

The Effects of Lactobacillus johnsonii on Diseases and Its Potential Applications
 and
and
Abstract
:1. Introduction
2. Comprehensive Characteristics of Identified L. johnsonii

{kind=link}
{kind=link}
{kind=link}
| Strain | BioSample | Size (Kb) | Replicons | CDS | Release Date |
|---|---|---|---|---|---|
| FI9785 | SAMEA2272487 | 3.55 | p9785S:NC_012552.1/AY862141.1 [19] | 2 | April 2009 |
| FI9785 | SAMEA2272487 | 26.27 | p9785L:NC_013505.1/FN357112.1 [20] | 26 | November 2009 |
| BS15 | SAMN04631277 | 45.84 | LJBSp1:NZ_CP016630.1/CP016630.1 | 43 | August 2016 |
| UMNLJ22 | SAMN04573146 | 27.88 | pUMNLJ22_1:NZ_CP021705.1/CP021705.1 | 34 | June 2017 |
| UMNLJ22 | SAMN04573146 | 24.93 | pUMNLJ22_2:NZ_CP021706.1/CP021706.1 | 24 | June 2017 |
| UMNLJ21 | SAMN04573145 | 21.52 | pUMNLJ21_1:NZ_CP021701.1/CP021701.1 | 22 | June 2017 |
| UMNLJ21 | SAMN04573145 | 15.25 | pUMNLJ21_2:NZ_CP021702.1/CP021702.1 | 20 | June 2017 |
| pf01 | SAMN02469597 | 26.46 | pLJPF01L:CP024782.1 [21] | 0 | November 2017 |
| pf01 | SAMN02469597 | 14.24 | pLJPF01S:CP024783.1 [21] | 0 | November 2017 |
| LL8 | SAMN13266521 | 77.56 | unnamed:NZ_CM019125.1/CM019125.1 | 73 | December 2019 |
| DC22.2 | SAMN11371966 | 7.65 | pLjDC22.2_1:NZ_CP039262.1/CP039262.1 | 10 | January 2020 |
| DC22.2 | SAMN11371966 | 5.75 | pLjDC22.2_2:NZ_CP039263.1/CP039263.1 | 3 | January 2020 |
| DC22.2 | SAMN11371966 | 7.08 | pLjDC22.1_3:NZ_CP039264.1/CP039264.1 | 4 | January 2020 |
| DC22.2 | SAMN11371966 | 13.77 | pLjDC22.2_4:NZ_CP039265.1/CP039265.1 | 6 | January 2020 |
| G2A | SAMN11618738 | 130.11 | unnamed1:NZ_CP040855.1/CP040855.1 | 154 | March 2020 |
| G2A | SAMN11618738 | 108.72 | unnamed2:NZ_CP040856.1/CP040856.1 | 103 | March 2020 |
| GHZ10a | SAMN16131614 | 13.65 | unnamed1:NZ_CP062069.1/CP062069.1 | 15 | October 2020 |
| GHZ10a | SAMN16131614 | 15.79 | unnamed2:NZ_CP062070.1/CP062070.1 | 18 | October 2020 |
3. Effects of L. johnsonii on Different Diseases
4. The Common Mechanism of L. johnsonii in Different Diseases
4.1. Respiratory Insults
4.2. Gastrointestinal Disease
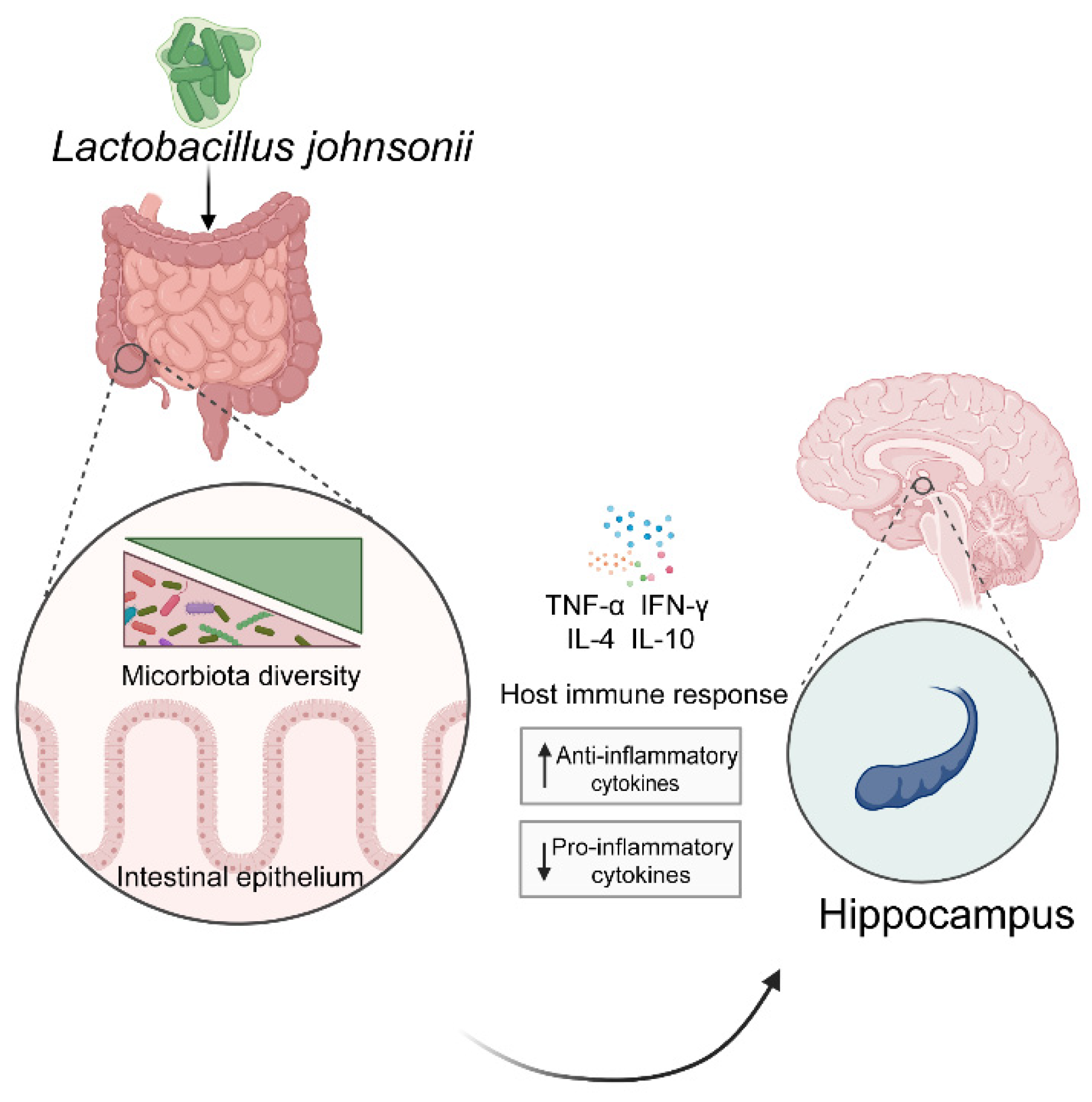
4.3. Mental Health
4.4. Obesity
4.5. Liver Diseases
5. Conclusions and Perspectives
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. Expert Consensus Document. The international scientific association for probiotics and prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.-Y.; Adachi, Y. Biological and genetic classification of canine intestinal lactic acid bacteria and bifidobacteria. Microbiol. Immunol. 2007, 51, 919–928. [Google Scholar] [CrossRef] [PubMed]
- Carina Audisio, M.; Torres, M.J.; Sabaté, D.C.; Ibarguren, C.; Apella, M.C. Properties of different lactic acid bacteria isolated from Apis mellifera L. Bee-gut. Microbiol. Res. 2011, 166, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Duar, R.M.; Lin, X.B.; Zheng, J.; Martino, M.E.; Grenier, T.; Pérez-Muñoz, M.E.; Leulier, F.; Gänzle, M.; Walter, J. Lifestyles in Transition: Evolution and Natural History of the Genus Lactobacillus. FEMS Microbiol. Rev. 2017, 41, S27–S48. [Google Scholar] [CrossRef] [PubMed]
- Yan, W.; Sun, C.; Yuan, J.; Yang, N. Gut metagenomic analysis reveals prominent roles of Lactobacillus and cecal microbiota in chicken feed efficiency. Sci. Rep. 2017, 7, 45308. [Google Scholar] [CrossRef] [PubMed]
- Rhee, S.J.; Lee, J.-E.; Lee, C.-H. Importance of lactic acid bacteria in Asian fermented foods. Microb. Cell Fact. 2011, 10 (Suppl. S1), S5. [Google Scholar] [CrossRef]
- Andrada, E.; Mechoud, M.A.; Abeijón-Mukdsi, M.C.; Chagra Dib, E.P.; Cerviño, S.; Perez Chaia, A.; Medina, R.B. Ferulic acid esterase producing Lactobacillus johnsonii from goat feces as corn silage inoculants. Microorganisms 2022, 10, 1732. [Google Scholar] [CrossRef]
- Mager, L.F.; Burkhard, R.; Pett, N.; Cooke, N.C.A.; Brown, K.; Ramay, H.; Paik, S.; Stagg, J.; Groves, R.A.; Gallo, M.; et al. Microbiome-derived inosine modulates response to checkpoint inhibitor immunotherapy. Science 2020, 369, 1481–1489. [Google Scholar] [CrossRef]
- O’Callaghan, J.; O’Toole, P.W. Lactobacillus: Host-microbe relationships. Curr. Top. Microbiol. Immunol. 2013, 358, 119–154. [Google Scholar] [CrossRef]
- Buhnik-Rosenblau, K.; Matsko-Efimov, V.; Jung, M.; Shin, H.; Danin-Poleg, Y.; Kashi, Y. Indication for co-evolution of Lactobacillus johnsonii with its hosts. BMC Microbiol. 2012, 12, 149. [Google Scholar] [CrossRef]
- Fan, X.; Mai, C.; Zuo, L.; Huang, J.; Xie, C.; Jiang, Z.; Li, R.; Yao, X.; Fan, X.; Wu, Q.; et al. Herbal formula BaWeiBaiDuSan alleviates polymicrobial sepsis-induced liver injury via increasing the gut microbiota Lactobacillus johnsonii and regulating macrophage anti-inflammatory activity in mice. Acta Pharm. Sin. B 2023, 13, 1164–1179. [Google Scholar] [CrossRef]
- Fonseca, W.; Malinczak, C.-A.; Fujimura, K.; Li, D.; McCauley, K.; Li, J.; Best, S.K.K.; Zhu, D.; Rasky, A.J.; Johnson, C.C.; et al. Maternal gut microbiome regulates immunity to RSV infection in offspring. J. Exp. Med. 2021, 218, e20210235. [Google Scholar] [CrossRef]
- Mu, Q.; Zhang, H.; Liao, X.; Lin, K.; Liu, H.; Edwards, M.R.; Ahmed, S.A.; Yuan, R.; Li, L.; Cecere, T.E.; et al. Control of lupus nephritis by changes of gut microbiota. Microbiome 2017, 5, 73. [Google Scholar] [CrossRef] [PubMed]
- Fonseca, W.; Lucey, K.; Jang, S.; Fujimura, K.E.; Rasky, A.; Ting, H.-A.; Petersen, J.; Johnson, C.C.; Boushey, H.A.; Zoratti, E.; et al. Lactobacillus johnsonii supplementation attenuates respiratory viral infection via metabolic reprogramming and immune cell modulation. Mucosal Immunol. 2017, 10, 1569–1580. [Google Scholar] [CrossRef] [PubMed]
- Sharma, P.; Tomar, S.K.; Sangwan, V.; Goswami, P.; Singh, R. Antibiotic resistance of Lactobacillus sp. isolated from commercial probiotic preparations. J. Food Saf. 2016, 36, 38–51. [Google Scholar] [CrossRef]
- Guo, H.; Pan, L.; Li, L.; Lu, J.; Kwok, L.; Menghe, B.; Zhang, H.; Zhang, W. Characterization of antibiotic resistance genes from Lactobacillus isolated from traditional dairy products. J. Food Sci. 2017, 82, 724–730. [Google Scholar] [CrossRef] [PubMed]
- Dec, M.; Nowaczek, A.; Stępień-Pyśniak, D.; Wawrzykowski, J.; Urban-Chmiel, R. Identification and antibiotic susceptibility of Lactobacilli isolated from turkeys. BMC Microbiol. 2018, 18, 168. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q.; Pei, Z.; Fang, Z.; Wang, H.; Zhu, J.; Lee, Y.; Zhang, H.; Zhao, J.; Lu, W.; Chen, W. Evaluation of tetracycline resistance and determination of the tentative microbiological cutoff values in lactic acid bacterial species. Microorganisms 2021, 9, 2128. [Google Scholar] [CrossRef]
- Horn, N.; Wegmann, U.; Narbad, A.; Gasson, M.J. Characterisation of a novel plasmid P9785S from Lactobacillus johnsonii FI9785. Plasmid 2005, 54, 176–183. [Google Scholar] [CrossRef]
- Wegmann, U.; Overweg, K.; Horn, N.; Goesmann, A.; Narbad, A.; Gasson, M.J.; Shearman, C. Complete genome sequence of Lactobacillus johnsonii FI9785, a competitive exclusion agent against pathogens in poultry. J. Bacteriol. 2009, 191, 7142–7143. [Google Scholar] [CrossRef]
- Lee, J.H.; Chae, J.P.; Lee, J.Y.; Lim, J.-S.; Kim, G.-B.; Ham, J.-S.; Chun, J.; Kang, D.-K. Genome sequence of Lactobacillus johnsonii PF01, isolated from piglet feces. J. Bacteriol. 2011, 193, 5030–5031. [Google Scholar] [CrossRef] [PubMed]
- Lyu, M.; Bai, Y.; Orihara, K.; Miyanaga, K.; Yamamoto, N. GAPDH released from Lactobacillus johnsonii MG enhances barrier function by upregulating genes associated with tight junctions. Microorganisms 2023, 11, 1393. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Li, Y.; Chu, B.; Yuan, L.; Liu, N.; Zhu, Y.; Wang, J. Lactobacillus johnsonii L531 alleviates the damage caused by salmonella typhimurium via inhibiting TLR4, NF-ΚB, and NLRP3 inflammasome signaling pathways. Microorganisms 2021, 9, 1983. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Xia, B.; He, T.; Li, D.; Su, J.-H.; Guo, L.; Wang, J.; Zhu, Y.-H. Oral administration of a select mixture of Lactobacillus and Bacillus alleviates inflammation and maintains mucosal barrier integrity in the ileum of pigs challenged with salmonella infantis. Microorganisms 2019, 7, 135. [Google Scholar] [CrossRef]
- Frei, R.; Akdis, M.; O’Mahony, L. Prebiotics, probiotics, synbiotics, and the immune system: Experimental data and clinical evidence. Curr. Opin. Gastroenterol. 2015, 31, 153–158. [Google Scholar] [CrossRef]
- van Baarlen, P.; Wells, J.M.; Kleerebezem, M. Regulation of intestinal homeostasis and immunity with probiotic lactobacilli. Trends Immunol. 2013, 34, 208–215. [Google Scholar] [CrossRef]
- Salazar, N.; Ruas-Madiedo, P.; Kolida, S.; Collins, M.; Rastall, R.; Gibson, G.; de Los Reyes-Gavilán, C.G. Exopolysaccharides produced by Bifidobacterium longum IPLA E44 and Bifidobacterium animalis subsp. Lactis IPLA R1 modify the composition and metabolic activity of human faecal microbiota in PH-controlled batch cultures. Int. J. Food Microbiol. 2009, 135, 260–267. [Google Scholar] [CrossRef]
- Charlet, R.; Le Danvic, C.; Sendid, B.; Nagnan-Le Meillour, P.; Jawhara, S. Oleic acid and palmitic acid from Bacteroides thetaiotaomicron and Lactobacillus johnsonii exhibit anti-inflammatory and antifungal properties. Microorganisms 2022, 10, 1803. [Google Scholar] [CrossRef]
- Ki Cha, B.; Mun Jung, S.; Hwan Choi, C.; Song, I.-D.; Woong Lee, H.; Joon Kim, H.; Hyuk, J.; Kyung Chang, S.; Kim, K.; Chung, W.-S.; et al. The effect of a multispecies probiotic mixture on the symptoms and fecal icrobiota in diarrhea-dominant irritable bowel syndrome: A randomized, double-blind, placebo-controlled trial. J. Clin. Gastroenterol. 2012, 46, 220–227. [Google Scholar] [CrossRef]
- Sanders, M.E.; Benson, A.; Lebeer, S.; Merenstein, D.J.; Klaenhammer, T.R. Shared mechanisms among probiotic taxa: Implications for general probiotic claims. Curr. Opin. Biotechnol. 2018, 49, 207–216. [Google Scholar] [CrossRef]
- Sanders, M.E.; Merenstein, D.J.; Reid, G.; Gibson, G.R.; Rastall, R.A. Probiotics and prebiotics in intestinal health and disease: From biology to the clinic. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 605–616. [Google Scholar] [CrossRef] [PubMed]
- Inoue, R.; Otsuka, M.; Nishio, A.; Ushida, K. Primary administration of Lactobacillus johnsonii NCC533 in weaning period suppresses the elevation of proinflammatory cytokines and CD86 gene expressions in skin lesions in NC/Nga mice. FEMS Immunol. Med. Microbiol. 2007, 50, 67–76. [Google Scholar] [CrossRef] [PubMed]
- Valladares, R.; Sankar, D.; Li, N.; Williams, E.; Lai, K.-K.; Abdelgeliel, A.S.; Gonzalez, C.F.; Wasserfall, C.H.; Larkin, J.; Schatz, D.; et al. Actobacillus johnsonii N6.2 mitigates the development of type 1 diabetes in BB-DP rats. PLoS ONE 2010, 5, e10507. [Google Scholar] [CrossRef] [PubMed]
- Lau, K.; Benitez, P.; Ardissone, A.; Wilson, T.D.; Collins, E.L.; Lorca, G.; Li, N.; Sankar, D.; Wasserfall, C.; Neu, J.; et al. Inhibition of type 1 diabetes correlated to a Lactobacillus johnsonii N6.2-mediated Th17 bias. J. Immunol. 2011, 186, 3538–3546. [Google Scholar] [CrossRef]
- Teixeira, L.D.; Kling, D.N.; Lorca, G.L. Lactobacillus johnsonii N6.2 diminishes caspase-1 maturation in the gastrointestinal system of diabetes prone rats. Benef. Microbes 2018, 9, 527–539. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.-H.; Yen, H.-R.; Lu, W.-L.; Ho, H.-H.; Lin, W.-Y.; Kuo, Y.-W.; Huang, Y.-Y.; Tsai, S.-Y.; Lin, H.-C. Adjuvant probiotics of Lactobacillus salivarius subsp. Salicinius AP-32, L. Johnsonii MH-68, and Bifidobacterium animalis subsp. Lactis CP-9 attenuate glycemic levels and inflammatory cytokines in patients with type 1 diabetes mellitus. Front. Endocrinol. 2022, 13, 754401. [Google Scholar] [CrossRef]
- Hsieh, P.-S.; Ho, H.-H.; Tsao, S.P.; Hsieh, S.-H.; Lin, W.-Y.; Chen, J.-F.; Kuo, Y.-W.; Tsai, S.-Y.; Huang, H.-Y. Multi-strain probiotic supplement attenuates streptozotocin-induced type-2 diabetes by reducing inflammation and β-cell death in rats. PLoS ONE 2021, 16, e0251646. [Google Scholar] [CrossRef]
- Fujimura, K.E.; Demoor, T.; Rauch, M.; Faruqi, A.A.; Jang, S.; Johnson, C.C.; Boushey, H.A.; Zoratti, E.; Ownby, D.; Lukacs, N.W.; et al. House dust exposure mediates gut microbiome Lactobacillus enrichment and airway immune defense against allergens and virus infection. Proc. Natl. Acad. Sci. USA 2014, 111, 805–810. [Google Scholar] [CrossRef]
- Felley, C.P.; Corthésy-Theulaz, I.; Rivero, J.L.; Sipponen, P.; Kaufmann, M.; Bauerfeind, P.; Wiesel, P.H.; Brassart, D.; Pfeifer, A.; Blum, A.L.; et al. Favourable effect of an acidified milk (LC-1) on Helicobacter pylori gastritis in man. Eur. J. Gastroenterol. Hepatol. 2001, 13, 25–29. [Google Scholar] [CrossRef]
- Gotteland, M.; Cruchet, S. Suppressive effect of frequent ingestion of Lactobacillus johnsonii La1 on Helicobacter pylori colonization in asymptomatic volunteers. J. Antimicrob. Chemother. 2003, 51, 1317–1319. [Google Scholar] [CrossRef]
- Michetti, P.; Dorta, G.; Wiesel, P.H.; Brassart, D.; Verdu, E.; Herranz, M.; Felley, C.; Porta, N.; Rouvet, M.; Blum, A.L.; et al. Effect of whey-based culture supernatant of Lactobacillus acidophilus (johnsonii) La1 on Helicobacter pylori infection in humans. Digestion 1999, 60, 203–209. [Google Scholar] [CrossRef] [PubMed]
- Cruchet, S.; Obregon, M.C.; Salazar, G.; Diaz, E.; Gotteland, M. Effect of the ingestion of a dietary product containing Lactobacillus johnsonii La1 on Helicobacter pylori colonization in children. Nutrition 2003, 19, 716–721. [Google Scholar] [CrossRef] [PubMed]
- Sgouras, D.N.; Panayotopoulou, E.G.; Martinez-Gonzalez, B.; Petraki, K.; Michopoulos, S.; Mentis, A. Lactobacillus johnsonii La1 attenuates Helicobacter Pylori-associated gastritis and reduces levels of proinflammatory chemokines in C57BL/6 mice. Clin. Diagn. Lab. Immunol. 2005, 12, 1378–1386. [Google Scholar] [CrossRef] [PubMed]
- Aiba, Y.; Nakano, Y.; Koga, Y.; Takahashi, K.; Komatsu, Y. A highly acid-resistant novel strain of Lactobacillus johnsonii No. 1088 has antibacterial activity, including that against Helicobacter Pylori, and inhibits gastrin-mediated acid production in mice. Microbiologyopen 2015, 4, 465–474. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, P.-S.; Tsai, Y.-C.; Chen, Y.-C.; Teh, S.-F.; Ou, C.-M.; King, V.A.-E. Eradication of Helicobacter pylori infection by the probiotic strains Lactobacillus johnsonii MH-68 and L. Salivarius ssp. Salicinius AP-32. Helicobacter 2012, 17, 466–477. [Google Scholar] [CrossRef]
- Pantoflickova, D.; Corthésy-Theulaz, I.; Dorta, G.; Stolte, M.; Isler, P.; Rochat, F.; Enslen, M.; Blum, A.L. Favourable effect of regular intake of fermented milk containing Lactobacillus johnsonii on Helicobacter pylori associated gastritis. Aliment. Pharmacol. Ther. 2003, 18, 805–813. [Google Scholar] [CrossRef]
- Hu, Y.; Zhao, M.; Lu, Z.; Lv, F.; Zhao, H.; Bie, X.L. johnsonii, L. plantarum, and L. rhamnosus alleviated enterohaemorrhagic Escherichia coli-induced diarrhoea in mice by regulating gut microbiota. Microb. Pathog. 2021, 154, 104856. [Google Scholar] [CrossRef]
- Wang, Y.; Li, A.; Liu, J.; Mehmood, K.; Wangdui, B.; Shi, H.; Luo, X.; Zhang, H.; Li, J.L. Pseudomesenteroides and L. johnsonii isolated from yaks in Tibet modulate gut microbiota in mice to ameliorate enteroinvasive Escherichia coli-induced diarrhea. Microb. Pathog. 2019, 132, 1–9. [Google Scholar] [CrossRef]
- Jia, D.-J.-C.; Wang, Q.-W.; Hu, Y.-Y.; He, J.-M.; Ge, Q.-W.; Qi, Y.-D.; Chen, L.-Y.; Zhang, Y.; Fan, L.-N.; Lin, Y.-F.; et al. Lactobacillus johnsonii alleviates colitis by TLR1/2-STAT3 mediated CD206+ macrophagesIL-10 activation. Gut Microbes 2022, 14, 2145843. [Google Scholar] [CrossRef]
- Fukushima, Y.; Miyaguchi, S.; Yamano, T.; Kaburagi, T.; Iino, H.; Ushida, K.; Sato, K. Improvement of nutritional status and incidence of infection in hospitalised, enterally fed elderly by feeding of fermented milk containing probiotic Lactobacillus johnsonii La1 (NCC533). Br. J. Nutr. 2007, 98, 969–977. [Google Scholar] [CrossRef]
- Zhang, Y.; Mu, T.; Yang, Y.; Zhang, J.; Ren, F.; Wu, Z. Lactobacillus johnsonii attenuates citrobacter rodentium-induced colitis by regulating inflammatory responses and endoplasmic reticulum stress in mice. J. Nutr. 2021, 151, 3391–3399. [Google Scholar] [CrossRef] [PubMed]
- Charlet, R.; Bortolus, C.; Sendid, B.; Jawhara, S. Bacteroides thetaiotaomicron and Lactobacillus johnsonii modulate intestinal inflammation and eliminate fungi via enzymatic hydrolysis of the fungal cell wall. Sci. Rep. 2020, 10, 11510. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Ni, X.; Qing, X.; Liu, L.; Lai, J.; Khalique, A.; Li, G.; Pan, K.; Jing, B.; Zeng, D. Probiotic enhanced intestinal immunity in broilers against subclinical necrotic enteritis. Front. Immunol. 2017, 8, 1592. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Ni, X.; Qing, X.; Liu, L.; Xin, J.; Luo, M.; Khalique, A.; Dan, Y.; Pan, K.; Jing, B.; et al. Probiotic Lactobacillus johnsonii BS15 improves blood parameters related to immunity in broilers experimentally infected with subclinical necrotic enteritis. Front. Microbiol. 2018, 9, 49. [Google Scholar] [CrossRef] [PubMed]
- Qing, X.; Zeng, D.; Wang, H.; Ni, X.; Liu, L.; Lai, J.; Khalique, A.; Pan, K.; Jing, B. Preventing subclinical necrotic enteritis through Lactobacillus johnsonii BS15 by ameliorating lipid metabolism and intestinal microflora in broiler chickens. AMB Express 2017, 7, 139. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Ni, X.; Qing, X.; Zeng, D.; Luo, M.; Liu, L.; Li, G.; Pan, K.; Jing, B. Live probiotic Lactobacillus johnsonii BS15 promotes growth performance and lowers fat deposition by improving lipid metabolism, intestinal development, and gut microflora in broilers. Front. Microbiol. 2017, 8, 1073. [Google Scholar] [CrossRef]
- Khalique, A.; Zeng, D.; Wang, H.; Qing, X.; Zhou, Y.; Xin, J.; Zeng, Y.; Pan, K.; Shu, G.; Jing, B.; et al. Transcriptome analysis revealed ameliorative effect of probiotic Lactobacillus johnsonii BS15 against subclinical necrotic enteritis induced hepatic inflammation in broilers. Microb. Pathog. 2019, 132, 201–207. [Google Scholar] [CrossRef]
- Xin, J.; Wang, H.; Sun, N.; Bughio, S.; Zeng, D.; Li, L.; Wang, Y.; Khalique, A.; Zeng, Y.; Pan, K.; et al. Probiotic alleviate fluoride-induced memory impairment by reconstructing gut microbiota in mice. Ecotoxicol. Environ. Saf. 2021, 215, 112108. [Google Scholar] [CrossRef]
- Sun, N.; Ni, X.; Wang, H.; Xin, J.; Zhao, Y.; Pan, K.; Jing, B.; Zeng, D. Probiotic Lactobacillus johnsonii BS15 prevents memory dysfunction induced by chronic high-fluorine intake through modulating intestinal environment and improving gut development. Probiotics Antimicrob. Proteins 2020, 12, 1420–1438. [Google Scholar] [CrossRef]
- Wang, H.; He, S.; Xin, J.; Zhang, T.; Sun, N.; Li, L.; Ni, X.; Zeng, D.; Ma, H.; Bai, Y. Psychoactive effects of Lactobacillus johnsonii against restraint stress-induced memory dysfunction in mice through modulating intestinal inflammation and permeability-a study based on the gut-brain axis hypothesis. Front. Pharmacol. 2021, 12, 662148. [Google Scholar] [CrossRef]
- Wang, H.; Sun, Y.; Xin, J.; Zhang, T.; Sun, N.; Ni, X.; Zeng, D.; Bai, Y. Lactobacillus johnsonii BS15 prevents psychological stress-induced memory dysfunction in mice by modulating the gut-brain axis. Front. Microbiol. 2020, 11, 1941. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, L.D.; Torrez Lamberti, M.F.; DeBose-Scarlett, E.; Bahadiroglu, E.; Garrett, T.J.; Gardner, C.L.; Meyer, J.L.; Lorca, G.L.; Gonzalez, C.F. Lactobacillus johnsonii N6.2 and blueberry phytophenols affect lipidome and gut microbiota composition of rats under high-fat diet. Front. Nutr. 2021, 8, 757256. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Hong, E.; Oh, S.; Kim, E. Non-viable Lactobacillus johnsonii JNU3402 protects against diet-induced obesity. Foods 2020, 9, 1494. [Google Scholar] [CrossRef] [PubMed]
- Yoon, H.; Lee, Y.; Park, H.; Kang, H.-J.; Ji, Y.; Holzapfel, W.H. Lactobacillus johnsonii BFE6154 ameliorates diet-induced hypercholesterolemia. Probiotics Antimicrob. Proteins 2021, 15, 451–459. [Google Scholar] [CrossRef] [PubMed]
- Xin, J.; Zeng, D.; Wang, H.; Ni, X.; Yi, D.; Pan, K.; Jing, B. Preventing non-alcoholic fatty liver disease through Lactobacillus johnsonii BS15 by attenuating inflammation and mitochondrial injury and improving gut environment in obese mice. Appl. Microbiol. Biotechnol. 2014, 98, 6817–6829. [Google Scholar] [CrossRef] [PubMed]
- Openshaw, P.J.; Dean, G.S.; Culley, F.J. Links between respiratory syncytial virus bronchiolitis and childhood asthma: Clinical and research approaches. Pediatr. Infect. Dis. J. 2003, 22, S58–S64. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.-M.; Yang, Y.-C.S.H.; Chou, H.-C.; Lin, S. Intranasal administration of Lactobacillus johnsonii attenuates hyperoxia-induced lung injury by modulating gut microbiota in neonatal mice. J. Biomed. Sci. 2023, 30, 57. [Google Scholar] [CrossRef]
- Liévin-Le Moal, V.; Servin, A.L. Anti-infective activities of Lactobacillus strains in the human intestinal microbiota: From probiotics to gastrointestinal anti-infectious biotherapeutic agents. Clin. Microbiol. Rev. 2014, 27, 167–199. [Google Scholar] [CrossRef]
- Bergonzelli, G.E.; Granato, D.; Pridmore, R.D.; Marvin-Guy, L.F.; Donnicola, D.; Corthésy-Theulaz, I.E. GroEL of Lactobacillus johnsonii La1 (NCC 533) is cell surface associated: Potential role in interactions with the host and the gastric pathogen Helicobacter pylori. Infect. Immun. 2006, 74, 425–434. [Google Scholar] [CrossRef]
- Avonts, L.; De Vuyst, L. Antimicrobial potential of probiotic lactic acid bacteria. Mededelingen (Rijksuniv. Te Gent. Fak. Van Landbouwkd. En Toegepaste Biol. Wet.) 2001, 66, 543–550. [Google Scholar]
- Makras, L.; Triantafyllou, V.; Fayol-Messaoudi, D.; Adriany, T.; Zoumpopoulou, G.; Tsakalidou, E.; Servin, A.; De Vuyst, L. Kinetic analysis of the antibacterial activity of probiotic Lactobacilli towards salmonella enterica serovar typhimurium reveals a role for lactic acid and other inhibitory compounds. Res. Microbiol. 2006, 157, 241–247. [Google Scholar] [CrossRef]
- Chen, K.; Wang, J.; Guo, L.; Wang, J.; Yang, L.; Hu, T.; Zhao, Y.; Wang, X.; Zhu, Y. Lactobacillus johnsonii L531 ameliorates Salmonella enterica Serovar Typhimurium diarrhea by modulating iron homeostasis and oxidative stress via the IRP2 pathway. Nutrients 2023, 15, 1127. [Google Scholar] [CrossRef]
- Skinner, J.T.; Bauer, S.; Young, V.; Pauling, G.; Wilson, J. An economic analysis of the impact of subclinical (mild) necrotic enteritis in broiler chickens. Avian Dis. 2010, 54, 1237–1240. [Google Scholar] [CrossRef]
- Nii, T.; Shinkoda, T.; Isobe, N.; Yoshimura, Y. Intravaginal injection of Lactobacillus johnsonii may modulates oviductal microbiota and mucosal barrier function of laying hens. Poult. Sci. 2023, 102, 102699. [Google Scholar] [CrossRef]
- Schmidt, C. Thinking from the gut. Nature 2015, 518, S12–S14. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, A.; Harty, S.; Lehto, S.M.; Moeller, A.H.; Dinan, T.G.; Dunbar, R.I.M.; Cryan, J.F.; Burnet, P.W.J. The microbiome in psychology and cognitive neuroscience. Trends Cogn. Sci. 2018, 22, 611–636. [Google Scholar] [CrossRef] [PubMed]
- Knierim, J.J. The hippocampus. Curr. Biol. 2015, 25, R1116–R1121. [Google Scholar] [CrossRef] [PubMed]
- Jang, S.-E.; Lim, S.-M.; Jeong, J.-J.; Jang, H.-M.; Lee, H.-J.; Han, M.J.; Kim, D.-H. Gastrointestinal inflammation by gut microbiota disturbance induces memory impairment in mice. Mucosal Immunol. 2018, 11, 369–379. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Duan, Y.; Li, R.; Liang, X.; Li, T.; Huang, X.; Yin, Y.; Yin, J. Gut microbial profiles and the role in lipid metabolism in shaziling pigs. Anim. Nutr. 2022, 9, 345–356. [Google Scholar] [CrossRef] [PubMed]
- Yamano, T.; Tanida, M.; Niijima, A.; Maeda, K.; Okumura, N.; Fukushima, Y.; Nagai, K. Effects of the probiotic strain Lactobacillus johnsonii strain La1 on autonomic nerves and blood glucose in rats. Life Sci. 2006, 79, 1963–1967. [Google Scholar] [CrossRef] [PubMed]
- Kingma, S.D.K.; Li, N.; Sun, F.; Valladares, R.B.; Neu, J.; Lorca, G.L. Lactobacillus johnsonii N6.2 stimulates the innate immune response through toll-like receptor 9 in Caco-2 cells and increases intestinal crypt paneth cell number in biobreeding diabetes-prone rats. J. Nutr. 2011, 141, 1023–1028. [Google Scholar] [CrossRef] [PubMed]
- Friedman, S.L.; Neuschwander-Tetri, B.A.; Rinella, M.; Sanyal, A.J. Mechanisms of NAFLD development and therapeutic strategies. Nat. Med. 2018, 24, 908–922. [Google Scholar] [CrossRef] [PubMed]
- Powell, E.E.; Wong, V.W.-S.; Rinella, M. Non-alcoholic fatty liver disease. Lancet 2021, 397, 2212–2224. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, R.R.; Gurung, M.; Li, Z.; García-Jaramillo, M.; Greer, R.; Gaulke, C.; Bauchinger, F.; You, H.; Pederson, J.W.; Vasquez-Perez, S.; et al. Transkingdom interactions between Lactobacilli and hepatic mitochondria attenuate western diet-induced diabetes. Nat. Commun. 2021, 12, 101. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.; Wu, F.; Zhang, F.; Li, X.; Wu, X.; Fu, J. Hepatoenteric protective effect of melanin from Inonotus hispidus on acute alcoholic liver injury in mice. Mol. Nutr. Food Res. 2023, 67, e2200562. [Google Scholar] [CrossRef] [PubMed]
- Vazquez-Munoz, R.; Thompson, A.; Russell, J.T.; Sobue, T.; Zhou, Y.; Dongari-Bagtzoglou, A. Insights from the Lactobacillus johnsonii genome suggest the production of metabolites with antibiofilm activity against the pathobiont candida albicans. Front. Microbiol. 2022, 13, 853762. [Google Scholar] [CrossRef]
- ElFeky, D.S.; Awad, A.R.; Shamseldeen, A.M.; Mowafy, H.L.; Hosny, S.A. Comparing the therapeutic potentials of Lactobacillus johnsonii vs. Lactobacillus acidophilus against vulvovaginal candidiasis in female rats: An in vivo study. Front. Microbiol. 2023, 14, 1222503. [Google Scholar] [CrossRef]
- Ahire, J.J.; Sahoo, S.; Kashikar, M.S.; Heerekar, A.; Lakshmi, S.G.; Madempudi, R.S. In vitro assessment of Lactobacillus crispatus UBLCp01, Lactobacillus gasseri UBLG36, and Lactobacillus johnsonii UBLJ01 as a potential vaginal probiotic candidate. Probiotics Antimicrob. Proteins 2023, 15, 275–286. [Google Scholar] [CrossRef]
- Liu, R.; Zhou, Y.; Chen, H.; Xu, H.; Zuo, M.; Chen, B.; Wang, H. Membrane vesicles from Lactobacillus johnsonii delay osteoarthritis progression via modulating macrophage glutamine synthetase/MTORC1 axis. Biomed. Pharmacother. 2023, 165, 115204. [Google Scholar] [CrossRef]
- Zhong, X.; Zhao, Y.; Huang, L.; Liu, J.; Wang, K.; Gao, X.; Zhao, X.; Wang, X. Remodeling of the gut microbiome by Lactobacillus johnsonii alleviates the development of acute myocardial infarction. Front. Microbiol. 2023, 14, 1140498. [Google Scholar] [CrossRef]
- Dang, A.T.; Marsland, B.J. Microbes, metabolites, and the gut-lung axis. Mucosal Immunol. 2019, 12, 843–850. [Google Scholar] [CrossRef] [PubMed]
- Carlet, J.; Jarlier, V.; Harbarth, S.; Voss, A.; Goossens, H.; Pittet, D.; The Participants of the 3rd World Healthcare-Associated Infections Forum. Ready for a World without Antibiotics? The pensières antibiotic resistance call to action. Antimicrob. Resist. Infect. Control 2012, 1, 11. [Google Scholar] [CrossRef] [PubMed]
- Global Burden of Bacterial Antimicrobial Resistance in 2019: A Systematic Analysis. Lancet 2022, 399. [CrossRef]
- Boucard, A.-S.; Florent, I.; Polack, B.; Langella, P.; Bermúdez-Humarán, L.G. Genome sequence and assessment of safety and potential probiotic traits of Lactobacillus johnsonii CNCM I-4884. Microorganisms 2022, 10, 273. [Google Scholar] [CrossRef] [PubMed]
- Adedeji, O.E.; Chae, S.A.; Ban, O.-H.; Bang, W.Y.; Kim, H.; Jeon, H.J.; Chinma, C.E.; Yang, J.; Jung, Y.H. Safety evaluation and anti-inflammatory activity of Lactobacillus johnsonii IDCC 9203 isolated from feces of breast-fed infants. Arch Microbiol 2022, 204, 470. [Google Scholar] [CrossRef] [PubMed]
- Gotteland, M.; Cires, M.J.; Carvallo, C.; Vega, N.; Ramirez, M.A.; Morales, P.; Rivas, P.; Astudillo, F.; Navarrete, P.; Dubos, C.; et al. Probiotic screening and safety evaluation of Lactobacillus strains from plants, artisanal goat cheese, human stools, and breast milk. J. Med. Food 2014, 17, 487–495. [Google Scholar] [CrossRef] [PubMed]
- Ma, N.; Guo, P.; Chen, J.; Qi, Z.; Liu, C.; Shen, J.; Sun, Y.; Chen, X.; Chen, G.-Q.; Ma, X. Poly-β-hydroxybutyrate alleviated diarrhea and colitis via Lactobacillus johnsonii biofilm-mediated maturation of sulfomucin. Sci. China Life Sci. 2022, 66, 1569–1588. [Google Scholar] [CrossRef]



| Strain | Disease | Experimental Models | Duration of Intervention | Treatment Results | Reference |
|---|---|---|---|---|---|
| La1 (NCC533) | AD | Atopic dermatitis NC/Nga mice | From 20 to 22 days of age | IL-8↓, IL-12↓, IL-23↓, CD86↓ | [32] |
| N6.2 | T1D | BB-DP rats | Pre-weaning to 1 day old during mother feeding and post-weaning at 21 days old | iNOS↓, IFNγ↓, Cox-2↑, claudin↑, occludin↓ | [33] |
| N6.2 | T1D | BB-DP rats | Daily until sacrifice at diabetes onset, or the culmination of the experiment at 140 days | IL-17↑, IL-23↑ | [34] |
| N6.2 | — | BB-DP rats | After weaning to 60 days of age | mature caspase-1↓, IL-1β↓ | [35] |
| La1 | Hyperglycemia | STZ-induced diabetes animal model and hyperglycemia model induced by intracranial injection of 2DG | 2 weeks | plasma glucose↓, glucagon levels↓ | [34] |
| A multi-strain probiotic supplement including L. johnsonii MH-68 | T1D | Patients with T1D | 24 weeks | fasting blood glucose↓, HbA1c↓, IL-8↓, IL-17↓, MIP-1β↓, RANTES↓, TNF-α↓, TGF-β1↑ | [36] |
| A multi-strain probiotic supplement including L. johnsonii MH-68 | T2D | STZ -induced diabetes animal model | 8 weeks | TNF-α↓, IL-6↓, IL-1β↓, β-cell mass↑ | [37] |
| L. johnsonii | Allergic or infectious airway disease | A similar experimental design as the CRA airway challenge model and RSV infection model | — | IL-4↓, IL-5↓, IL-13↓, IL-17↓, CD11c/CD11b and CD11c/CD8, as well as CD69 activated CD4 and CD8 T cells↓ | [38] |
| L. johnsonii | RSV | RSV infection model | 1 week | IL-4↓, IL-5↓, IL-13↓, IL-6↓, IL-1b↓, TNFα↓, IFNβ↑, DHA↑, AcedoPC↑ | [14] |
| L. johnsonii | RSV | RSV infection model | — | IL-4↓, IL-5↓, IL-13↓, IL-17↓, Gob5(mucogenic gene mRNA level) ↓Th2↓, IFN-γ↑ | [12] |
| La1 | H. pylori-associated gastritis | Healthy adult volunteers of both genders infected by H. pylori | 3 weeks | — | [39] |
| La1 | H. pylori-associated gastritis | Healthy adult volunteers of both genders infected by H. pylori | 2 weeks | δ13CO2 over baseline (DOB)↓ | [40] |
| La1 | H. pylori infection | Healthy adult volunteers of both genders infected by H. pylori | 2 weeks | — | [41] |
| La1 | H. pylori infection | Asymptomatic school children | 4 weeks | — | [42] |
| La1 | H. pylori-associated gastritis | H. pylori SS1 strain infection mode in C57BL/6 mice | 3 months | anti-H. pylori IgG antibody titers↓ | [43] |
| No. 1088 | H. pylori infection | Male germ-free Balb/c mice | 2 weeks or 4 weeks | — | [44] |
| MH-68 | H. pylori infection | SPF SD male rats | 4 weeks | — | [45] |
| L. johnsonii | H. pylori-associated gastritis | Healthy adult volunteers of both genders infected by H. pylori | 16 weeks | H. pylori density↓ | [46] |
| NJ13 | Enterohaemorrhagic E. coli-induced diarrhoea | Female mice | — | — | [47] |
| LJ1 | Enteroinvasive E. coli-induced diarrhea | KM mice | 8-22days | — | [48] |
| L. johnsonii | UC | DSS-induced chronic colitis mice model and human sample | — | IL10↑, TLR1/2↑, MRC1↑ | [49] |
| La1 (NCC533) | — | Completely enterally fed elderly in-patients aged over 70 years | 12 weeks | serum albumin↑, Blood Hb↑, blood phagocytic activity↑, TNF-α↓ | [50] |
| L. johnsonii | Rodentium-Induced colitis | female C57BL/6J mice | 2 weeks | CD4, CD8, CD11b, F4/80, TNF-α, IL-1β, IL-6, IL-17A, ssMCP1, Cox2↓ | [51] |
| L. johnsonii + B. thetaiotaomicron | Colitis | DSS-induced chronic colitis mice model | 5 days | IgA↑, IL-1β↓, IL-10↑, TLR9↑, TLR↓, MBL-C↓ | [52] |
| BS15 | SNE | Broiler chickens (Cobb 500) | days 1-28 or days 29-42 | CD4↑, CD4/CD8↑, sIgA in ileum↑ | [53] |
| BS15 | SNE | Broiler chickens (Cobb 500) | 28days or 42days | SOD↑, CAT↑, IHR↑, T-AOC↑, IgG↑ and IgA↑ in serum, IFN-γ↑, CD3CD4 percentage↑, CD3CD4/CD3CD8↑ | [54] |
| BS15 | SNE | Broiler chickens (Cobb 500) | 4 weeks | ALT↓, AST↓, TC↓, HDL-C↑, PPARγ and ATGL↑ in adipose tissue, IGF-1 and EGF↑ in jejunum and ileum, ACC, FAS and SREBP-1c↓ inhepatic expressions, PPARα and CPT-1↑ in hepatic expressions | [55] |
| BS15 | SNE | Broiler chickens (Cobb 500) | 6 weeks | HDL-C↑, TG↓, LDL-C↓, SREBP-1c and FAS↓ in hepatic expressions | [56] |
| BS15 | SNE induced hepatic inflammation | Broiler chickens (Cobb 500) | 4 weeks | FOS↓ | [57] |
| BS15 | Fluoride-induced memory impairment | Male ICR mice | 98 days | BDNF↑, CREB↑, Bcl-xl↑, Bad↓ | [58] |
| BS15 | Memory dysfunction Induced by chronic high-fluorine intake | Male ICR mice | 98 days | mRNA levels of Dbn, MAP-2, and SYP↑, T-AOC, and GSH-Px↑ in hippocampuls, sIgA ↓ in the jejunal mucosa, MDA↑, SOD↑, CAT activities↑, GSH↑ | [59] |
| BS15 | Memory dysfunction in mice after RS | 5C7BL/6J male mice | 4 weeks | the mRNA expression levels of BDNF, CREB, SCF, c-Fos, and NMDAR↑, DA, 5-HT, and Ach levels↑, the mRNA expression level of IL-4↑, GABA↑, mRNA expression levels of bcl-2 and Bcl-xL↑ | [60] |
| BS15 | Psychological stress-induced memory dysfunction | WAS in ICR male mice | 4 weeks | mRNA-expression levels of tight junction proteins claudin-1, occludin, and ZO-1 in the jejunum and ileum↑, TNF-α↓, IFN-γ↓, and IL-1β↓, mRNA levels of BDNF↑, CREB↑ | [61] |
| N6.2 and BB | Diet-induced obesity | HFD model | 15 weeks | SREBP-1↓, SCAP↓, LCFA in the serum↑ | [62] |
| JNU3402 | Diet-induced obesity | HFD model C57BL/6J mice | 14 weeks | ACOX↑, CPT1↑, PGC1α↑, PPARγ↑, TG↓, FAS↓, ACC↓, SREBP1c↓, hepatic cholesterol level↓ | [63] |
| BFE6154 | Diet-induced hypercholesterolemia | HFHCD model C57BL/6J mice | 4 weeks | LDL↓, ABCG8↓, NPC1L1↓, ABCG5↑, HDL↑, LDLR↑ | [64] |
| BS15 | NAFLD | HFD model ICR mice | 17 weeks | LPS↓, TG↓, LDLC↓, ALT↓, FFA↓, HDLC↓, UCP-2 and cytochrome c↓ in mitochondria, the hepatic expression of Acc 1, Fas, TNFα and PPARγ↓, the hepatic expression of Fiaf↑ | [65] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Z.; Zhao, L.; Wu, J.; Pan, Y.; Zhao, G.; Li, Z.; Zhang, L. The Effects of Lactobacillus johnsonii on Diseases and Its Potential Applications. Microorganisms 2023, 11, 2580. https://doi.org/10.3390/microorganisms11102580
Zhang Z, Zhao L, Wu J, Pan Y, Zhao G, Li Z, Zhang L. The Effects of Lactobacillus johnsonii on Diseases and Its Potential Applications. Microorganisms. 2023; 11(10):2580. https://doi.org/10.3390/microorganisms11102580
Chicago/Turabian StyleZhang, Ziyi, Lanlan Zhao, Jiacheng Wu, Yingmiao Pan, Guoping Zhao, Ziyun Li, and Lei Zhang. 2023. "The Effects of Lactobacillus johnsonii on Diseases and Its Potential Applications" Microorganisms 11, no. 10: 2580. https://doi.org/10.3390/microorganisms11102580