
Anti-Toxin Responses to Natural Enterotoxigenic Escherichia coli (ETEC) Infection in Adults and Children in Bangladesh
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design
2.2. Sample Collection
2.3. ELISA
2.4. Toxin Neutralization
2.5. Linear Peptide Epitope Mapping
3. Results
3.1. Study Participants
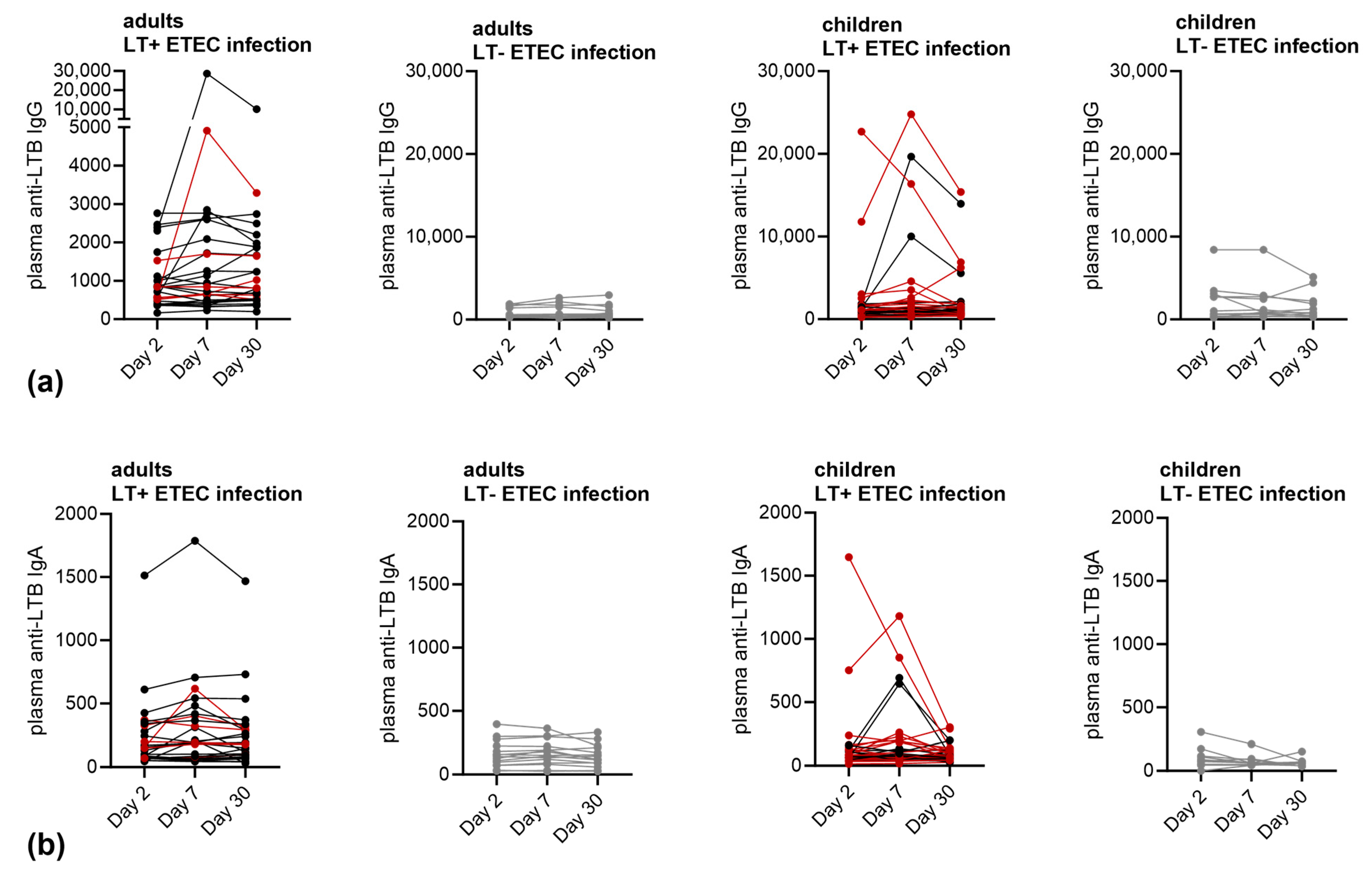
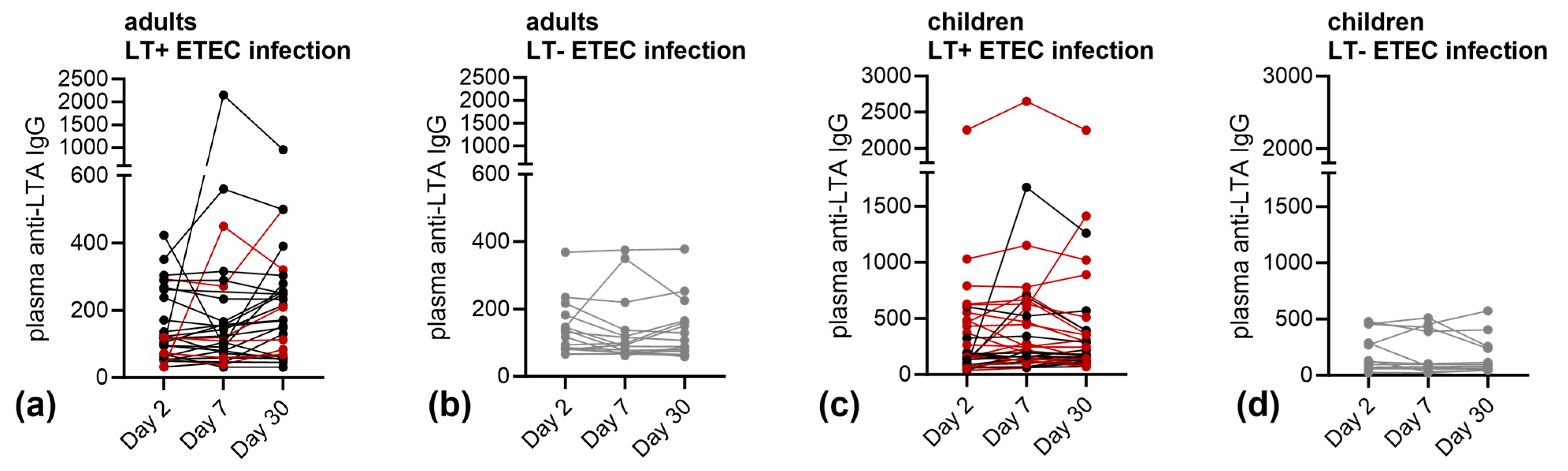
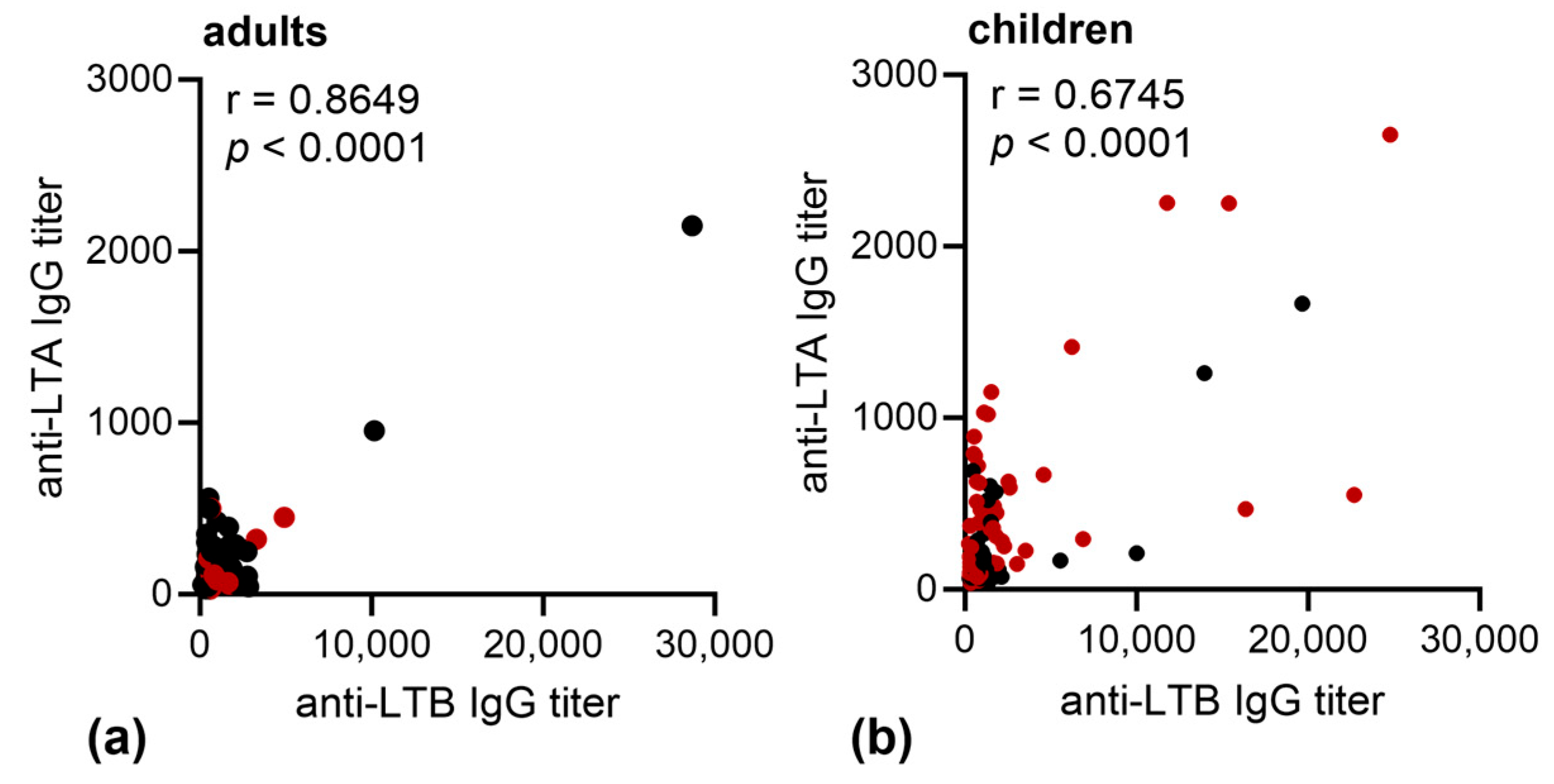
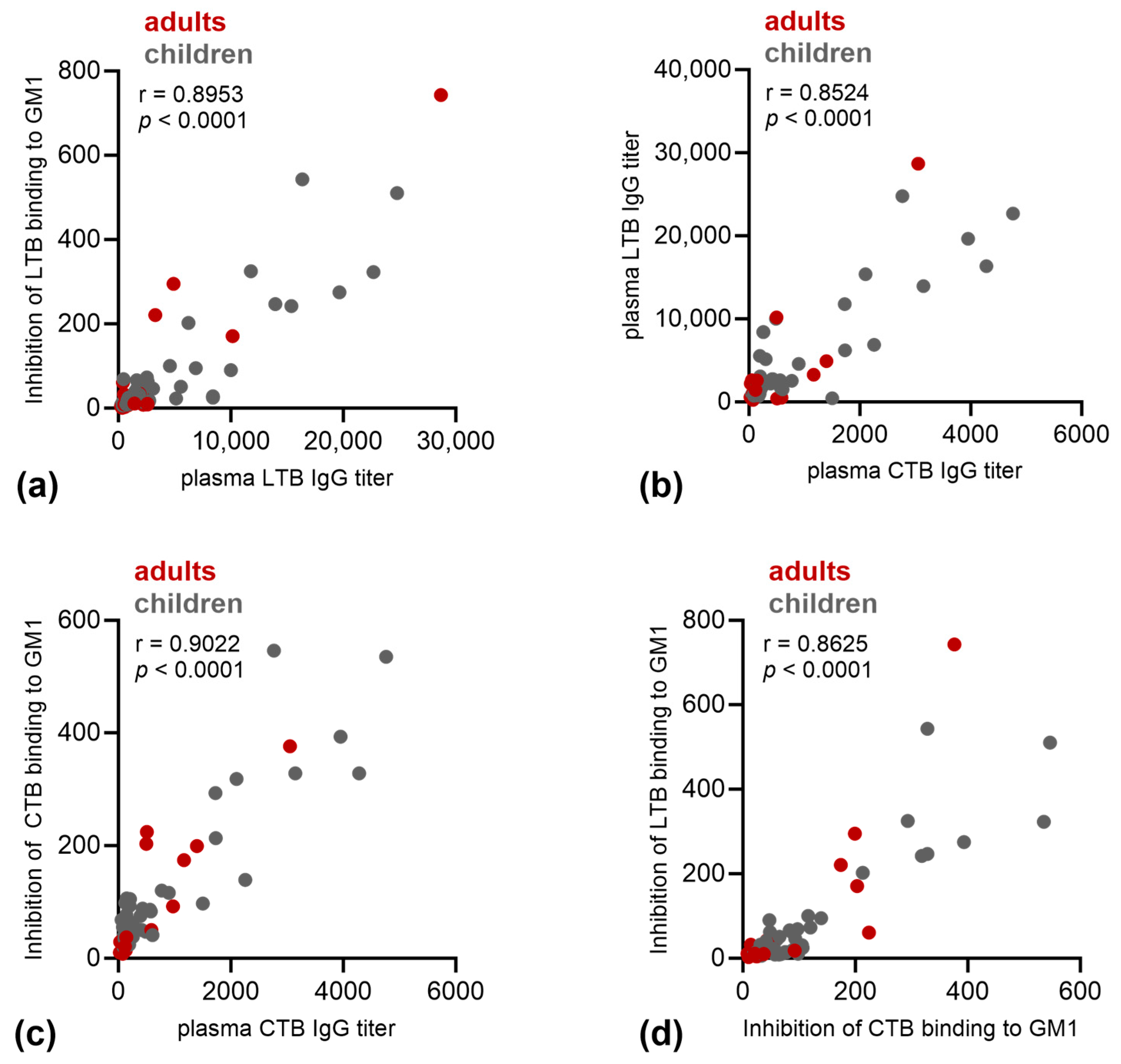
3.2. Immune Responses against LT
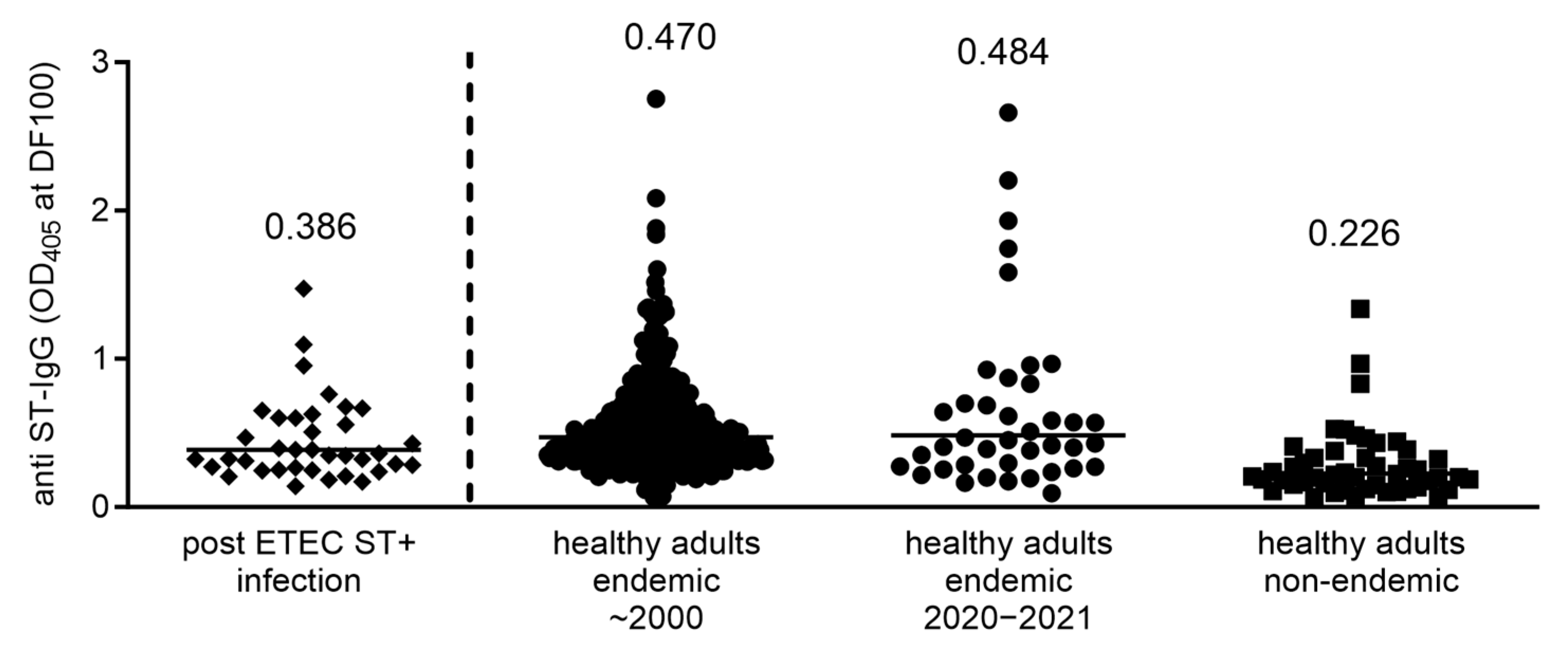
3.3. Immune Responses against ST
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kotloff, K.L.; Nataro, J.P.; Blackwelder, W.C.; Nasrin, D.; Farag, T.H.; Panchalingam, S.; Wu, Y.; Sow, S.O.; Sur, D.; Breiman, R.F.; et al. Burden and aetiology of diarrhoeal disease in infants and young children in developing countries (the Global Enteric Multicenter Study, GEMS): A prospective, case-control study. Lancet 2013, 382, 209–222. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Platts-Mills, J.A.; Juma, J.; Kabir, F.; Nkeze, J.; Okoi, C.; Operario, D.J.; Uddin, J.; Ahmed, S.; Alonso, P.L.; et al. Use of quantitative molecular diagnostic methods to identify causes of diarrhoea in children: A reanalysis of the GEMS case-control study. Lancet 2016, 388, 1291–1301. [Google Scholar] [CrossRef] [PubMed]
- Troeger, C.; Forouzanfar, M.; Rao, P.C.; Khalil, I.; Brown, A.; Reiner, R.C.; Fullman, N.; Thompson, R.; Abajobir, A.; Ahmed, M.; et al. Estimates of global, regional, and national morbidity, mortality, and aetiologies of diarrhoeal diseases: A systematic analysis for the Global Burden of Disease Study 2015. Lancet Infect. Dis. 2017, 17, 909–948. [Google Scholar] [CrossRef] [PubMed]
- George, C.M.; Burrowes, V.; Perin, J.; Oldja, L.; Biswas, S.; Sack, D.; Ahmed, S.; Haque, R.; Bhuiyan, N.A.; Parvin, T.; et al. Enteric Infections in Young Children are Associated with Environmental Enteropathy and Impaired Growth. Trop. Med. Int. Health 2018, 23, 26–33. [Google Scholar] [CrossRef] [PubMed]
- Mani, S.; Wierzba, T.; Walker, R.I. Status of vaccine research and development for Shigella. Vaccine 2016, 34, 2887–2894. [Google Scholar] [CrossRef] [PubMed]
- Bourgeois, A.L.; Wierzba, T.F.; Walker, R.I. Status of vaccine research and development for enterotoxigenic Escherichia coli. Vaccine 2016, 34, 2880–2886. [Google Scholar] [CrossRef]
- Spangler, B.D. Structure and function of cholera toxin and the related Escherichia coli heat-labile enterotoxin. Microbiol. Rev. 1992, 56, 622–647. [Google Scholar] [CrossRef] [PubMed]
- Fleckenstein, J.M.; Hardwidge, P.R.; Munson, G.P.; Rasko, D.A.; Sommerfelt, H.; Steinsland, H. Molecular mechanisms of enterotoxigenic Escherichia coli infection. Microbes Infect. 2010, 12, 89–98. [Google Scholar] [CrossRef]
- Horstman, A.L.; Kuehn, M.J. Enterotoxigenic Escherichia coli Secretes Active Heat-Labile Enterotoxin via Outer Membrane Vesicles. J. Biol. Chem. 2000, 275, 12489–12496. [Google Scholar] [CrossRef]
- Jelinek, T.; Kollaritsch, H. Vaccination with Dukoral against travelers’ diarrhea (ETEC) and cholera. Expert Rev. Vaccines 2008, 7, 561–567. [Google Scholar] [CrossRef]
- Zhang, W.; Zhang, C.; Francis, D.H.; Fang, Y.; Knudsen, D.; Nataro, J.P.; Robertson, D.C. Genetic Fusions of Heat-Labile (LT) and Heat-Stable (ST) Toxoids of Porcine Enterotoxigenic Escherichia coli Elicit Neutralizing Anti-LT and Anti-STa antibodies. Infect. Immun. 2010, 78, 316–325. [Google Scholar] [CrossRef] [PubMed]
- Harutyunyan, S.; Neuhauser, I.; Mayer, A.; Aichinger, M.; Szijártó, V.; Nagy, G.; Nagy, E.; Girardi, P.; Malinoski, F.J.; Henics, T. Characterization of ShigETEC, a Novel Live Attenuated Combined Vaccine against Shigellae and ETEC. Vaccines 2020, 8, 689. [Google Scholar] [CrossRef] [PubMed]
- Guan, W.; Liu, W.; Bao, J.; Li, J.; Yuan, C.; Tang, J.; Shi, D. Analysis and application of a neutralizing linear epitope on liable toxin B of enterotoxin Escherichia coli. Appl. Microbiol. Biotechnol. 2015, 99, 5985–5996. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, I.; Kiyono, H.; Jackson, R.J.; Fujihashi, K.; Staats, H.F.; Hamada, S.; Clements, J.D.; Bost, K.L.; McGhee, J.R. Epitope maps of the Escherichia coli heat-labile toxin B subunit for development of a synthetic oral vaccine. Infect. Immun. 1996, 64, 1290–1298. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Duan, Q.; Zhang, W. Significance of Enterotoxigenic Escherichia coli (ETEC) Heat-Labile Toxin (LT) Enzymatic Subunit Epitopes in LT Enterotoxicity and Immunogenicity. Appl. Environ. Microbiol. 2018, 84, e00849-18. [Google Scholar] [CrossRef] [PubMed]
- Walker, R.; Kaminski, R.W.; Porter, C.; Choy, R.K.M.; White, J.A.; Fleckenstein, J.M.; Cassels, F.; Bourgeois, L. Vaccines for Protecting Infants from Bacterial Causes of Diarrheal Disease. Microorganisms 2021, 9, 1382. [Google Scholar] [CrossRef] [PubMed]
- Platts-Mills, J.A.; Babji, S.; Bodhidatta, L.; Gratz, J.; Haque, R.; Havt, A.; McCormick, B.J.; McGrath, M.; Olortegui, M.P.; Samie, A.; et al. Pathogen-specific burdens of community diarrhoea in developing countries: A multisite birth cohort study (MAL-ED). Lancet Glob. Health 2015, 3, e564–e575. [Google Scholar] [CrossRef]
- Turunen, K.; Antikainen, J.; Lääveri, T.; Kirveskari, J.; Svennerholm, A.-M.; Kantele, A. Clinical aspects of heat-labile and heat-stable toxin-producing enterotoxigenic Escherichia coli: A prospective study among Finnish travellers. Travel Med. Infect. Dis. 2020, 38, 101855. [Google Scholar] [CrossRef]
- Paschke, C.; Apelt, N.; Fleischmann, E.; Perona, P.; Walentiny, C.; Löscher, T.; Herbinger, K.-H. Controlled study on enteropathogens in travellers returning from the tropics with and without diarrhoea. Clin. Microbiol. Infect. 2011, 17, 1194–1200. [Google Scholar] [CrossRef]
- Steinsland, H.; Valentiner-Branth, P.; Gjessing, H.K.; Aaby, P.; Mølbak, K.; Sommerfelt, H. Protection from natural infections with enterotoxigenic Escherichia coli: Longitudinal study. Lancet 2003, 362, 286–291. [Google Scholar] [CrossRef]
- Girardi, P.; Harutyunyan, S.; Neuhauser, I.; Glaninger, K.; Korda, O.; Nagy, G.; Nagy, E.; Szijártó, V.; Pall, D.; Szarka, K.; et al. Evaluation of the Safety, Tolerability and Immunogenicity of ShigETEC, an Oral Live Attenuated Shigella-ETEC Vaccine in Placebo-Controlled Randomized Phase 1 Trial. Vaccines 2022, 10, 340. [Google Scholar] [CrossRef] [PubMed]
- Zegeye, E.D.; Govasli, M.L.; Sommerfelt, H.; Puntervoll, P. Development of an enterotoxigenic Escherichia coli vaccine based on the heat-stable toxin. Hum. Vaccines Immunother. 2019, 15, 1379–1388. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Ruan, X.; Zhang, C.; Lawson, S.R.; Knudsen, D.E.; Nataro, J.P.; Robertson, D.C.; Zhang, W. Heat-Labile- and Heat-Stable-Toxoid Fusions (LT R192G -STa P13F) of Human Enterotoxigenic Escherichia coli Elicit Neutralizing Antitoxin Antibodies. Infect. Immun. 2011, 79, 4002–4009. [Google Scholar] [CrossRef] [PubMed]
- Ruan, X.; Robertson, D.C.; Nataro, J.P.; Clements, J.D.; Zhang, W. Characterization of Heat-Stable (STa) Toxoids of Enterotoxigenic Escherichia coli Fused to Double Mutant Heat-Labile Toxin Peptide in Inducing Neutralizing Anti-STa Antibodies. Infect. Immun. 2014, 82, 1823–1832. [Google Scholar] [CrossRef] [PubMed]
- Klipstein, F.; Engert, R.; Houghten, R. Immunisation of Volunteers with A Synthetic Peptide Vaccine for Enterotoxigenic Escherichia coli. Lancet 1986, 327, 471–473. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gender | Age | |||||
|---|---|---|---|---|---|---|
| Male Number (%) | Female Number (%) | 1–11 Months Number (%) | 12–23 Months Number (%) | 2–5 Years Number (%) | 18–45 Years Number (%) | |
| Children (n = 46) | 26 | 20 | 5 | 14 | 27 | |
| (56.5%) | (43.5%) | (10.9%) | (30.4%) | (58.7%) | ||
| Adults (n = 43) | 17 | 26 | 43 | |||
| (39.5%) | (60.5%) | (100%) | ||||
| Infection | |||||
|---|---|---|---|---|---|
| LT+/ST− Number (%) | LT+/STh+ Number (%) | LT+/STp+ Number (%) | STh+ Number (%) | STp+ Number (%) | |
| Children (n = 46) | 19 | 9 | 6 | 7 | 5 |
| (41.3%) | (19.6%) | (13%) | (15.2%) | (10.9%) | |
| Adults (n = 43) | 6 | 21 | 1 | 10 | 5 |
| (14%) | (48.8%) | (2.3%) | (23.3%) | (11.6%) | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Girardi, P.; Bhuiyan, T.R.; Lundin, S.B.; Harutyunyan, S.; Neuhauser, I.; Khanam, F.; Nagy, G.; Szijártó, V.; Henics, T.; Nagy, E.; et al. Anti-Toxin Responses to Natural Enterotoxigenic Escherichia coli (ETEC) Infection in Adults and Children in Bangladesh. Microorganisms 2023, 11, 2524. https://doi.org/10.3390/microorganisms11102524
Girardi P, Bhuiyan TR, Lundin SB, Harutyunyan S, Neuhauser I, Khanam F, Nagy G, Szijártó V, Henics T, Nagy E, et al. Anti-Toxin Responses to Natural Enterotoxigenic Escherichia coli (ETEC) Infection in Adults and Children in Bangladesh. Microorganisms. 2023; 11(10):2524. https://doi.org/10.3390/microorganisms11102524
Chicago/Turabian StyleGirardi, Petra, Taufiqur Rahman Bhuiyan, Samuel B. Lundin, Shushan Harutyunyan, Irene Neuhauser, Farhana Khanam, Gábor Nagy, Valéria Szijártó, Tamás Henics, Eszter Nagy, and et al. 2023. "Anti-Toxin Responses to Natural Enterotoxigenic Escherichia coli (ETEC) Infection in Adults and Children in Bangladesh" Microorganisms 11, no. 10: 2524. https://doi.org/10.3390/microorganisms11102524