
Encystation and Stress Responses under the Control of Ubiquitin-like Proteins in Pathogenic Amoebae
 , ,
, , {kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Stress Responses Common in Eukaryotes
3. Mechanisms of Stress in Pathogenic Amoebae during Encystation
4. Mechanisms of Stress in Pathogenic Amoebae during Accumulation of Reactive Oxygen Species
5. UPR and Endoplasmic Reticulum Stress Responses Identified in Pathogenic Amoebae
6. Ubiquitin, Ubiquitin-like Protein Modifications, and Proteasome Activities in Pathogenic Amoebae
6.1. Autophagy-Related Proteins in Pathogenic Amoebae
6.2. UFM1, URM1 in Amoebae, a Terra Incognita
7. Conclusions and Perspectives
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Manna, D.; Ehrenkaufer, G.M.; Lozano-Amado, D.; Singh, U. Entamoeba Stage Conversion: Progress and New Insights. Curr. Opin. Microbiol. 2020, 58, 62–68. [Google Scholar] [CrossRef] [PubMed]
- Bínová, E.; Bína, D.; Nohýnková, E. DNA Content in Acanthamoeba during Two Stress Defense Reactions: Encystation, Pseudocyst Formation and Cell Cycle. Eur. J. Protistol. 2021, 77, 125745. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Jiang, L.; Zhao, Y.; Ju, X.; Wang, L.; Jin, L.; Fine, R.D.; Li, M. Biological Characteristics and Pathogenicity of Acanthamoeba. Front. Microbiol. 2023, 14, 1147077. [Google Scholar] [CrossRef] [PubMed]
- Legendre, M.; Lartigue, A.; Bertaux, L.; Jeudy, S.; Bartoli, J.; Lescot, M.; Alempic, J.-M.; Ramus, C.; Bruley, C.; Labadie, K.; et al. In-Depth Study of Mollivirus sibericum, a New 30,000-y-Old Giant Virus Infecting Acanthamoeba. Proc. Natl. Acad. Sci. USA 2015, 112, E5327–E5335. [Google Scholar] [CrossRef]
- Visvesvara, G.S.; Moura, H.; Schuster, F.L. Pathogenic and Opportunistic Free-Living Amoebae: Acanthamoeba Spp., Balamuthia mandrillaris, Naegleria fowleri, and Sappinia diploidea. FEMS Immunol. Med. Microbiol. 2007, 50, 1–26. [Google Scholar] [CrossRef]
- Lloyd, D. Encystment in Acanthamoeba Castellanii: A Review. Exp. Parasitol. 2014, 145, S20–S27. [Google Scholar] [CrossRef]
- Khunkitti, W.; Lloyd, D.; Furr, J.R.; Russell, A.D. Aspects of the Mechanisms of Action of Biguanides on Trophozoites and Cysts of Acanthamoeba Castellanii. J. Appl. Microbiol. 1997, 82, 107–114. [Google Scholar] [CrossRef]
- Mafra CS, P.; Carrijo-Carvalho, L.C.; Chudzinski-Tavassi, A.M.; de Carvalho Taguchi, F.M.; Foronda, A.S.; de Souza Carvalho, F.R.; de Freitas, D. Antimicrobial Action of Biguanides on the Viability of Acanthamoeba Cysts and Assessment of Cell Toxicity. Investig. Ophthalmol. Vis. Sci. 2013, 54, 6363–6372. [Google Scholar] [CrossRef]
- Hargrave, S.L.; McCulley, J.P.; Husseini, Z. Results of a Trial of Combined Propamidine Isethionate and Neomycin Therapy for Acanthamoeba Keratitis. Ophthalmology 1999, 106, 952–957. [Google Scholar] [CrossRef]
- Polat, Z.A.; Walochnik, J.; Obwaller, A.; Vural, A.; Dursun, A.; Arici, M.K. Miltefosine and Polyhexamethylene Biguanide: A New Drug Combination for the Treatment of Acanthamoeba keratitis. Clin. Exp. Ophthalmol. 2014, 42, 151–158. [Google Scholar] [CrossRef]
- Hiti, K.; Walochnik, J.; Faschinger, C.; Haller-Schober, E.-M.; Aspöck, H. One- and Two-Step Hydrogen Peroxide Contact Lens Disinfection Solutions against Acanthamoeba: How Effective Are They? Eye 2005, 19, 1301–1305. [Google Scholar] [CrossRef] [PubMed]
- Labruyère, E.; Thibeaux, R.; Olivo-Marin, J.-C.; Guillén, N. Crosstalk between Entamoeba Histolytica and the Human Intestinal Tract during Amoebiasis. Parasitology 2019, 146, 1140–1149. [Google Scholar] [CrossRef] [PubMed]
- Guillén, N. Pathogenicity and Virulence of Entamoeba Histolytica, the Agent of Amoebiasis. Virulence 2023, 14, 2158656. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Sun, H.; Liu, D.; Jiang, X.; Zheng, M.; Zhu, W.; Liu, Q.; Zheng, W.; Feng, X. Ac-HSP20 Is Associated with the Infectivity and Encystation of Acanthamoeba Castellanii. Front. Microbiol. 2021, 11, 595080. [Google Scholar] [CrossRef] [PubMed]
- Field, J.; Van Dellen, K.; Ghosh, S.K.; Samuelson, J. Responses of Entamoeba Invadens to Heat Shock and Encystation Are Related. J. Eukaryot. Microbiol. 2000, 47, 511–514. [Google Scholar] [CrossRef]
- Manna, D.; Lozano-Amado, D.; Ehrenkaufer, G.; Singh, U. The NAD+ Responsive Transcription Factor ERM-BP Functions Downstream of Cellular Aggregation and Is an Early Regulator of Development and Heat Shock Response in Entamoeba. Front. Cell. Infect. Microbiol. 2020, 10, 363. [Google Scholar] [CrossRef]
- Mornico, D.; Hon, C.-C.; Koutero, M.; Weber, C.; Coppee, J.-Y.; Dillies, M.-A.; Guillen, N. Genomic Determinants for Initiation and Length of Natural Antisense Transcripts in Entamoeba Histolytica. Sci. Rep. 2020, 10, 20190. [Google Scholar] [CrossRef]
- Bernard, C.; Locard-Paulet, M.; Noël, C.; Duchateau, M.; Giai Gianetto, Q.; Moumen, B.; Rattei, T.; Hechard, Y.; Jensen, L.J.; Matondo, M.; et al. A Time-Resolved Multi-Omics Atlas of Acanthamoeba castellanii Encystment. Nat. Commun. 2022, 13, 4104. [Google Scholar] [CrossRef]
- Sharma, M.; Zhang, H.; Ehrenkaufer, G.; Singh, U. Stress Response in Entamoeba Histolytica Is Associated with Robust Processing of tRNA to tRNA Halves. mBio 2023, 14, e03450-22. [Google Scholar] [CrossRef]
- Ilic, D.; Magnussen, H.M.; Tirard, M. Stress—Regulation of SUMO Conjugation and of Other Ubiquitin-Like Modifiers. Semin. Cell Dev. Biol. 2022, 132, 38–50. [Google Scholar] [CrossRef]
- Pandita, T.; Kumar, R.; Horikoshi, N.; Singh, M.; Gupta, A.; Misra, H.; Albuquerque, K.; Hunt, C. Chromatin Modifications and the DNA Damage Response to Ionizing Radiation. Front. Oncol. 2013, 2. [Google Scholar]
- Zhao, Y.; Hu, X.; Liu, Y.; Dong, S.; Wen, Z.; He, W.; Zhang, S.; Huang, Q.; Shi, M. ROS Signaling under Metabolic Stress: Cross-Talk between AMPK and AKT Pathway. Mol. Cancer 2017, 16, 79. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Anand, S.K.; Singh, N.; Dwivedi, U.N.; Kakkar, P. AMP-Activated Protein Kinase: An Energy Sensor and Survival Mechanism in the Reinstatement of Metabolic Homeostasis. Exp. Cell Res. 2023, 428, 113614. [Google Scholar] [CrossRef] [PubMed]
- Di Meo, S.; Reed, T.T.; Venditti, P.; Victor, V.M. Role of ROS and RNS Sources in Physiological and Pathological Conditions. Oxid. Med. Cell. Longev. 2016, 1245049. [Google Scholar] [CrossRef]
- Parzych, K.R.; Klionsky, D.J. An Overview of Autophagy: Morphology, Mechanism, and Regulation. Antioxid. Redox Signal. 2014, 20, 460–473. [Google Scholar] [CrossRef] [PubMed]
- Barna, J.; Csermely, P.; Vellai, T. Roles of Heat Shock Factor 1 beyond the Heat Shock Response. Cell. Mol. Life Sci. 2018, 75, 2897–2916. [Google Scholar] [CrossRef] [PubMed]
- Fulda, S.; Gorman, A.M.; Hori, O.; Samali, A. Cellular Stress Responses: Cell Survival and Cell Death. Int. J. Cell Biol. 2010, 214074. [Google Scholar] [CrossRef]
- Gebert, M.; Sławski, J.; Kalinowski, L.; Collawn, J.F.; Bartoszewski, R. The Unfolded Protein Response: A Double-Edged Sword for Brain Health. Antioxidants 2023, 12, 1648. [Google Scholar] [CrossRef]
- Krishnan, D.; Ghosh, S.K. Morphological and Motility Features of the Stable Bleb-Driven Monopodial Form of Entamoeba and Its Importance in Encystation. Infect. Immun. 2020, 88, e00903-19. [Google Scholar] [CrossRef]
- Chávez-Munguía, B.; Cristóbal-Ramos, A.R.; González-Robles, A.; Tsutsumi, V.; Martínez-Palomo, A. Ultrastructural Study of Entamoeba Invadens Encystation and Excystation. J. Submicrosc. Cytol. Pathol. 2003, 35, 235–243. [Google Scholar]
- Garajová, M.; Mrva, M.; Vaškovicová, N.; Martinka, M.; Melicherová, J.; Valigurová, A. Cellulose Fibrils Formation and Organisation of Cytoskeleton during Encystment Are Essential for Acanthamoeba Cyst Wall Architecture. Sci. Rep. 2019, 9, 4466. [Google Scholar] [CrossRef] [PubMed]
- Chávez-Munguía, B.; Salazar-Villatoro, L.; Lagunes-Guillén, A.; Omaña-Molina, M.; Espinosa-Cantellano, M.; Martínez-Palomo, A. Acanthamoeba Castellanii Cysts: New Ultrastructural Findings. Parasitol. Res. 2013, 112, 1125–1130. [Google Scholar] [CrossRef] [PubMed]
- Herman, E.; Siegesmund, M.A.; Bottery, M.J.; van Aerle, R.; Shather, M.M.; Caler, E.; Dacks, J.B.; van der Giezen, M. Membrane Trafficking Modulation during Entamoeba Encystation. Sci. Rep. 2017, 7, 12854. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.; Naiyer, S.; Bhattacharya, S. Ultra-Structural Analysis and Morphological Changes during the Differentiation of Trophozoite to Cyst in Entamoeba Invadens. Mol. Biochem. Parasitol. 2021, 242, 111363. [Google Scholar] [CrossRef]
- Goswami, P.; Samanta, S.K.; Agarwal, T.; Ghosh, S.K. Stress-Responsive AMP Kinase like Protein Regulates Encystation of Entamoeba Invadens. Mol. Biochem. Parasitol. 2022, 251, 111507. [Google Scholar] [CrossRef]
- Motavallihaghi, S.; Khodadadi, I.; Goudarzi, F.; Afshar, S.; Shahbazi, A.E.; Maghsood, A.H. The Role of Acanthamoeba castellanii (T4 Genotype) Antioxidant Enzymes in Parasite Survival under H2O2-Induced Oxidative Stress. Parasitol. Int. 2022, 87, 102523. [Google Scholar] [CrossRef]
- Sifaoui, I.; -Yanes, E.C.; Reyes-Batlle, M.; Rodríguez-Expósito, R.L.; Bazzocchi, I.L.; Jiménez, I.A.; Piñero, J.E.; Lorenzo-Morales, J.; Weaver, L.K. High Oxygen Concentrations Inhibit Acanthamoeba spp. Parasitol. Res. 2021, 120, 3001–3005. [Google Scholar] [CrossRef]
- Sifaoui, I.; Zeouk, I.; Reyes-Batlle, M.; Lopez-Arencibia, A.; Piñero, J.E.; Lorenzo-Morales, J. Inhibition of Acanthamoeba Polyphaga by Chlorhexidine-Mediated Oxidative Stress Response. J. Glob. Antimicrob. Resist. 2022, 30, 468–473. [Google Scholar] [CrossRef]
- Rodríguez-Expósito, R.L.; Sifaoui, I.; Reyes-Batlle, M.; Maciver, S.K.; Piñero, J.E.; Lorenzo-Morales, J. Statins Induce Actin Cytoskeleton Disassembly and an Apoptosis-Like Process in Acanthamoeba spp. Antibiotics 2022, 11, 280. [Google Scholar] [CrossRef]
- Antos-Krzeminska, N.; Kicinska, A.; Nowak, W.; Jarmuszkiewicz, W. Acanthamoeba Castellanii Uncoupling Protein: A Complete Sequence, Activity, and Role in Response to Oxidative Stress. Int. J. Mol. Sci. 2023, 24, 12501. [Google Scholar] [CrossRef]
- Köhsler, M.; Leitsch, D.; Loufouma Mbouaka, A.; Wekerle, M.; Walochnik, J. Transcriptional Changes of Proteins of the Thioredoxin and Glutathione Systems in Acanthamoeba spp. under Oxidative Stress—An RNA Approach. Parasite 2022, 29, 24. [Google Scholar] [CrossRef] [PubMed]
- de Obeso Fernández del Valle, A.; Scheckhuber, C.Q.; Chavaro-Pérez, D.A.; Ortega-Barragán, E.; Maciver, S.K. mRNA Sequencing Reveals Upregulation of Glutathione S-Transferase Genes during Acanthamoeba Encystation. Microorganisms 2023, 11, 992. [Google Scholar] [CrossRef] [PubMed]
- Motavalli, M.; Khodadadi, I.; Fallah, M.; Maghsood, A.H. Effect of Oxidative Stress on Vital Indicators of Acanthamoeba Castellanii (T4 Genotype). Parasitol. Res. 2018, 117, 2957–2962. [Google Scholar] [CrossRef]
- Jeelani, G.; Nozaki, T. Entamoeba Thiol-Based Redox Metabolism: A Potential Target for Drug Development. Mol. Biochem. Parasitol. 2016, 206, 39–45. [Google Scholar] [CrossRef] [PubMed]
- Husain, A.; Jeelani, G.; Sato, D.; Nozaki, T. Global Analysis of Gene Expression in Response to L-Cysteine Deprivation in the Anaerobic Protozoan Parasite Entamoeba Histolytica. BMC Genom. 2011, 12, 275. [Google Scholar] [CrossRef] [PubMed]
- Santos, H.J.; Nozaki, T. The Mitosome of the Anaerobic Parasitic Protist Entamoeba Histolytica: A Peculiar and Minimalist Mitochondrion-Related Organelle. J. Eukaryot. Microbiol. 2022, 69, e12923. [Google Scholar] [CrossRef] [PubMed]
- Mi-ichi, F.; Miyamoto, T.; Takao, S.; Jeelani, G.; Hashimoto, T.; Hara, H.; Nozaki, T.; Yoshida, H. Entamoeba Mitosomes Play an Important Role in Encystation by Association with Cholesteryl Sulfate Synthesis. Proc. Natl. Acad. Sci. USA 2015, 112, E2884–E2890. [Google Scholar] [CrossRef]
- Ali, V.; Shigeta, Y.; Tokumoto, U.; Takahashi, Y.; Nozaki, T. An Intestinal Parasitic Protist, Entamoeba Histolytica, Possesses a Non-Redundant Nitrogen Fixation-like System for Iron-Sulfur Cluster Assembly under Anaerobic Conditions*. J. Biol. Chem. 2004, 279, 16863–16874. [Google Scholar] [CrossRef]
- Maralikova, B.; Ali, V.; Nakada-Tsukui, K.; Nozaki, T.; Van Der Giezen, M.; Henze, K.; Tovar, J. Bacterial-Type Oxygen Detoxification and Iron-Sulfur Cluster Assembly in Amoebal Relict Mitochondria. Cell. Microbiol. 2010, 12, 331–342. [Google Scholar] [CrossRef]
- van der Giezen, M.; Cox, S.; Tovar, J. The Iron-Sulfur Cluster Assembly Genes iscS and iscU of Entamoeba Histolytica Were Acquired by Horizontal Gene Transfer. BMC Evol. Biol. 2004, 4, 7. [Google Scholar] [CrossRef]
- Leitsch, D.; Williams, C.F.; Hrdý, I. Redox Pathways as Drug Targets in Microaerophilic Parasites. Trends Parasitol. 2018, 34, 576–589. [Google Scholar] [CrossRef] [PubMed]
- Husain, A.; Sato, D.; Jeelani, G.; Soga, T.; Nozaki, T. Dramatic Increase in Glycerol Biosynthesis upon Oxidative Stress in the Anaerobic Protozoan Parasite Entamoeba Histolytica. PLoS Negl. Trop. Dis. 2012, 6, e1831. [Google Scholar] [CrossRef] [PubMed]
- Jeelani, G.; Sato, D.; Husain, A.; Escueta-de Cadiz, A.; Sugimoto, M.; Soga, T.; Suematsu, M.; Nozaki, T. Metabolic Profiling of the Protozoan Parasite Entamoeba Invadens Revealed Activation of Unpredicted Pathway during Encystation. PLoS ONE 2012, 7, e37740. [Google Scholar] [CrossRef] [PubMed]
- Santos, H.J.; Hanadate, Y.; Imai, K.; Watanabe, H.; Nozaki, T. Entamoeba Histolytica EHD1 Is Involved in Mitosome-Endosome Contact. mBio 2022, 13, e03849-21. [Google Scholar] [CrossRef]
- Pineda, E.; Perdomo, D. Entamoeba Histolytica under Oxidative Stress: What Countermeasure Mechanisms Are in Place? Cells 2017, 6, 44. [Google Scholar] [CrossRef]
- Kumari, R.; Gupta, P.; Tiwari, S. Ubc7/Ube2g2 Ortholog in Entamoeba Histolytica: Connection with the Plasma Membrane and Phagocytosis. Parasitol. Res. 2018, 117, 1599–1611. [Google Scholar] [CrossRef]
- Hendrick, H.M.; Welter, B.H.; Hapstack, M.A.; Sykes, S.E.; Sullivan, W.J.; Temesvari, L.A. Phosphorylation of Eukaryotic Initiation Factor-2α during Stress and Encystation in Entamoeba Species. PLoS Pathog. 2016, 12, e1006085. [Google Scholar] [CrossRef]
- Walters, H.A.; Welter, B.H.; Sullivan, W.J.; Temesvari, L.A. Phosphorylation of Eukaryotic Initiation Factor 2-α in Response to Endoplasmic Reticulum and Nitrosative Stress in the Human Protozoan Parasite, Entamoeba Histolytica. Mol. Biochem. Parasitol. 2019, 234, 111223. [Google Scholar] [CrossRef]
- Walters, H.A.; Welter, B.H.; Moss, H.C.; Villano, M.A.; Orobio-Hurtado, R.; Sullivan, W.J.; Temesvari, L.A. Eukaryotic Initiation Factor 2α Kinases Regulate Virulence Functions, Stage Conversion, and the Stress Response in Entamoeba Invadens. mSphere 2022, 7, e00131-22. [Google Scholar] [CrossRef]
- Santos, F.; Nequiz, M.; Hernández-Cuevas, N.A.; Hernández, K.; Pineda, E.; Encalada, R.; Guillén, N.; Luis-García, E.; Saralegui, A.; Saavedra, E.; et al. Maintenance of Intracellular Hypoxia and Adequate Heat Shock Response Are Essential Requirements for Pathogenicity and Virulence of Entamoeba Histolytica. Cell. Microbiol. 2015, 17, 1037–1051. [Google Scholar] [CrossRef]
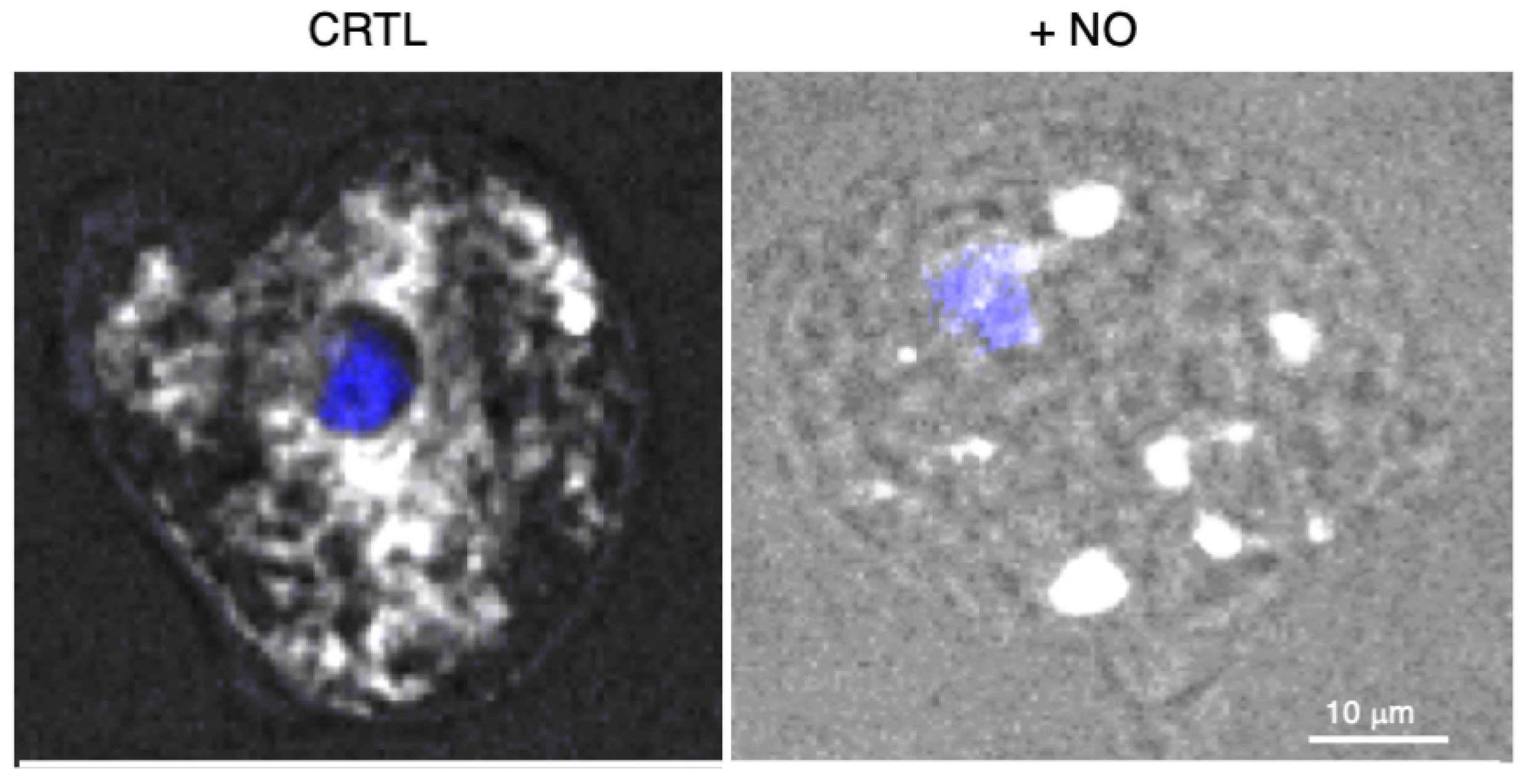
- Santi-Rocca, J.; Smith, S.; Weber, C.; Pineda, E.; Hon, C.-C.; Saavedra, E.; Olivos-García, A.; Rousseau, S.; Dillies, M.-A.; Coppée, J.-Y.; et al. Endoplasmic Reticulum Stress-Sensing Mechanism Is Activated in Entamoeba Histolytica upon Treatment with Nitric Oxide. PLoS ONE 2012, 7, e31777. [Google Scholar] [CrossRef] [PubMed]
- Vilas-Boas, E.A.; Almeida, D.C.; Roma, L.P.; Ortis, F.; Carpinelli, A.R. Lipotoxicity and β-Cell Failure in Type 2 Diabetes: Oxidative Stress Linked to NADPH Oxidase and ER Stress. Cells 2021, 10, 3328. [Google Scholar] [CrossRef] [PubMed]
- Bhattarai, K.R.; Riaz, T.A.; Kim, H.-R.; Chae, H.-J. The Aftermath of the Interplay between the Endoplasmic Reticulum Stress Response and Redox Signaling. Exp. Mol. Med. 2021, 53, 151–167. [Google Scholar] [CrossRef] [PubMed]
- Pearez-Serranoa, J.; MartõÂneza, J.; PeÂreza, B.; Bernadinab, W.E.; RodrõÂguez-Caabeiroa, F. In Vitro Shock Response to Different Stressors in Free Living and Pathogenic Acanthamoeba. Int. J. Parasitol. 2000, 30, 829–835. [Google Scholar] [CrossRef] [PubMed]
- Wideman, J.G.; Gawryluk, R.M.R.; Gray, M.W.; Dacks, J.B. The Ancient and Widespread Nature of the ER–Mitochondria Encounter Structure. Mol. Biol. Evol. 2013, 30, 2044–2049. [Google Scholar] [CrossRef]
- Lefaki, M.; Papaevgeniou, N.; Chondrogianni, N. Redox Regulation of Proteasome Function. Redox Biol. 2017, 13, 452–458. [Google Scholar] [CrossRef]
- Siddiqui, R.; Saleem, S.; Khan, N.A. The Effect of Peptidic and Non-Peptidic Proteasome Inhibitors on the Biological Properties of Acanthamoeba Castellanii Belonging to the T4 Genotype. Exp. Parasitol. 2016, 168, 16–24. [Google Scholar] [CrossRef]
- Grau-Bové, X.; Sebé-Pedrós, A.; Ruiz-Trillo, I. The Eukaryotic Ancestor Had a Complex Ubiquitin Signaling System of Archaeal Origin. Mol. Biol. Evol. 2015, 32, 726–739. [Google Scholar] [CrossRef]
- Arya, S.; Sharma, G.; Gupta, P.; Tiwari, S. In Silico Analysis of Ubiquitin/Ubiquitin-like Modifiers and Their Conjugating Enzymes in Entamoeba Species. Parasitol. Res. 2012, 111, 37–51. [Google Scholar] [CrossRef]
- Moon, E.-K.; Chung, D.-I.; Hong, Y.-C.; Ahn, T.-I.; Kong, H.-H. Acanthamoeba Castellanii: Gene Profile of Encystation by ESTs Analysis and KOG Assignment. Exp. Parasitol. 2008, 119, 111–116. [Google Scholar] [CrossRef]
- Woo, J.S.; Liang, S.T.; Lo, R.L. Significance of an Absent or Reversed End Diastolic Flow in Doppler Umbilical Artery Waveforms. J. Ultrasound Med. 1987, 6, 291–297. [Google Scholar] [CrossRef]
- Sou, Y.; Tanida, I.; Komatsu, M.; Ueno, T.; Kominami, E. Phosphatidylserine in Addition to Phosphatidylethanolamine Is an in Vitro Target of the Mammalian Atg8 Modifiers, LC3, GABARAP, and GATE-16. J. Biol. Chem. 2006, 281, 3017–3024. [Google Scholar] [CrossRef] [PubMed]
- Régnacq, M.; Voisin, P.; Héchard, Y.; Bergès, T.; Braquart-Varnier, C.; Samba-Louaka, A. Identification of Atg8 from Acanthamoeba Castellanii by Genetic Complementation in Saccharomyces Cerevisiae. Mol. Biochem. Parasitol. 2016, 210, 55–57. [Google Scholar] [CrossRef] [PubMed]
- Moon, E.-K.; Chung, D.-I.; Hong, Y.; Kong, H.-H. Atg3-Mediated Lipidation of Atg8 Is Involved in Encystation of Acanthamoeba. Korean J. Parasitol. 2011, 49, 103–108. [Google Scholar] [CrossRef] [PubMed]
- Nakada-Tsukui, K.; Watanabe, N.; Shibata, K.; Wahyuni, R.; Miyamoto, E.; Nozaki, T. Proteomic Analysis of Atg8-Dependent Recruitment of Phagosomal Proteins in the Enteric Protozoan Parasite Entamoeba Histolytica. Front. Cell. Infect. Microbiol. 2022, 12, 961645. [Google Scholar] [CrossRef]
- Picazarri, K.; Nakada-Tsukui, K.; Tsuboi, K.; Miyamoto, E.; Watanabe, N.; Kawakami, E.; Nozaki, T. Atg8 Is Involved in Endosomal and Phagosomal Acidification in the Parasitic Protist E Ntamoeba Histolytica. Cell. Microbiol. 2015, 17, 1510–1522. [Google Scholar] [CrossRef]
- Duszenko, M.; Ginger, M.L.; Brennand, A.; Gualdrón-López, M.; Colombo, M.I.; Coombs, G.H.; Coppens, I.; Jayabalasingham, B.; Langsley, G.; Lisboa de Castro, S.; et al. Autophagy in Protists. Autophagy 2011, 7, 127–158. [Google Scholar] [CrossRef]
- Kim, S.-H.; Moon, E.-K.; Hong, Y.; Chung, D.-I.; Kong, H.-H. Autophagy Protein 12 Plays an Essential Role in Acanthamoeba Encystation. Exp. Parasitol. 2015, 159, 46–52. [Google Scholar] [CrossRef]
- Mizushima, N.; Noda, T.; Ohsumi, Y. Apg16p Is Required for the Function of the Apg12p–Apg5p Conjugate in the Yeast Autophagy Pathway. EMBO J. 1999, 18, 3888–3896. [Google Scholar] [CrossRef]
- Song, S.-M.; Han, B.-I.; Moon, E.-K.; Lee, Y.-R.; Yu, H.S.; Jha, B.K.; Danne, D.-B.S.; Kong, H.-H.; Chung, D.-I.; Hong, Y. Autophagy Protein 16-Mediated Autophagy Is Required for the Encystation of Acanthamoeba Castellanii. Mol. Biochem. Parasitol. 2012, 183, 158–165. [Google Scholar] [CrossRef]
- Komatsu, M.; Chiba, T.; Tatsumi, K.; Iemura, S.; Tanida, I.; Okazaki, N.; Ueno, T.; Kominami, E.; Natsume, T.; Tanaka, K. A Novel Protein-Conjugating System for Ufm1, a Ubiquitin-Fold Modifier. EMBO J. 2004, 23, 1977–1986. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Ma, X.; Jing, Y.; Zhang, G.; Zhang, D.; Mao, Z.; Ma, X.; Liu, H.; Chen, F. P4HB UFMylation Regulates Mitochondrial Function and Oxidative Stress. Free Radic Biol. Med. 2022, 188, 277–286. [Google Scholar] [CrossRef] [PubMed]
- Scavone, F.; Gumbin, S.C.; Da Rosa, P.A.; Kopito, R.R. RPL26/uL24 UFMylation Is Essential for Ribosome-Associated Quality Control at the Endoplasmic Reticulum. Proc. Natl. Acad. Sci. USA 2023, 120, e2220340120. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Q.; Wang, Y.; Xiang, M.; Hua, J.; Zhou, T.; Chen, F.; Lv, X.; Huang, J.; Cai, Y. UFL1, a UFMylation E3 Ligase, Plays a Crucial Role in Multiple Cellular Stress Responses. Front. Endocrinol. 2023, 14, 1123124. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.; Wang, X.; Li, D.; Cao, L.; Cui, H.; Xu, G. UFBP1, a Key Component in Ufmylation, Enhances Drug Sensitivity by Promoting Proteasomal Degradation of Oxidative Stress-Response Transcription Factor Nrf2. Oncogene 2021, 40, 647–662. [Google Scholar] [CrossRef]
- Karpiyevich, M.; Artavanis-Tsakonas, K. Ubiquitin-Like Modifiers: Emerging Regulators of Protozoan Parasites. Biomolecules 2020, 10, 1403. [Google Scholar] [CrossRef]
- Gannavaram, S.; Sharma, P.; Duncan, R.C.; Salotra, P.; Nakhasi, H.L. Mitochondrial Associated Ubiquitin Fold Modifier-1 Mediated Protein Conjugation in Leishmania Donovani. PLoS ONE 2011, 6, e16156. [Google Scholar] [CrossRef]
- Tan, Q.; Wang, J.; Chen, J.; Liu, X.; Chen, X.; Xiao, Q.; Li, J.; Li, H.; Zhao, X.; Zhang, X. Involvement of Urm1, a Ubiquitin-Like Protein, in the Regulation of Oxidative Stress Response of Toxoplasma Gondii. Microbiol. Spectr. 2022, 10, e02394-21. [Google Scholar] [CrossRef]
- Ravichandran, K.E.; Kaduhr, L.; Skupien-Rabian, B.; Shvetsova, E.; Sokołowski, M.; Krutyhołowa, R.; Kwasna, D.; Brachmann, C.; Lin, S.; Guzman Perez, S.; et al. E2/E3-independent Ubiquitin-like Protein Conjugation by Urm1 Is Directly Coupled to Cysteine Persulfidation. EMBO J. 2022, 41, e111318. [Google Scholar] [CrossRef]
- Khoshnood, B.; Dacklin, I.; Grabbe, C. A Proteomics Approach to Identify Targets of the Ubiquitin-like Molecule Urm1 in Drosophila Melanogaster. PLoS ONE 2017, 12, e0185611. [Google Scholar] [CrossRef]
- Brachmann, C.; Kaduhr, L.; Jüdes, A.; Ravichandran, K.E.; West, J.D.; Glatt, S.; Schaffrath, R. Redox Requirements for Ubiquitin-like Urmylation of Ahp1, a 2-Cys Peroxiredoxin from Yeast. Redox Biol. 2020, 30, 101438. [Google Scholar] [CrossRef] [PubMed]
- Verner, Z.; Žárský, V.; Le, T.; Narayanasamy, R.K.; Rada, P.; Rozbeský, D.; Makki, A.; Belišová, D.; Hrdý, I.; Vancová, M.; et al. Anaerobic Peroxisomes in Entamoeba Histolytica Metabolize Myo-Inositol. PLoS Pathog. 2021, 17, e1010041. [Google Scholar] [CrossRef] [PubMed]
- Shabardina, V.; Kischka, T.; Kmita, H.; Suzuki, Y.; Makałowski, W. Environmental Adaptation of Acanthamoeba Castellanii and Entamoeba Histolytica at Genome Level as Seen by Comparative Genomic Analysis. Int. J. Biol. Sci. 2018, 14, 306–320. [Google Scholar] [CrossRef] [PubMed]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Samba-Louaka, A.; Labruyère, E.; Matondo, M.; Locard-Paulet, M.; Olivo-Marin, J.-C.; Guillen, N. Encystation and Stress Responses under the Control of Ubiquitin-like Proteins in Pathogenic Amoebae. Microorganisms 2023, 11, 2670. https://doi.org/10.3390/microorganisms11112670
Samba-Louaka A, Labruyère E, Matondo M, Locard-Paulet M, Olivo-Marin J-C, Guillen N. Encystation and Stress Responses under the Control of Ubiquitin-like Proteins in Pathogenic Amoebae. Microorganisms. 2023; 11(11):2670. https://doi.org/10.3390/microorganisms11112670
Chicago/Turabian StyleSamba-Louaka, Ascel, Elisabeth Labruyère, Mariette Matondo, Marie Locard-Paulet, Jean-Christophe Olivo-Marin, and Nancy Guillen. 2023. "Encystation and Stress Responses under the Control of Ubiquitin-like Proteins in Pathogenic Amoebae" Microorganisms 11, no. 11: 2670. https://doi.org/10.3390/microorganisms11112670