
Natural Product Cordycepin (CD) Inhibition for NRP1/CD304 Expression and Possibly SARS-CoV-2 Susceptibility Prevention on Cancers
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Online Databases
2.2. Antibodies and Reagents
2.3. Cell Culture
2.4. CD Treatments and Isolation of Mouse Lymphocytes
2.5. Western Blotting
2.6. Semi-Quantitative RT-PCR
2.7. Molecular Docking
2.8. Cell Transfection and Syncytial Formation
2.9. Statistical Analysis
3. Results
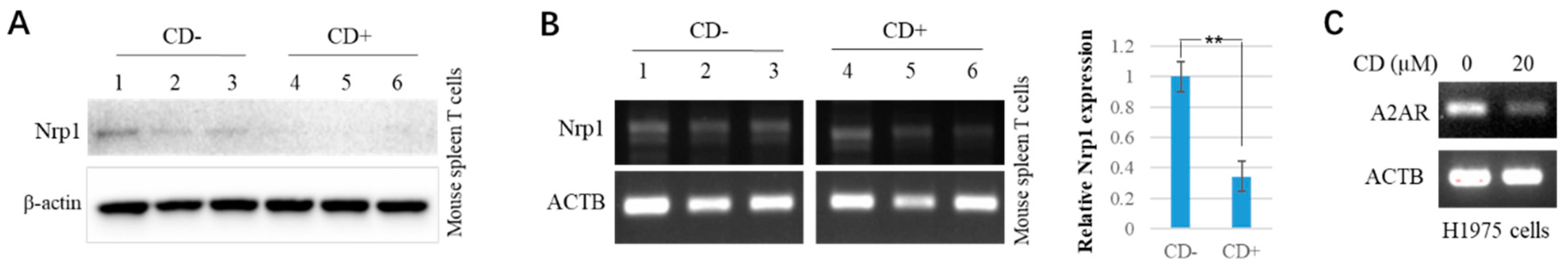
3.1. Cordycepin (CD) Inhibits NRP1 Expression in Various Cells
3.2. Docking and Molecular Interaction Study of CD with NRP1
3.3. CD Inhibits Syncytial Formation Likely through NRP1
3.4. CD Roles in Immune Molecules and NRP1 Expression Analysis on Correlated Genes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Takagi, S.; Tsuji, T.; Amagai, T.; Takamatsu, T.; Fujisawa, H. Specific cell surface labels in the visual centers of xenopus laevis tadpole identified using monoclonal antibodies. Dev. Biol. 1987, 122, 90–100. [Google Scholar] [CrossRef]
- Fujisawa, H.; Ohtsuki, T.; Takagi, S.; Tsuji, T. An aberrant retinal pathway and visual centers in xenopus tadpoles share a common cell surface molecule, a5 antigen. Dev. Biol. 1989, 135, 231–240. [Google Scholar] [CrossRef] [PubMed]
- Kolodkin, A.L.; Levengood, D.V.; Rowe, E.G.; Tai, Y.T.; Giger, R.J.; Ginty, D.D. Neuropilin is a semaphorin iii receptor. Cell 1997, 90, 753–762. [Google Scholar] [CrossRef] [PubMed]
- He, Z.; Tessier-Lavigne, M. Neuropilin is a receptor for the axonal chemorepellent semaphorin iii. Cell 1997, 90, 739–751. [Google Scholar] [CrossRef]
- Soker, S.; Takashima, S.; Miao, H.Q.; Neufeld, G.; Klagsbrun, M. Neuropilin-1 is expressed by endothelial and tumor cells as an isoform-specific receptor for vascular endothelial growth factor. Cell 1998, 92, 735–745. [Google Scholar] [CrossRef] [PubMed]
- Al-Zeheimi, N.; Gao, Y.; Greer, P.A.; Adham, S.A. Neuropilin-1 knockout and rescue confirms its role to promote metastasis in MDA-MB-231 breast cancer cells. Int. J. Mol. Sci. 2023, 24, 7792. [Google Scholar] [CrossRef] [PubMed]
- Cantuti-Castelvetri, L.; Ojha, R.; Pedro, L.D.; Djannatian, M.; Franz, J.; Kuivanen, S.; van der Meer, F.; Kallio, K.; Kaya, T.; Anastasina, M.; et al. Neuropilin-1 facilitates SARS-CoV-2 cell entry and infectivity. Science 2020, 370, 856–860. [Google Scholar] [CrossRef] [PubMed]
- Daly, J.L.; Simonetti, B.; Klein, K.; Chen, K.E.; Williamson, M.K.; Anton-Plagaro, C.; Shoemark, D.K.; Simon-Gracia, L.; Bauer, M.; Hollandi, R.; et al. Neuropilin-1 is a host factor for SARS-CoV-2 infection. Science 2020, 370, 861–865. [Google Scholar] [CrossRef]
- Mayi, B.S.; Leibowitz, J.A.; Woods, A.T.; Ammon, K.A.; Liu, A.E.; Raja, A. The role of neuropilin-1 in COVID-19. PLoS Pathog. 2021, 17, e1009153. [Google Scholar] [CrossRef]
- Li, T.; Fu, J.; Cheng, J.; Elfiky, A.A.; Wei, C.; Fu, J. New progresses on cell surface protein HSPA5/BIP/GRP78 in cancers and COVID-19. Front. Immunol. 2023, 14, 1166680. [Google Scholar] [CrossRef]
- Fu, J.; Zhou, B.; Zhang, L.; Balaji, K.S.; Wei, C.; Liu, X.; Chen, H.; Peng, J.; Fu, J. Expressions and significances of the angiotensin-converting enzyme 2 gene, the receptor of SARS-CoV-2 for COVID-19. Mol. Biol. Rep. 2020, 47, 4383–4392. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Horby, P.W.; Hayden, F.G.; Gao, G.F. A novel coronavirus outbreak of global health concern. Lancet 2020, 395, 470–473. [Google Scholar] [CrossRef] [PubMed]
- Coutard, B.; Valle, C.; de Lamballerie, X.; Canard, B.; Seidah, N.G.; Decroly, E. The spike glycoprotein of the new coronavirus 2019-nCoV contains a furin-like cleavage site absent in CoV of the same clade. Antiviral. Res. 2020, 176, 104742. [Google Scholar] [CrossRef]
- Katopodis, P.; Randeva, H.S.; Spandidos, D.A.; Saravi, S.; Kyrou, I.; Karteris, E. Host cell entry mediators implicated in the cellular tropism of SARSCoV2, the pathophysiology of COVID19 and the identification of microRNAs that can modulate the expression of these mediators (review). Int. J. Mol. Med. 2022, 49, 20. [Google Scholar] [CrossRef] [PubMed]
- Ishitoku, M.; Mokuda, S.; Araki, K.; Watanabe, H.; Kohno, H.; Sugimoto, T.; Yoshida, Y.; Sakaguchi, T.; Masumoto, J.; Hirata, S.; et al. Tumor necrosis factor and interleukin-1beta upregulate NRP2 expression and promote SARS-CoV-2 proliferation. Viruses 2023, 15, 1498. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Zhao, L.; Zhang, X.; Zhang, J.; Shang, H.; Liang, G. Neuropilin-1, a myeloid cell-specific protein, is an inhibitor of hiv-1 infectivity. Proc. Natl. Acad. Sci. USA 2022, 119, e2114884119. [Google Scholar] [CrossRef] [PubMed]
- Adimulam, T.; Arumugam, T.; Naidoo, A.; Naidoo, K.; Ramsuran, V. Polymorphisms within the SARS-CoV-2 human receptor genes associate with variable disease outcomes across ethnicities. Genes 2023, 14, 1798. [Google Scholar] [CrossRef]
- Adimulam, T.; Arumugam, T.; Gokul, A.; Ramsuran, V. Genetic variants within SARS-CoV-2 human receptor genes may contribute to variable disease outcomes in different ethnicities. Int. J. Mol. Sci. 2023, 24, 8711. [Google Scholar] [CrossRef] [PubMed]
- Pal, D.; De, K.; Yates, T.B.; Kolape, J.; Muchero, W. Mutating novel interaction sites in nrp1 reduces SARS-CoV-2 spike protein internalization. iScience 2023, 26, 106274. [Google Scholar] [CrossRef]
- Ozkan Oktay, E.; Kaman, T.; Karasakal, O.F.; Enisoglu Atalay, V. In silico prediction and molecular docking of SNPs in nrp1 gene associated with SARS-CoV-2. Biochem. Genet. 2023, 1–20. [Google Scholar] [CrossRef]
- Chapoval, S.P.; Keegan, A.D. Perspectives and potential approaches for targeting neuropilin 1 in SARS-CoV-2 infection. Mol. Med. 2021, 27, 162. [Google Scholar] [CrossRef]
- Ackermann, M.; Verleden, S.E.; Kuehnel, M.; Haverich, A.; Welte, T.; Laenger, F.; Vanstapel, A.; Werlein, C.; Stark, H.; Tzankov, A.; et al. Pulmonary vascular endothelialitis, thrombosis, and angiogenesis in COVID-19. N. Engl. J. Med. 2020, 383, 120–128. [Google Scholar] [CrossRef]
- Mercurio, A.M. Vegf/neuropilin signaling in cancer stem cells. Int. J. Mol. Sci. 2019, 20, 490. [Google Scholar] [CrossRef] [PubMed]
- Rachner, T.D.; Kasimir-Bauer, S.; Goebel, A.; Erdmann, K.; Hoffmann, O.; Rauner, M.; Hofbauer, L.C.; Kimmig, R.; Bittner, A.K. Soluble neuropilin-1 is an independent marker of poor prognosis in early breast cancer. J. Cancer Res. Clin. Oncol. 2021, 147, 2233–2238. [Google Scholar] [CrossRef] [PubMed]
- Nasarre, C.; Roth, M.; Jacob, L.; Roth, L.; Koncina, E.; Thien, A.; Labourdette, G.; Poulet, P.; Hubert, P.; Cremel, G.; et al. Peptide-based interference of the transmembrane domain of neuropilin-1 inhibits glioma growth in vivo. Oncogene 2010, 29, 2381–2392. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.B.; Zhang, H.; Zhang, J.P.; Li, Y.; Zhao, B.; Feng, G.K.; Du, Y.; Xiong, D.; Zhong, Q.; Liu, W.L.; et al. Neuropilin 1 is an entry factor that promotes ebv infection of nasopharyngeal epithelial cells. Nat. Commun. 2015, 6, 6240. [Google Scholar] [CrossRef]
- Chen, M.; Wang, M.H.; Shen, X.G.; Liu, H.; Zhang, Y.Y.; Peng, J.M.; Meng, F.; Wang, T.Y.; Bai, Y.Z.; Sun, M.X.; et al. Neuropilin-1 facilitates pseudorabies virus replication and viral glycoprotein b promotes its degradation in a furin-dependent manner. J. Virol. 2022, 96, e0131822. [Google Scholar] [CrossRef]
- Lane, R.K.; Guo, H.; Fisher, A.D.; Diep, J.; Lai, Z.; Chen, Y.; Upton, J.W.; Carette, J.; Mocarski, E.S.; Kaiser, W.J. Necroptosis-based crispr knockout screen reveals neuropilin-1 as a critical host factor for early stages of murine cytomegalovirus infection. Proc. Natl. Acad. Sci. USA 2020, 117, 20109–20116. [Google Scholar] [CrossRef]
- Ghez, D.; Lepelletier, Y.; Lambert, S.; Fourneau, J.M.; Blot, V.; Janvier, S.; Arnulf, B.; van Endert, P.M.; Heveker, N.; Pique, C.; et al. Neuropilin-1 is involved in human t-cell lymphotropic virus type 1 entry. J. Virol. 2006, 80, 6844–6854. [Google Scholar] [CrossRef]
- Kolaric, A.; Jukic, M.; Bren, U. Novel small-molecule inhibitors of the SARS-CoV-2 spike protein binding to neuropilin 1. Pharmaceuticals 2022, 15, 165. [Google Scholar] [CrossRef]
- Charoute, H.; Elkarhat, Z.; Elkhattabi, L.; El Fahime, E.; Oukkache, N.; Rouba, H.; Barakat, A. Computational screening of potential drugs against COVID-19 disease: The neuropilin-1 receptor as molecular target. Virusdisease 2022, 33, 23–31. [Google Scholar] [CrossRef] [PubMed]
- Alshawaf, E.; Hammad, M.M.; Marafie, S.K.; Ali, H.; Al-Mulla, F.; Abubaker, J.; Mohammad, A. Discovery of natural products to block SARS-CoV-2 s-protein interaction with neuropilin-1 receptor: A molecular dynamics simulation approach. Microb. Pathog. 2022, 170, 105701. [Google Scholar] [CrossRef] [PubMed]
- Ganguly, A.; Mandi, M.; Dutta, A.; Rajak, P. In silico analysis reveals the inhibitory potential of madecassic acid against entry factors of SARS-CoV-2. ACS Appl. Bio Mater. 2023, 6, 652–662. [Google Scholar] [CrossRef]
- Karkashan, A.; Attar, R. Computational screening of natural products to identify potential inhibitors for human neuropilin-1 (nrp1) receptor to abrogate the binding of SARS-CoV-2 and host cell. J. Biomol. Struct. Dyn. 2023, 41(19), 9987–9996. [Google Scholar] [CrossRef] [PubMed]
- Skrbic, R.; Travar, M.; Stojiljkovic, M.P.; Djuric, D.M.; Surucic, R. Folic acid and leucovorin have potential to prevent SARS-CoV-2-virus internalization by interacting with s-glycoprotein/neuropilin-1 receptor complex. Molecules 2023, 28, 2294. [Google Scholar] [CrossRef]
- Hashizume, M.; Takashima, A.; Ono, C.; Okamoto, T.; Iwasaki, M. Phenothiazines inhibit SARS-CoV-2 cell entry via a blockade of spike protein binding to neuropilin-1. Antiviral Res. 2023, 209, 105481. [Google Scholar] [CrossRef]
- Perez-Miller, S.; Patek, M.; Moutal, A.; Duran, P.; Cabel, C.R.; Thorne, C.A.; Campos, S.K.; Khanna, R. Novel compounds targeting neuropilin receptor 1 with potential to interfere with SARS-CoV-2 virus entry. ACS Chem. Neurosci. 2021, 12, 1299–1312. [Google Scholar] [CrossRef]
- Li, D.; Liu, X.; Zhang, L.; He, J.; Chen, X.; Liu, S.; Fu, J.; Fu, S.; Chen, H.; Fu, J.; et al. COVID-19 disease and malignant cancers: The impact for the furin gene expression in susceptibility to SARS-CoV-2. Int. J. Biol. Sci. 2021, 17, 3954–3967. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Luo, J.; Jiang, W.; Chen, L.; Miao, L.; Han, C. Cordycepin: A review of strategies to improve the bioavailability and efficacy. Phytother. Res. 2023, 37, 3839–3858. [Google Scholar] [CrossRef]
- Wei, C.; Khan, M.A.; Du, J.; Cheng, J.; Tania, M.; Leung, E.L.; Fu, J. Cordycepin inhibits triple-negative breast cancer cell migration and invasion by regulating emt-tfs slug, twist1, snail1, and zeb1. Front. Oncol. 2022, 12, 898583. [Google Scholar] [CrossRef]
- Wei, C.; Yao, X.; Jiang, Z.; Wang, Y.; Zhang, D.; Chen, X.; Fan, X.; Xie, C.; Cheng, J.; Fu, J.; et al. Cordycepin inhibits drug-resistance non-small cell lung cancer progression by activating ampk signaling pathway. Pharmacol. Res. 2019, 144, 79–89. [Google Scholar] [CrossRef]
- He, J.; Liu, S.; Tan, Q.; Liu, Z.; Fu, J.; Li, T.; Wei, C.; Liu, X.; Mei, Z.; Cheng, J.; et al. Antiviral potential of small molecules cordycepin, thymoquinone, and n6, n6-dimethyladenosine targeting SARS-CoV-2 entry protein adam17. Molecules 2022, 27, 9044. [Google Scholar] [CrossRef] [PubMed]
- Fu, J.; Liu, S.; Tan, Q.; Liu, Z.; Qian, J.; Li, T.; Du, J.; Song, B.; Li, D.; Zhang, L.; et al. Impact of tmprss2 expression, mutation prognostics, and small molecule (cd, ad, tq, and tqfl12) inhibition on pan-cancer tumors and susceptibility to SARS-CoV-2. Molecules 2022, 27, 7413. [Google Scholar] [CrossRef] [PubMed]
- Ru, B.; Wong, C.N.; Tong, Y.; Zhong, J.Y.; Zhong, S.S.W.; Wu, W.C.; Chu, K.C.; Wong, C.Y.; Lau, C.Y.; Chen, I.; et al. Tisidb: An integrated repository portal for tumor-immune system interactions. Bioinformatics 2019, 35, 4200–4202. [Google Scholar] [CrossRef] [PubMed]
- Untergasser, A.; Cutcutache, I.; Koressaar, T.; Ye, J.; Faircloth, B.C.; Remm, M.; Rozen, S.G. Primer3—New capabilities and interfaces. Nucleic Acids Res. 2012, 40, e115. [Google Scholar] [CrossRef]
- Kubra, S.; Zhang, H.; Si, Y.; Gao, X.; Wang, T.; Pan, L.; Li, L.; Zhong, N.; Fu, J.; Zhang, B.; et al. Reggamma regulates circadian clock by modulating bmal1 protein stability. Cell Death Discov. 2021, 7, 335. [Google Scholar] [CrossRef]
- Wei, C.; Liu, Y.; Liu, X.; Cheng, J.; Fu, J.; Xiao, X.; Moses, R.E.; Li, X.; Fu, J. The speckle-type poz protein (spop) inhibits breast cancer malignancy by destabilizing twist1. Cell Death Discov. 2022, 8, 389. [Google Scholar] [CrossRef]
- Zhang, L.; Wei, C.; Li, D.; He, J.; Liu, S.; Deng, H.; Cheng, J.; Du, J.; Liu, X.; Chen, H.; et al. COVID-19 receptor and malignant cancers: Association of ctsl expression with susceptibility to SARS-CoV-2. Int. J. Biol. Sci. 2022, 18, 2362–2371. [Google Scholar] [CrossRef]
- Liu, G.; Du, W.; Sang, X.; Tong, Q.; Wang, Y.; Chen, G.; Yuan, Y.; Jiang, L.; Cheng, W.; Liu, D.; et al. Rna g-quadruplex in tmprss2 reduces SARS-CoV-2 infection. Nat. Commun. 2022, 13, 1444. [Google Scholar] [CrossRef]
- Kim, S.; Thiessen, P.A.; Bolton, E.E.; Chen, J.; Fu, G.; Gindulyte, A.; Han, L.; He, J.; He, S.; Shoemaker, B.A.; et al. Pubchem substance and compound databases. Nucleic Acids Res. 2016, 44, D1202–D1213. [Google Scholar] [CrossRef]
- Goodsell, D.S.; Olson, A.J. Automated docking of substrates to proteins by simulated annealing. Proteins 1990, 8, 195–202. [Google Scholar] [CrossRef]
- Jocher, G.; Grass, V.; Tschirner, S.K.; Riepler, L.; Breimann, S.; Kaya, T.; Oelsner, M.; Hamad, M.S.; Hofmann, L.I.; Blobel, C.P.; et al. Adam10 and adam17 promote SARS-CoV-2 cell entry and spike protein-mediated lung cell fusion. EMBO Rep. 2022, 23, e54305. [Google Scholar] [CrossRef] [PubMed]
- Braga, L.; Ali, H.; Secco, I.; Chiavacci, E.; Neves, G.; Goldhill, D.; Penn, R.; Jimenez-Guardeno, J.M.; Ortega-Prieto, A.M.; Bussani, R.; et al. Drugs that inhibit tmem16 proteins block SARS-CoV-2 spike-induced syncytia. Nature 2021, 594, 88–93. [Google Scholar] [CrossRef] [PubMed]
- Cekic, C.; Linden, J. Purinergic regulation of the immune system. Nat. Rev. Immunol. 2016, 16, 177–192. [Google Scholar] [CrossRef] [PubMed]
- Fong, L.; Hotson, A.; Powderly, J.D.; Sznol, M.; Heist, R.S.; Choueiri, T.K.; George, S.; Hughes, B.G.M.; Hellmann, M.D.; Shepard, D.R.; et al. Adenosine 2a receptor blockade as an immunotherapy for treatment-refractory renal cell cancer. Cancer Discov. 2020, 10, 40–53. [Google Scholar] [CrossRef] [PubMed]
- Novitskiy, S.V.; Ryzhov, S.; Zaynagetdinov, R.; Goldstein, A.E.; Huang, Y.; Tikhomirov, O.Y.; Blackburn, M.R.; Biaggioni, I.; Carbone, D.P.; Feoktistov, I.; et al. Adenosine receptors in regulation of dendritic cell differentiation and function. Blood 2008, 112, 1822–1831. [Google Scholar] [CrossRef]
- Liu, J.; Shi, Y.; Liu, X.; Zhang, D.; Bai, Y.; Xu, Y.; Wang, M. Blocking adenosine/A2AR pathway for cancer therapy. Zhongguo Fei Ai Za Zhi 2022, 25, 460–467. [Google Scholar]
- Zhou, X.; Luo, L.; Dressel, W.; Shadier, G.; Krumbiegel, D.; Schmidtke, P.; Zepp, F.; Meyer, C.U. Cordycepin is an immunoregulatory active ingredient of cordyceps sinensis. Am. J. Chin. Med. 2008, 36, 967–980. [Google Scholar] [CrossRef]
- Cunningham, K.G.; Manson, W.; Spring, F.S.; Hutchinson, S.A. Cordycepin, a metabolic product isolated from cultures of Cordyceps militaris (linn.) link. Nature 1950, 166, 949. [Google Scholar] [CrossRef]
- Rabie, A.M. Potent inhibitory activities of the adenosine analogue cordycepin on SARS-CoV-2 replication. ACS Omega 2022, 7, 2960–2969. [Google Scholar] [CrossRef]
- Radhi, M.; Ashraf, S.; Lawrence, S.; Tranholm, A.A.; Wellham, P.A.D.; Hafeez, A.; Khamis, A.S.; Thomas, R.; McWilliams, D.; de Moor, C.H. A systematic review of the biological effects of cordycepin. Molecules 2021, 26, 5886. [Google Scholar] [CrossRef]
- Shi, L.; Cao, H.; Fu, S.; Jia, Z.; Lu, X.; Cui, Z.; Yu, D. Cordycepin enhances hyperthermia-induced apoptosis and cell cycle arrest by modulating the mapk pathway in human lymphoma u937 cells. Mol. Biol. Rep. 2022, 49, 8673–8683. [Google Scholar] [CrossRef] [PubMed]
- Tuli, H.S.; Sharma, A.K.; Sandhu, S.S.; Kashyap, D. Cordycepin: A bioactive metabolite with therapeutic potential. Life Sci. 2013, 93, 863–869. [Google Scholar] [CrossRef] [PubMed]
- Qin, P.; Li, X.; Yang, H.; Wang, Z.Y.; Lu, D. Therapeutic potential and biological applications of cordycepin and metabolic mechanisms in cordycepin-producing fungi. Molecules 2019, 24, 2231. [Google Scholar] [CrossRef]
- Tuli, H.S.; Sandhu, S.S.; Sharma, A.K. Pharmacological and therapeutic potential of cordyceps with special reference to cordycepin. 3 Biotech 2014, 4, 1–12. [Google Scholar] [CrossRef]
- Bibi, S.; Hasan, M.M.; Wang, Y.B.; Papadakos, S.P.; Yu, H. Cordycepin as a promising inhibitor of SARS-CoV-2 RNA dependent rna polymerase (rdrp). Curr. Med. Chem. 2022, 29, 152–162. [Google Scholar] [CrossRef]
- Wang, Z.; Wang, N.; Yang, L.; Song, X.Q. Bioactive natural products in COVID-19 therapy. Front. Pharmacol. 2022, 13, 926507. [Google Scholar] [CrossRef]
- Fu, J.; Song, B.; Du, J.; Liu, S.; He, J.; Xiao, T.; Zhou, B.; Li, D.; Liu, X.; He, T.; et al. Impact of bsg/cd147 gene expression on diagnostic, prognostic and therapeutic strategies towards malignant cancers and possible susceptibility to SARS-CoV-2. Mol. Biol. Rep. 2023, 50, 2269–2281. [Google Scholar] [CrossRef]
- Powell, J.; Mota, F.; Steadman, D.; Soudy, C.; Miyauchi, J.T.; Crosby, S.; Jarvis, A.; Reisinger, T.; Winfield, N.; Evans, G.; et al. Small molecule neuropilin-1 antagonists combine antiangiogenic and antitumor activity with immune modulation through reduction of transforming growth factor beta (tgfbeta) production in regulatory t-cells. J. Med. Chem. 2018, 61, 4135–4154. [Google Scholar] [CrossRef]
- Hou, D.; Cao, W.; Kim, S.; Cui, X.; Ziarnik, M.; Im, W.; Zhang, X.F. Biophysical investigation of interactions between SARS-CoV-2 spike protein and neuropilin-1. Protein Sci. 2023, 32, e4773. [Google Scholar] [CrossRef] [PubMed]
- Hossain, M.G.; Akter, S.; Uddin, M.J. Emerging role of neuropilin-1 and angiotensin-converting enzyme-2 in renal carcinoma-associated COVID-19 pathogenesis. Infect. Dis. Rep. 2021, 13, 902–909. [Google Scholar] [CrossRef] [PubMed]
- Gold, S.A.; Margulis, V. Uncovering a link between COVID-19 and renal cell carcinoma. Nat. Rev. Urol. 2023, 20, 330–331. [Google Scholar] [CrossRef] [PubMed]
- Zalpoor, H.; Shapourian, H.; Akbari, A.; Shahveh, S.; Haghshenas, L. Increased neuropilin-1 expression by COVID-19: A possible cause of long-term neurological complications and progression of primary brain tumors. Hum. Cell 2022, 35, 1301–1303. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Primers | Sequence (from 5′-3′) | GenBank No. | Size (bp) |
|---|---|---|---|---|
| Cd28 | RT-mCD28-L | acaacgagaggagcaatgga | NM_007642.4 | 401 |
| RT-mCD28-R | gcccagtagaggtccaaagt | |||
| Cxcl12 | RT-mCXCL12-L | ctttcactctcggtccacct | NM_001012477.2 | 258 |
| RT-mCXCL12-R | gcaacaatctgaagggcaca | |||
| Csf1r | RT-mCSF1R-L | gcctcttcctctgttccctt | NM_001037859.2 | 372 |
| RT-mCSF1R-R | attcagggtccaaggtccag | |||
| Kdr | RT-mKDR-L | ggagtctgtgcctgagaact | NM_010612.3 | 440 |
| RT-mKDR-R | acagaggcgatgaatggtga | |||
| Ccr1 | RT-mCCR1-L | ttggaaccagagagaagccg | NM_001295.3 | 259 |
| RT-mCCR1-R | agaaatggccaggttcagga | |||
| Il2ra | RT-mIL2RA-L | acaagaacggcaccatccta | NM_008367.3 | 525 |
| RT-mIL2RA-R | agtctgtggtggttatgggg |
| Protein−Ligand Complex | Binding Affinity (kcal/mol) | NRP-1-Contacting Residues |
|---|---|---|
| CD-NRP1(b1b2) | −6.3 # | Lys105, Leu131, Pro281 *, Glu282 |
| CD-NRP1(b1) | −6.5 # | Tyr297, Asn300 *, Trp301, Asp320 *, Thr349 *, Tyr353 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, T.; Luo, N.; Fu, J.; Du, J.; Liu, Z.; Tan, Q.; Zheng, M.; He, J.; Cheng, J.; Li, D.; et al. Natural Product Cordycepin (CD) Inhibition for NRP1/CD304 Expression and Possibly SARS-CoV-2 Susceptibility Prevention on Cancers. Microorganisms 2023, 11, 2953. https://doi.org/10.3390/microorganisms11122953
Li T, Luo N, Fu J, Du J, Liu Z, Tan Q, Zheng M, He J, Cheng J, Li D, et al. Natural Product Cordycepin (CD) Inhibition for NRP1/CD304 Expression and Possibly SARS-CoV-2 Susceptibility Prevention on Cancers. Microorganisms. 2023; 11(12):2953. https://doi.org/10.3390/microorganisms11122953
Chicago/Turabian StyleLi, Ting, Na Luo, Jiewen Fu, Jiaman Du, Zhiying Liu, Qi Tan, Meiling Zheng, Jiayue He, Jingliang Cheng, Dabing Li, and et al. 2023. "Natural Product Cordycepin (CD) Inhibition for NRP1/CD304 Expression and Possibly SARS-CoV-2 Susceptibility Prevention on Cancers" Microorganisms 11, no. 12: 2953. https://doi.org/10.3390/microorganisms11122953