carboxidivorans Type II R+M 2 |
| RMII.2_rv | TCGCCTACCTGGATATTGTAAG | C. carboxidivorans Type II R+M 2 | |
| RIII.1_fw | TCGGCCTTAAGAGAAGGTTG | 238 | C. carboxidivorans Type III R 1 |
| RIII.1_rv | TTACGCCACCATCTTCTTCG | C. carboxidivorans Type III R 1 | |
| RIII.2_fw | TGGATGGGATTGTCCGAGAG | 189 | C. carboxidivorans Type III R 2 |
| RIII.2_rv | GAAAGGCTGCCGACTTTAAC | C. carboxidivorans Type III R 2 | |
| RIV.1_fw | AAGTGCTGGATAGAGCAAATAC | 225 | C. carboxidivorans Type IV R 1 |
| RIV.1_rv | CAAACTGCATGTCACATTGTTC | C. carboxidivorans Type IV R 1 | |
| RIV.2_fw | TACACAGCTATTCGCAATGATG | 269 | C. carboxidivorans Type IV R 2 |
| RIV.2_rv | TCCAGCCACTTTATTGTTTCAC | C. carboxidivorans Type IV R 2 | |
| RIV.3_fw | ATGGACTAGAGGCGGATATG | 220 | C. carboxidivorans Type IV R 3 |
| RIV.3_rv | GCTGCTTTCTCCAAGTACTG | C. carboxidivorans Type IV R 3 | |
| MI.1_fw | AACCCATGTGCTGAGGATAAG | 243 | C. carboxidivorans Type I M1 |
| MI.1_rv | GTTCATGGAGGCTATTCTAACC | C. carboxidivorans Type I M1 | |
| MI.2_fw | TGGCTCTTATGAATGCTATGC | 296 | C. carboxidivorans Type I M2 |
| MI.2_rv | TATCTTTGTTCCGTCACCTTCC | C. carboxidivorans Type I M2 | |
| MI.3_fw | AGTAGTTACGAGCGGTGTAG | 278 | C. carboxidivorans Type I M3 |
| MI.3_rv | GTGGATGGTCAATCCCTTTC | C. carboxidivorans Type I M3 | |
| MII.1_fw | TGCCTATGAAAGCACATGAAG | 226 | C. carboxidivorans Type II M1 |
| MII.1_rv | TGTGTTTGGTGCAGAGGATAAG | C. carboxidivorans Type II M1 | |
| MIII.1_fw | GCTGCAGGGTATGAAAGTTG | 165 | C. carboxidivorans Type III M1 |
| MIII.1_rv | AAGAGGCGATGGTCGTTTAG | C. carboxidivorans Type III M1 | |
| MIII.2_fw | GCAGCAGTACCAATTCTCAATC | 271 | C. carboxidivorans Type III M2 |
| MIII.2_rv | AAAGTAACTCCTCCCTCAGAAG | C. carboxidivorans Type III M2 |


| Methylation Motif | Type | Name | C. carboxidivorans | E. coli | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| % of Motifs Detected | # of Motifs Detected | # of Motifs in Genome | Mean QV | Mean Coverage | % of Motifs Detected | # of Motifs Detected | # of Motifs in Genome | Mean QV | Mean Coverage | |||
| CAYNNNNNCTGC a | I.1 | M.CcaP7I | 99.5 | 1047 | 1052 | 201.2 | 177.0 | 32.0 a | 91 a | 284 a | 62.1 a | 209.6 a |
| GCAGNNNNNRTG b | 99.5 | 1047 | 1052 | 212.9 | 177.0 | 20.6 b | 219 b | 1060 b | 59.4 b | 210.1 b | ||
| CCANNNNNNNNTCG | I.2 | M.CcaP7II | 99.1 | 241 | 243 | 207.1 | 182.1 | 99.6 | 3126 | 3137 | 188.0 | 142.4 |
| CGANNNNNNNNTGG | 98.7 | 240 | 243 | 211.3 | 182.9 | 98.7 | 3097 | 3137 | 177.6 | 143.1 | ||
| GCANNNNNNNTNNCG | I.3 | M.CcaP7III | 99.5 | 451 | 453 | 208.0 | 183.4 | 99.3 | 4075 | 4100 | 146.2 | 108.3 |
| CGNNANNNNNNNTGC | 99.5 | 451 | 453 | 201.3 | 183.3 | 99.4 | 4078 | 4100 | 144.9 | 108.0 | ||
| CTSAG * | II.1 | M.CcaP7ORF 16660P | - | - | - | - | - | - | - | - | - | - |
| GATAAT | II RM.1 | CcaP7IV | 84.8 | 6251 | 7367 | 152.9 | 179.7 | 98.2 | 3609 | 3672 | 248.7 | 211.1 |
| CRAAAAR | II RM.2 | CcaP7ORF 7120P | 97.3 | 6321 | 6490 | 166.7 | 177.8 | 99.3 | 4368 | 4395 | 247.5 | 257.0 |
| AGAAGC ** | III.1 | M.CcaP7ORF 17830P | 99.6 | 4629 | 4647 | 245.0 | 180.4 | - | - | - | - | - |
| GAAAT *** | III.2 | M.CcaP7ORF 8705P | - | - | - | - | - | 99.2 | 12,981 | 13,076 | 167.1 | 133.6 |

References
- Phillips, J.R.; Clausen, E.C.; Gaddy, J.L. Synthesis Gas as Substrate for the Biological Production of Fuels and Chemicals. Appl. Biochem. Biotechnol. 1994, 45/46, 145–157. [Google Scholar] [CrossRef]
- Köpke, M.; Held, C.; Hujer, S.; Liesegang, H.; Wiezer, A.; Wollherr, A.; Ehrenreich, A.; Liebl, W.; Gottschalk, G.; Durre, P. Clostridium ljungdahlii represents a microbial production platform based on syngas. Proc. Natl. Acad. Sci. USA 2010, 107, 13087–13092. [Google Scholar] [CrossRef] [PubMed]
- Liew, M.F.; Köpke, M.; Simpson, D.S. Gas Fermentation for Commercial Biofuels Production. In Liquid, Gaseous and Solid Biofuels; Fang, Z., Ed.; IntechOpen: London, UK, 2013. [Google Scholar] [CrossRef]
- Sun, X.; Atiyeh, H.K.; Huhnke, R.L.; Tanner, R.S. Syngas fermentation process development for production of biofuels and chemicals: A review. Bioresour. Technol. Rep. 2019, 7, 100279. [Google Scholar] [CrossRef]
- Fackler, N.; Heijstra, B.D.; Rasor, B.J.; Brown, H.; Martin, J.; Ni, Z.; Shebek, K.M.; Rosin, R.R.; Simpson, S.D.; Tyo, K.E.; et al. Stepping on the Gas to a Circular Economy: Accelerating Development of Carbon-Negative Chemical Production from Gas Fermentation. Annu. Rev. Chem. Biomol. Eng. 2021, 12, 439–470. [Google Scholar] [CrossRef] [PubMed]
- Shen, S.; Gu, Y.; Chai, C.; Jiang, W.; Zhuang, Y.; Wang, Y. Enhanced alcohol titre and ratio in carbon monoxide-rich off-gas fermentation of Clostridium carboxidivorans through combination of trace metals optimization with variable-temperature cultivation. Bioresour. Technol. 2017, 239, 236–243. [Google Scholar] [CrossRef] [PubMed]
- Kottenhahn, P.; Philipps, G.; Jennewein, S. Hexanol biosynthesis from syngas by Clostridium carboxidivorans P7—Product toxicity, temperature dependence and in situ extraction. Heliyon 2021, 7, e07732. [Google Scholar] [CrossRef] [PubMed]
- Oh, H.J.; Gong, G.; Ahn, J.H.; Ko, J.K.; Lee, S.-M.; Um, Y. Effective hexanol production from carbon monoxide using extractive fermentation with Clostridium carboxidivorans P7. Bioresour. Technol. 2023, 367, 128201. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.; Li, W.; Lin, M.; Yang, S.T. Metabolic engineering of Clostridium carboxidivorans for enhanced ethanol and butanol production from syngas and glucose. Bioresour. Technol. 2019, 284, 415–423. [Google Scholar] [CrossRef]
- Raleigh, E.A.; Murray, N.E.; Revel, H.; Blumenthal, R.M.; Westaway, D.; Reith, A.D.; Rigby, P.W.; Elhai, J.; Hanahan, D. McrA and McrB restriction phenotypes of some E. coli strains and implications for gene cloning. Nucleic Acids Res. 1988, 16, 1563–1575. [Google Scholar] [CrossRef]
- Pyne, M.E.; Moo-Young, M.; Chung, D.A.; Chou, C.P. Development of an electrotransformation protocol for genetic manipulation of Clostridium pasteurianum. Biotechnol. Biofuels 2013, 6, 50. [Google Scholar] [CrossRef]
- Pyne, M.E.; Bruder, M.; Moo-Young, M.; Chung, D.A.; Chou, C.P. Technical guide for genetic advancement of underdeveloped and intractable Clostridium. Biotechnol. Adv. 2014, 32, 623–641. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.-N.; Liebl, W.; Ehrenreich, A. Restriction-deficient mutants and marker-less genomic modification for metabolic engineering of the solvent producer Clostridium saccharobutylicum. Biotechnol. Biofuels 2018, 11, 264. [Google Scholar] [CrossRef] [PubMed]
- Woods, C.; Humphreys, C.M.; Rodrigues, R.M.; Ingle, P.; Rowe, P.; Henstra, A.M.; Kopke, M.; Simpson, S.D.; Winzer, K.; Minton, N.P. A Novel Conjugal Donor Strain for Improved DNA transfer into Clostridium spp. Anaerobe 2019, 59, 184–191. [Google Scholar] [CrossRef] [PubMed]
- Bernheim, A.; Sorek, R. The pan-immune system of bacteria: Antiviral defence as a community resource. Nat. Rev. Microbiol. 2020, 18, 113–119. [Google Scholar] [CrossRef]
- Seong, H.J.; Han, S.-W.; Sul, W.J. Prokaryotic DNA methylation and its functional roles. J. Microbiol. 2021, 59, 242–248. [Google Scholar] [CrossRef]
- Suzuki, H. Host-Mimicking Strategies in DNA Methylation for Improved Bacterial Transformation; INTECH Open Access Publisher: London, UK, 2012; ISBN 9535108816. [Google Scholar] [CrossRef]
- Murray, N.E. Type I restriction systems: Sophisticated molecular machines (a legacy of Bertani and Weigle). Microbiol. Mol. Biol. Rev. 2000, 64, 412–434. [Google Scholar] [CrossRef]
- Loenen, W.A.; Dryden, D.T.; Raleigh, E.A.; Wilson, G.G. Type I restriction enzymes and their relatives. Nucleic Acids Res. 2014, 42, 20–44. [Google Scholar] [CrossRef]
- Pingoud, A.; Wilson, G.G.; Wende, W. Type II restriction endonucleases--a historical perspective and more. Nucleic Acids Res. 2014, 42, 7489–7527. [Google Scholar] [CrossRef]
- Wilson, G.G. Organization of restriction-modification systems. Nucleic Acids Res. 1991, 19, 2539–2566. [Google Scholar] [CrossRef]
- Wilson, G.G.; Murray, N.E. Restriction and modification systems. Annu. Rev. Genet. 1991, 25, 585–627. [Google Scholar] [CrossRef]
- Loenen, W.A.M.; Raleigh, E.A. The other face of restriction: Modification-dependent enzymes. Nucleic Acids Res. 2014, 42, 56–69. [Google Scholar] [CrossRef]
- Kirk, J.A.; Fagan, R.P. Heat shock increases conjugation efficiency in Clostridium difficile. Anaerobe 2016, 42, 1–5. [Google Scholar] [CrossRef]
- Oliveira, P.H.; Ribis, J.W.; Garrett, E.M.; Trzilova, D.; Kim, A.; Sekulovic, O.; Mead, E.A.; Pak, T.; Zhu, S.; Deikus, G.; et al. Epigenomic characterization of Clostridioides difficile finds a conserved DNA methyltransferase that mediates sporulation and pathogenesis. Nat. Microbiol. 2020, 5, 166–180. [Google Scholar] [CrossRef]
- Johnston, C.D.; Cotton, S.L.; Rittling, S.R.; Starr, J.R.; Borisy, G.G.; Dewhirst, F.E.; Lemon, K.P. Systematic evasion of the restriction-modification barrier in bacteria. Proc. Natl. Acad. Sci. USA 2019, 116, 11454–11459. [Google Scholar] [CrossRef]
- Riley, L.A.; Ji, L.; Schmitz, R.J.; Westpheling, J.; Guss, A.M. Rational development of transformation in Clostridium thermocellum ATCC 27405 via complete methylome analysis and evasion of native restriction-modification systems. J. Ind. Microbiol. Biotechnol. 2019, 46, 1435–1443. [Google Scholar] [CrossRef]
- Eid, J.; Fehr, A.; Gray, J.; Luong, K.; Lyle, J.; Otto, G.; Peluso, P.; Rank, D.; Baybayan, P.; Bettman, B.; et al. Real-time DNA sequencing from single polymerase molecules. Science 2009, 323, 133–138. [Google Scholar] [CrossRef]
- Flusberg, B.A.; Webster, D.R.; Lee, J.H.; Travers, K.J.; Olivares, E.C.; Clark, T.A.; Korlach, J.; Turner, S.W. Direct detection of DNA methylation during single-molecule, real-time sequencing. Nat. Methods 2010, 7, 461–465. [Google Scholar] [CrossRef]
- Feng, Z.; Fang, G.; Korlach, J.; Clark, T.; Luong, K.; Zhang, X.; Wong, W.; Schadt, E. Detecting DNA Modifications from SMRT Sequencing Data by Modeling Sequence Context Dependence of Polymerase Kinetic. PLoS Comput. Biol. 2013, 9, e1002935. [Google Scholar] [CrossRef]
- Roberts, R.J.; Vincze, T.; Posfai, J.; Macelis, D. REBASE—A database for DNA restriction and modification: Enzymes, genes and genomes. Nucleic Acids Res. 2015, 43, D298–D299. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Yang, J.; Chai, C.; Yang, S.; Jiang, W.; Gu, Y. Complete genome sequence of Clostridium carboxidivorans P7(T), a syngas-fermenting bacterium capable of producing long-chain alcohols. J. Biotechnol. 2015, 211, 44–45. [Google Scholar] [CrossRef] [PubMed]
- Biosciences, P. Detecting DNA base modifications using single molecule, real-time sequencing. White Pap. Base Modif. 2015. Available online: https://www.pacb.com/wp-content/uploads/2015/09/WP_Detecting_DNA_Base_Modifications_Using_SMRT_Sequencing.pdf (accessed on 24 February 2023).
- Dong, M.-J.; Luo, H.; Gao, F. Ori-Finder 2022: A Comprehensive Web Server for Prediction and Analysis of Bacterial Replication Origins. Genom. Proteom. Bioinform. 2022, 20, 1207–1213. [Google Scholar] [CrossRef]
- McGlinchey, A.S.; Zepeda-Rivera, M.A.; Stepanovica, M.; Baryiames, A.A.; Jones, D.S.; LaCourse, K.D.; Bullman, S.; Johnston, C.D. Complete Genome Sequence of Clostridium cadaveris IFB3C5, Isolated from a Human Colonic Adenocarcinoma. Microbiol. Resour. Announc. 2022, 11, e0113521. [Google Scholar] [CrossRef]
- Philipps, G.; de Vries, S.; Jennewein, S. Development of a metabolic pathway transfer and genomic integration system for the syngas-fermenting bacterium Clostridium ljungdahlii. Biotechnol. Biofuels 2019, 12, 112. [Google Scholar] [CrossRef]
- Pregnon, G.; Minton, N.P.; Soucaille, P. Genome Sequence of Eubacterium limosum B2 and Evolution for Growth on a Mineral Medium with Methanol and CO2 as Sole Carbon Sources. Microorganisms 2022, 10, 1790. [Google Scholar] [CrossRef]
- Utturkar, S.M.; Klingeman, D.M.; Bruno-Barcena, J.M.; Chinn, M.S.; Grunden, A.M.; Köpke, M.; Brown, S.D. Sequence data for Clostridium autoethanogenum using three generations of sequencing technologies. Sci. Data 2015, 2, 150014. [Google Scholar] [CrossRef]
- REBASE. Available online: http://rebase.neb.com (accessed on 24 February 2023).
- Jurenaite-Urbanaviciene, S.; Kazlauskiene, R.; Urbelyte, V.; Maneliene, Z.; Petruyte, M.; Lubys, A.; Janulaitis, A. Characterization of BseMII, a new type IV restriction–modification system, which recognizes the pentanucleotide sequence 5′-CTCAG(N)10/8↓. Nucleic Acids Res. 2001, 29, 895–903. [Google Scholar] [CrossRef]
- Minton, N.P.; Ehsaan, M.; Humphreys, C.M.; Little, G.T.; Baker, J.; Henstra, A.M.; Liew, F.; Kelly, M.L.; Sheng, L.; Schwarz, K.; et al. A roadmap for gene system development in Clostridium. Anaerobe 2016, 41, 104–112. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Xu, M.; Yang, S.-T. Restriction modification system analysis and development of in vivo methylation for the transformation of Clostridium cellulovorans. Appl. Microbiol. Biotechnol. 2016, 100, 2289–2299. [Google Scholar] [CrossRef] [PubMed]

| Type | Subunit | Gene Name | Predicted Recognition Site | Coordinates | ORF (bp) | Short Name |
|---|---|---|---|---|---|---|
| I | S | S.CcaP7I | CAYNNNNNCTGC | 2727394–2728617 c | 1224 | Type I.1 |
| M | M.CcaP7I | CAYNNNNNCTGC | 2728619–2730115 c | 1497 | ||
| R | CcaP7IP | CAYNNNNNCTGC | 2731721–2735029 c | 3309 | ||
| I | R | CcaP7IIP | CCANNNNNNNNTCG | 827150–830422 | 3273 | Type I.2 |
| M | M.CcaP7II | CCANNNNNNNNTCG | 830425–831846 | 1422 | ||
| S | S.CcaP7II | CCANNNNNNNNTCG | 831846–833189 | 1344 | ||
| I | R | CcaP7IIIP | GCANNNNNNNTNNCG | 5411396–5414605 | 3210 | Type I.3 |
| S | S.CcaP7III | GCANNNNNNNTNNCG | 5414864–5416072 | 1209 | ||
| M | M.CcaP7III | GCANNNNNNNTNNCG | 5416106–5417695 | 1590 | ||
| II | M | M.CcaP7ORF16660P | CTSAG | 3757440–3758204 c | 765 | Type II.1 |
| II | RM | CcaP7IV | GATAAT | 5299217–5300845 | 1629 | Type II RM.1 |
| II | RM | CcaP7ORF7120P | - | 1647209–1650934 | 3726 | Type II RM.2 |
| III | R | CcaP7ORF17830P | - | 4003447–4006389 c | 2943 | Type III.1 |
| M | M.CcaP7ORF17830P | 4006403–4008355 c | 1953 | |||
| III | R | CcaP7ORF8705P | - | 2023100–2025358 c | 2259 | Type III.2 |
| M | M.CcaP7ORF8705P | 2025352–2027229 c | 1878 | |||
| IV | R | CcaP7ORF3495P | - | 805395–808574 | 3180 | Type IV.1 |
| IV | R | CcaP7McrB2P | - | 815415–816887 | 1473 | Type IV.2 |
| IV | R | CcaP7McrBP | - | 816865–818568 | 1704 | Type IV.3 |
| Organism | Genome Size (bp) | Accession Number | PacBio | Putative REs | Putative MTases |
|---|---|---|---|---|---|
| Clostridium acetobutylicum ATCC 824 | 3,940,880 | AE001437 (NC_003030) | No | 3 | 6 |
| Clostridium autoethanogenum DSM 10061 | 4,352,446 | CP012395 | Yes | 6 | 4 |
| Clostridium beijerinckii NCIMB 8052 | 6,000,632 | CP000721 (NC_009617) | No | 2 | 2 |
| Clostridium botulinum A ATCC 19397 | 3,863,450 | CP000726 (NC_009697) | Yes | 2 | 5 |
| Clostridium carboxidivorans P7 | 5,732,880 | CP011803 | Yes | 10 | 8 |
| Clostridium cellulolyticum H10 | 4,068,724 | CP001348 (NC_011898) | No | 4 | 10 |
| Clostridium cellulovorans 743B | 5,262,222 | CP002160 (NC_014393) | No | 6 | 13 |
| Clostridium difficile 630 | 4,290,252 | AM180355 (NC_009089) | Yes | 2 | 5 |
| Clostridium diolis DSM 15410 | 5,940,808 | CP043998 | Yes | 2 | 1 |
| Clostridium kluyveri DSM 555 | 3,964,618 | CP000673 (NC_009706) | No | 5 | 13 (2 a) |
| Clostridium ljungdahlii DSM 13528 | 4,630,065 | CP001666 (NC_014328) | Yes | 5 | 7 |
| Clostridium pasteurianum BC1 | 4,990,707 | CP003261 | No | 4 | 5 |
| Clostridium pasteurianum DSM 525 = ATCC 6013 | 4,352,852 | CP013018 | Yes | 4 | 8 |
| Clostridium perfringens ATCC 13124 | 3,256,683 | CP000246 (NC_008261) | Yes | 6 | 7 |
| Clostridium sporogenes DSM 795 | 4,142,990 | CP011663 | Yes | 2 | 4 |
| Clostridium thermocellum ATCC 27405 | 3,843,301 | CP000568 (NC_009012) | Yes | 5 | 11 |
| Methylation Motif | Type | Name | Homology |
|---|---|---|---|
| CAYNNNNNCTGC | Type I.1 | M.CcaP7I | None |
| GCAGNNNNNRTG | |||
| CCANNNNNNNNTCG | Type I.2 | M.CcaP7II | Motif included in CCANNNNNNNNTCGT/ ACGANNNNNNNNTGG found in Vibrio harveyi NCTC12970 |
| CGANNNNNNNNTGG | |||
| GCANNNNNNNTNNCG | Type I.3 | M.CcaP7III | Motif included in GGCANNNNNNNTNNCG/ CGNNANNNNNNNTGC found in Klebsiella pneumoniae AR_0139 |
| CGNNANNNNNNNTGC | |||
| CTSAG * | Type II.1 | M.CcaP7ORF16660P | Predicted for many strains (including Clostridia) and several confirmed by PacBio, but also other species such as Klebsiella pneumoniae NCTC9151, Lactobacillus acidophilus, and Bacillus stearothermophilus Isl 15-111 (with gold standard enzyme) |
| GATAAT | Type II RM.1 | CcaP7IV | Several strains from Clostridium botulinum and Clostridium sporogenes |
| CRAAAAR | Type II RM.2 | CcaP7ORF7120P | Motif CRAAAAR: Clostridium cadaveris IFB3C5 a Motif CAAAAAR: several strains from Clostridium botulinum and Clostridium sporogenes; other species such as Clostridium pasteurianum, Clostridium tetani, Clostridium autoethanogenum, Clostridium ljungdahlii b, Eubacterium limosum B2 c and Acetobacterium woodii DSM 1030 Motif CAAAAA: Clostridium difficile 630 (with gold standard enzyme) |
| AGAAGC ** | Type III.1 | M.CcaP7ORF17830P | Lactococcus lactis subsp. lactis strain UC073 |
| GAAAT *** | Type III.2 | M.CcaP7ORF8705P | Pseudopropionibacterium propionicum NCTC11666 Corynebacterium diphtheriae NCTC10838 Helicobacter fennelliae NCTC13102 Arachnia propionica F0231 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kottenhahn, P.; Philipps, G.; Bunk, B.; Spröer, C.; Jennewein, S. The Restriction–Modification Systems of Clostridium carboxidivorans P7. Microorganisms 2023, 11, 2962. https://doi.org/10.3390/microorganisms11122962
Kottenhahn P, Philipps G, Bunk B, Spröer C, Jennewein S. The Restriction–Modification Systems of Clostridium carboxidivorans P7. Microorganisms. 2023; 11(12):2962. https://doi.org/10.3390/microorganisms11122962
Chicago/Turabian StyleKottenhahn, Patrick, Gabriele Philipps, Boyke Bunk, Cathrin Spröer, and Stefan Jennewein. 2023. "The Restriction–Modification Systems of Clostridium carboxidivorans P7" Microorganisms 11, no. 12: 2962. https://doi.org/10.3390/microorganisms11122962