

Bioprospecting of Actinobacterial Diversity and Antibacterial Secondary Metabolites from the Sediments of Four Saline Lakes on the Northern Tibetan Plateau

 , , , , , ,
, , , , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
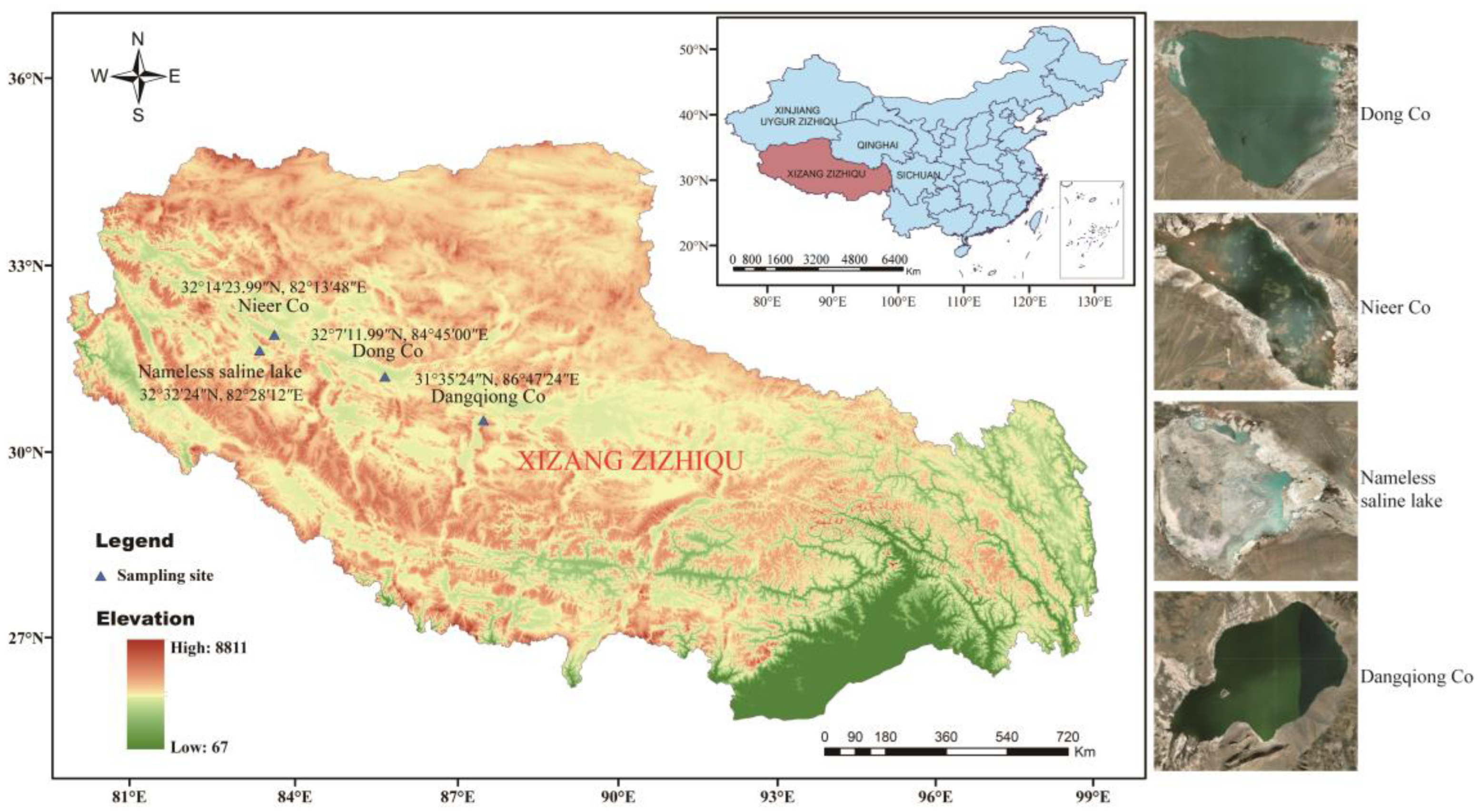
2.1. Samples Collection and Preparation
2.2. Actinobacteria Isolation
2.3. Molecular Identification and Phylogenetic Analysis
2.4. MATRIX Cultivation and Antimicrobial Activity Assay
2.5. Determination of Antibacterial Mechanism
2.6. Scale-Up Cultivation and Antimicrobial Tests with Paper-Disk Diffusion Method
2.7. UPLC/ESI-QTOF-MS/MS Analyses
2.8. Molecular Networking and Chemical Prediction
2.9. Genome Sequencing and Biosynthesis Gene Clusters (BGCs) Identification
3. Results
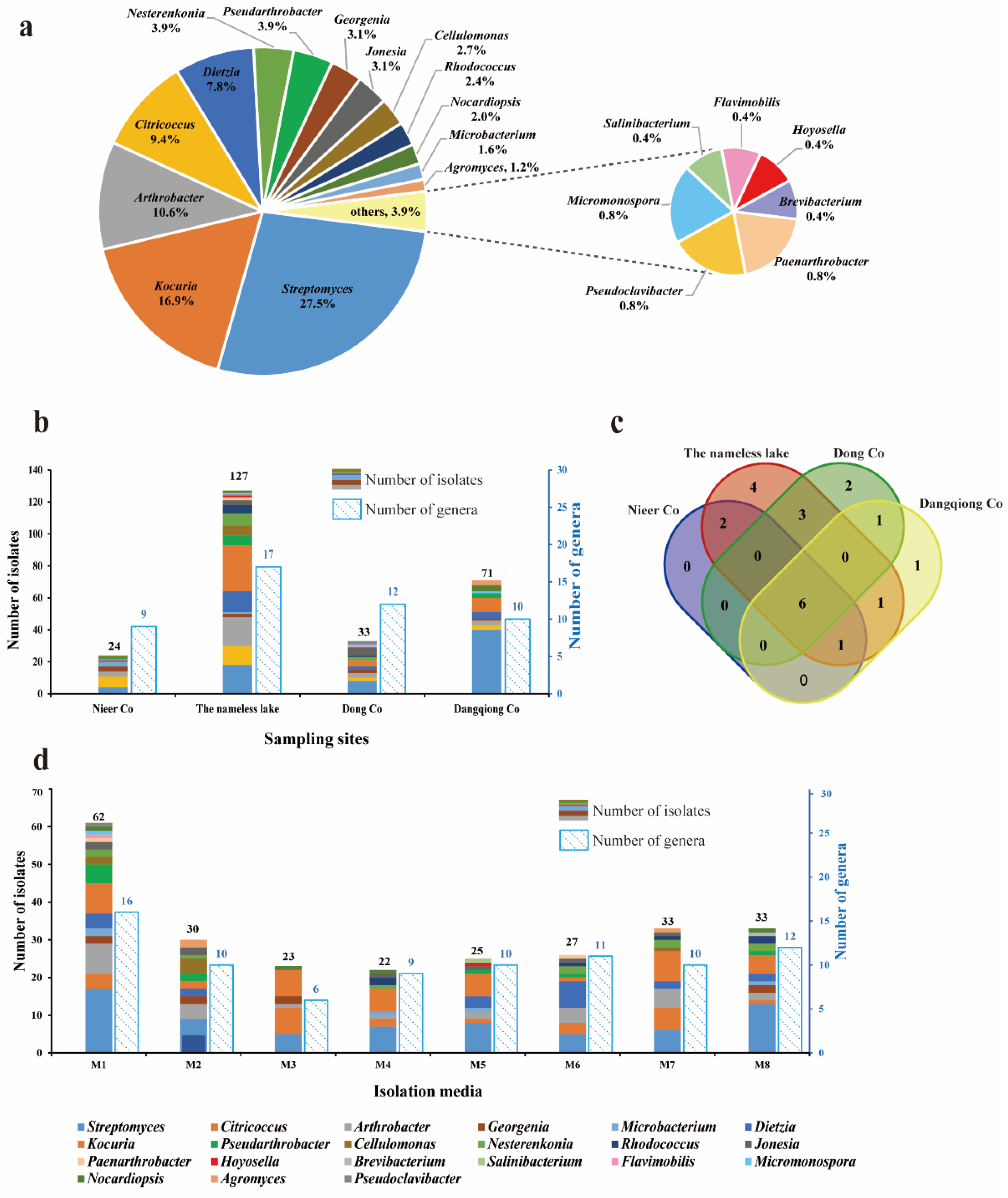
3.1. Taxonomy and Biodiversity of Cultivable Actinobacteria
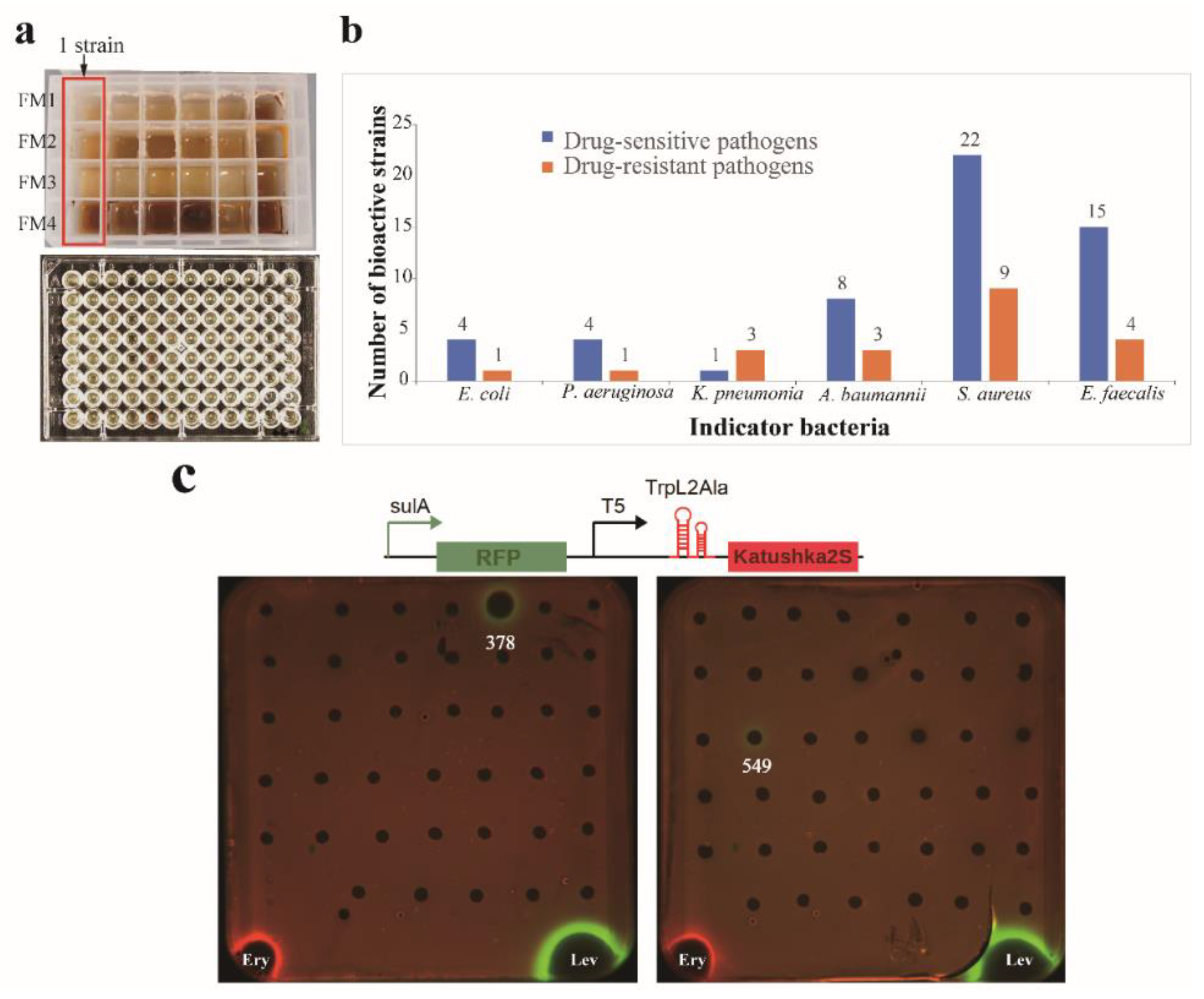
3.2. MATRIX Cultivation and Antibacterial Activity
3.3. Assay of Antibacterial Mechanism Action
3.4. Scale-up Cultivation and Antimicrobial Activity Revaluation of Strains 378 and 549
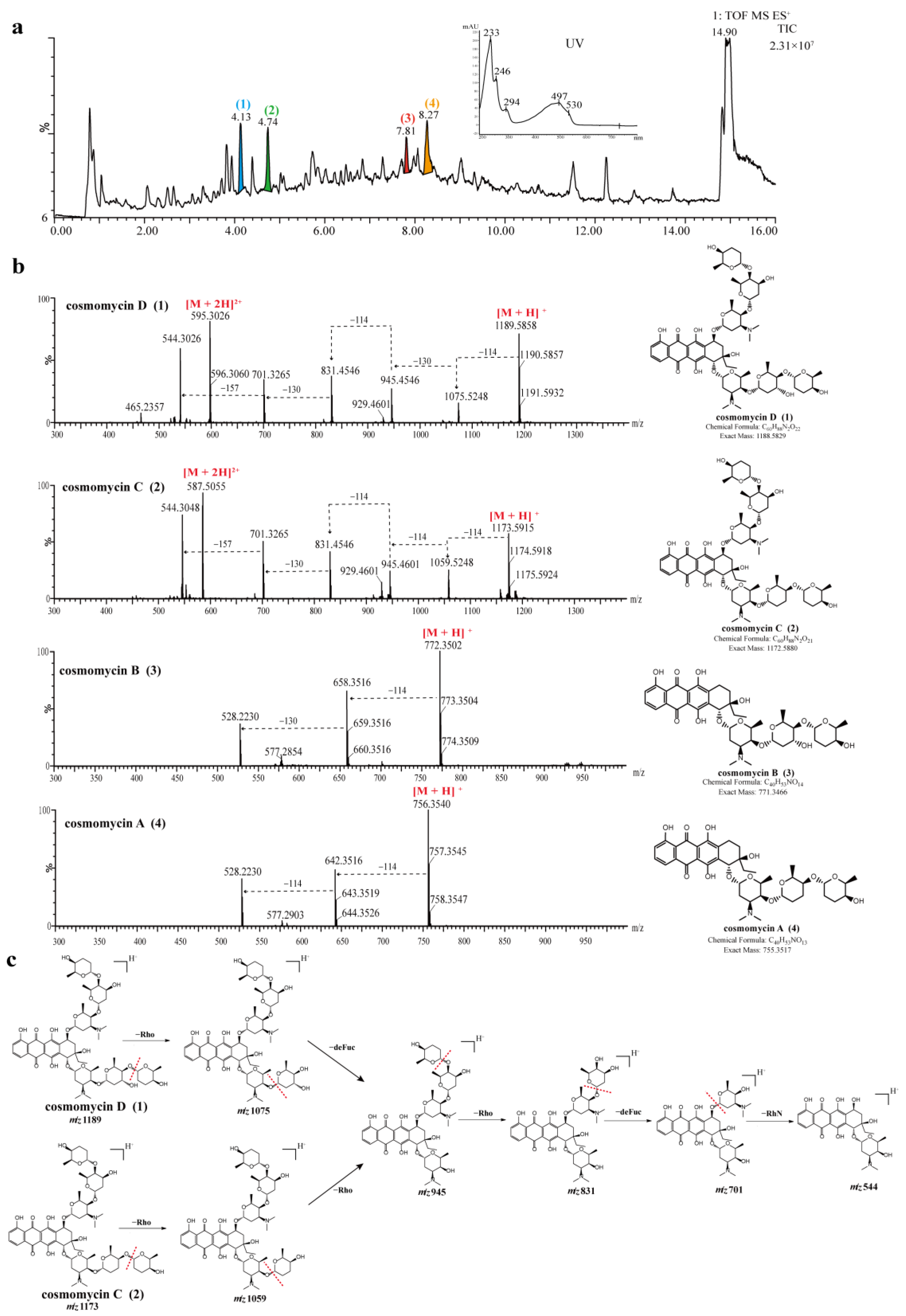
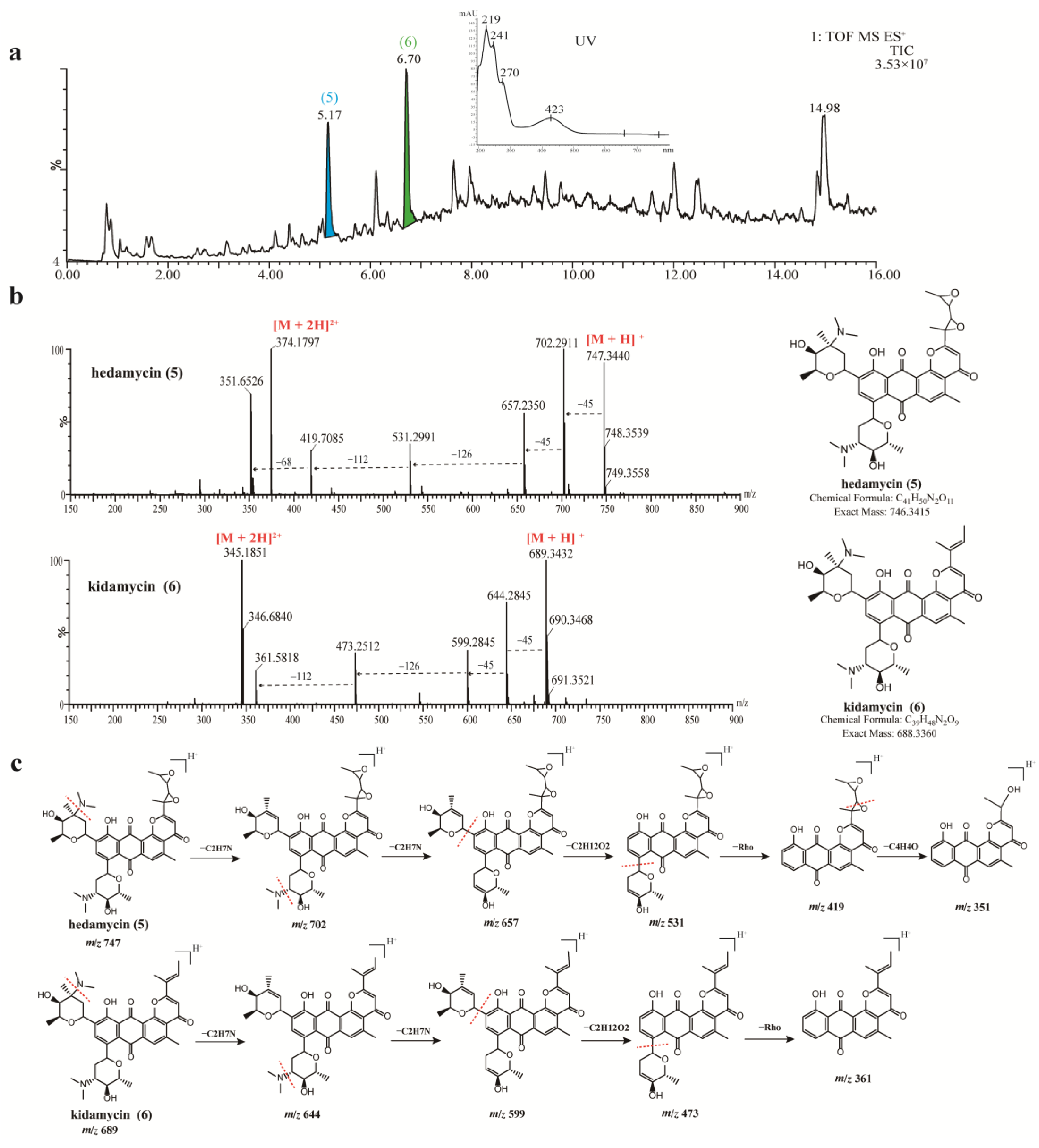
3.5. Chemical Analysis of Crude Extracts from Strains 378 and 549
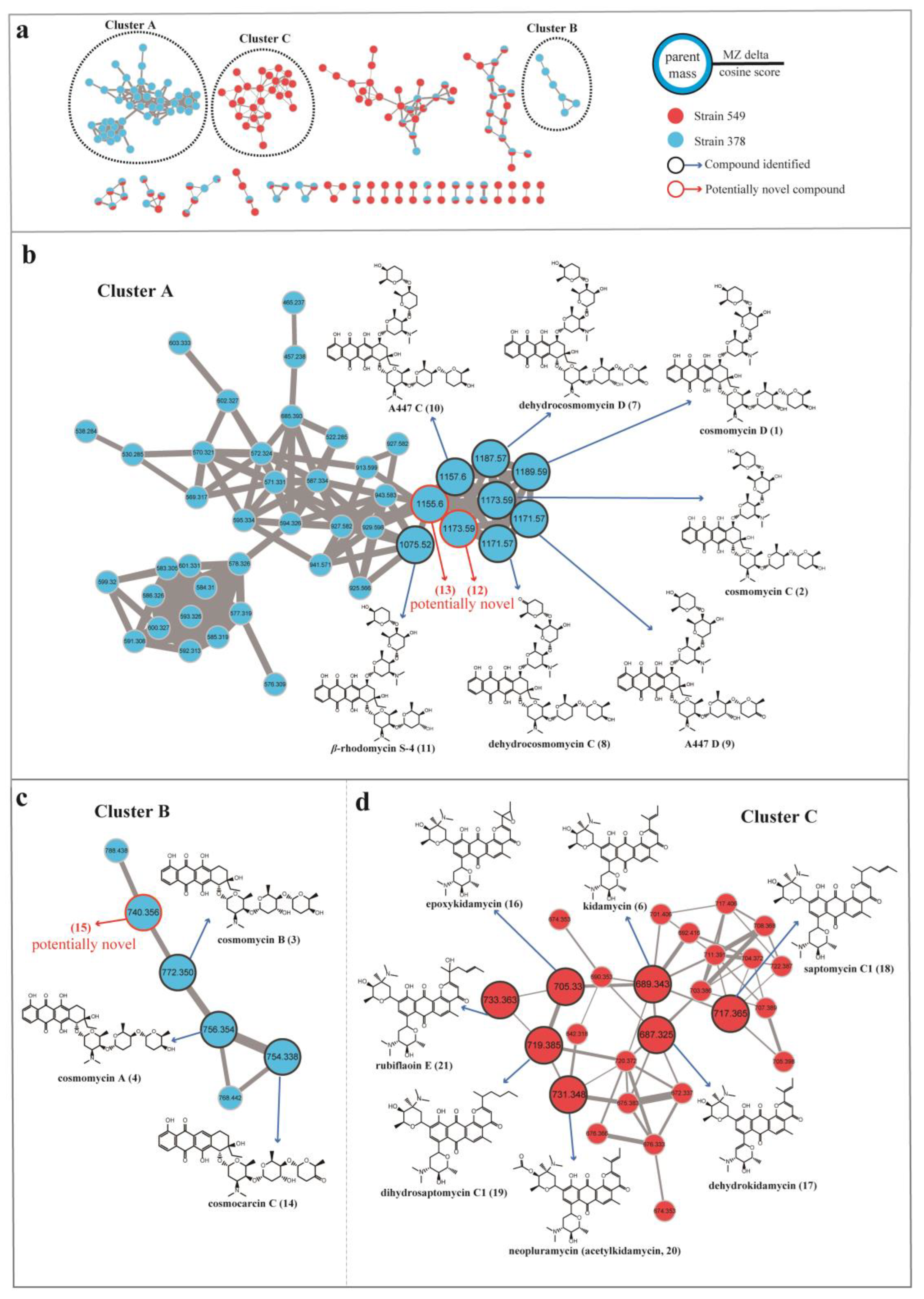
3.6. Molecular Networking Analysis of Streptomyces 378 and 549
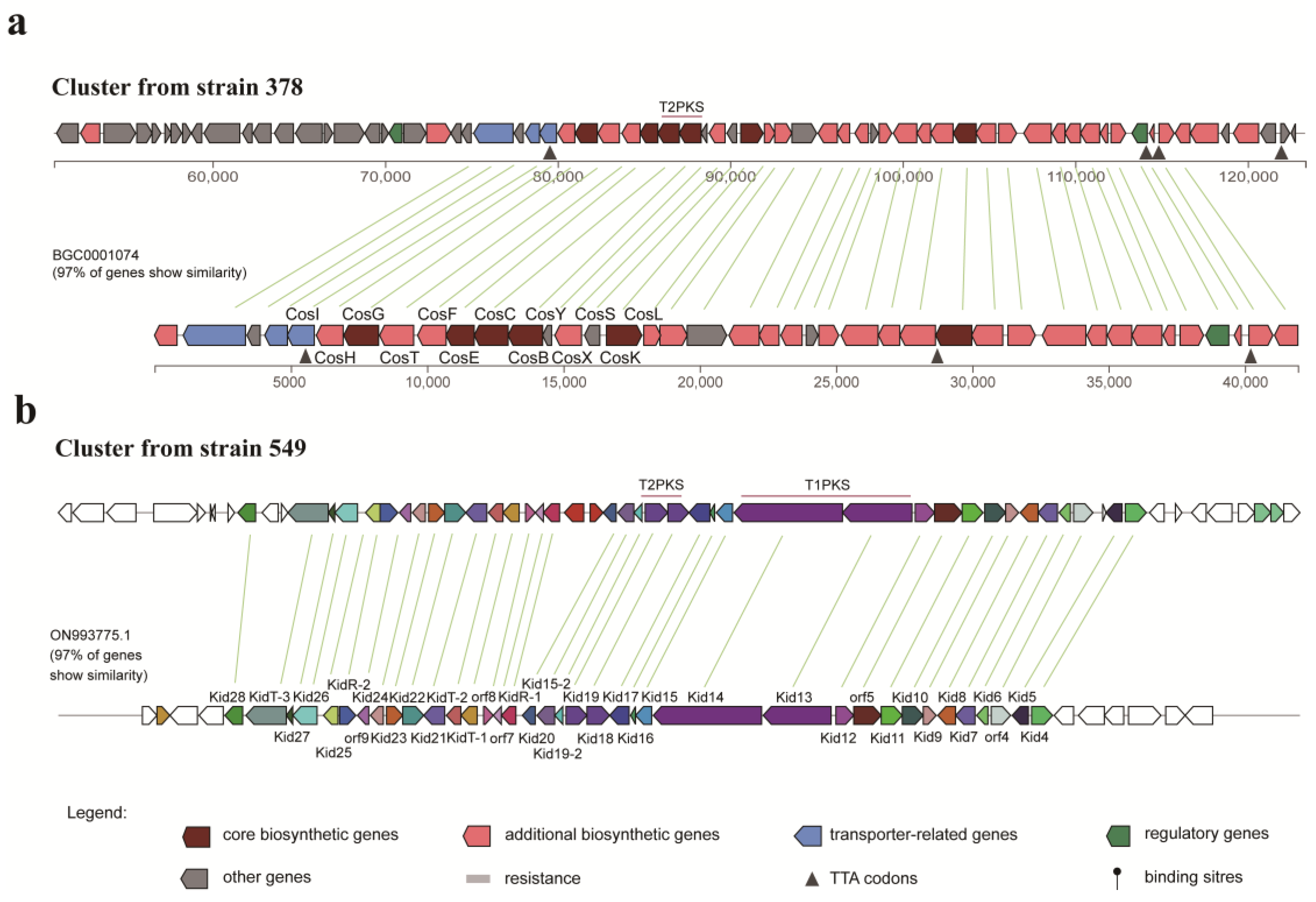
3.7. Identification of Biosynthetic Gene Clusters (BGCs) for Strains 378 and 549
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Barka, E.A.; Vatsa, P.; Sanchez, L.; Gaveau-Vaillant, N.; Jacquard, C.; Meier-Kolthoff, J.P.; Klenk, H.P.; Clément, C.; Ouhdouch, Y.; van Wezel, G.P. Taxonomy, physiology, and natural products of actinobacteria. Microbiol. Mol. Biol. Rev. 2015, 80, 1–43. [Google Scholar] [CrossRef] [PubMed]
- Genilloud, O. Actinomycetes: Still a source of novel antibiotics. Nat. Prod. Rep. 2017, 34, 1203–1232. [Google Scholar] [CrossRef] [PubMed]
- Hoel, D.; Williams, D.N. Antibiotics: Past, present, and future. Unearthing nature’s magic bullets. Postgrad. Med. 1997, 101, 114–122. [Google Scholar] [CrossRef] [PubMed]
- Clark, S.J. Magic bullets. Am. J. Med. 1992, 92, 85–87. [Google Scholar] [CrossRef] [PubMed]
- Sekurova, O.N.; Schneider, O.; Zotchev, S.B. Novel bioactive natural products from bacteria via bioprospecting, genome mining and metabolic engineering. Microb. Biotechnol. 2019, 12, 828–844. [Google Scholar] [CrossRef] [PubMed]
- Schneider, Y.K. Bacterial Natural Product Drug Discovery for New Antibiotics: Strategies for Tackling the Problem of Antibiotic Resistance by Efficient Bioprospecting. Antibiotics 2021, 10, 842. [Google Scholar] [CrossRef] [PubMed]
- Xie, F.; Pathom-Aree, W. Actinobacteria from desert: Diversity and biotechnological applications. Front. Microbiol. 2021, 12, 765531. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.B.; Ye, W.W.; Han, Y.; Deng, Z.X.; Hong, K. Natural products from mangrove actinomycetes. Mar. Drugs 2014, 12, 2590–2613. [Google Scholar] [CrossRef]
- Rangseekaew, P.; Pathom-Aree, W. Cave actinobacteria as producers of bioactive metabolites. Front. Microbiol. 2019, 10, 387. [Google Scholar] [CrossRef]
- Tortorella, E.; Tedesco, P.; Palma Esposito, F.; January, G.G.; Fani, R.; Jaspars, M.; de Pascale, D. Antibiotics from Deep-Sea Microorganisms: Current Discoveries and Perspectives. Mar. Drugs 2018, 16, 355. [Google Scholar] [CrossRef]
- Jose, P.A.; Maharshi, A.; Jha, B. Actinobacteria in natural products research: Progress and prospects. Microbiol. Res. 2021, 246, 126708. [Google Scholar] [CrossRef] [PubMed]
- Uma, G.; Babu, M.M.; Prakashy, V.S.G.; Nisha, S.J.; Citarasu, T. Nature and bioprospecting of haloalkaliphilics: A review. World J. Microbiol. Biotechnol. 2020, 36, 66. [Google Scholar] [CrossRef] [PubMed]
- Hui, M.L.; Tan, L.T.; Letchumanan, V.; He, Y.W.; Fang, C.M.; Chan, K.G.; Law, J.W.; Lee, L.H. The extremophilic actinobacteria: From microbes to medicine. Antibiotics 2021, 10, 682. [Google Scholar] [CrossRef]
- Zhao, W. Ecology of Inland Saline Lakes in China, 1st ed.; Science Press: Beijing, China, 2010; p. 3. (In Chinese) [Google Scholar]
- Javor, B. Hypersaline Environments: Microbiology and Biogeochemistry, 1st ed.; Springer: Berlin, Germany, 1989; pp. 189–204. [Google Scholar]
- Fernández, A.B.; Vera-Gargallo, B.; Sánchez-Porro, C.; Ghai, R.; Papke, R.T.; Rodriguez-Valera, F.; Ventosa, A. Comparison of prokaryotic community structure from Mediterranean and Atlantic saltern concentrator ponds by a metagenomic approach. Front. Microbiol. 2014, 5, 196. [Google Scholar] [CrossRef] [PubMed]
- Emerson, J.B.; Andrade, K.; Thomas, B.C.; Norman, A.; Allen, E.E.; Heidelberg, K.B.; Banfield, J.F. Virus-host and CRISPR dynamics in archaea-dominated hypersaline Lake Tyrrell, Victoria, Australia. Archaea 2013, 2013, 370871. [Google Scholar] [CrossRef] [PubMed]
- Oren, A. Halophilic archaea on earth and in space: Growth and survival under extreme conditions. Philos. Trans. A Math. Phys. Eng. Sci. 2014, 372, 20140194. [Google Scholar] [CrossRef] [PubMed]
- Baxter, B.K. Great Salt Lake microbiology: A historical perspective. Int. Microbiol. 2018, 21, 79–95. [Google Scholar] [CrossRef]
- Jacob, J.H.; Hussein, E.I.; Shakhatreh, M.A.K.; Cornelison, C.T. Microbial community analysis of the hypersaline water of the Dead Sea using high-throughput amplicon sequencing. Microbiologyopen 2017, 6, e00500. [Google Scholar] [CrossRef]
- Abdallah, M.B.; Karray, F.; Mhiri, N.; Mei, N.; Quéméneur, M.; Cayol, J.L.; Erauso, G.; Tholozan, J.L.; Alazard, D.; Sayadi, S. Prokaryotic diversity in a Tunisian hypersaline lake, Chott El Jerid. Extremophiles 2016, 20, 125–138. [Google Scholar] [CrossRef]
- Naghoni, A.; Emtiazi, G.; Amoozegar, M.A.; Cretoiu, M.S.; Stal, L.J.; Etemadifar, Z.; Shahzadeh Fazeli, S.A.; Bolhuis, H. Microbial diversity in the hypersaline Lake Meyghan, Iran. Sci. Rep. 2017, 7, 11522. [Google Scholar] [CrossRef]
- Cytryn, E.; Minz, D.; Oremland, R.S.; Cohen, Y. Distribution and diversity of archaea corresponding to the limnological cycle of a hypersaline stratified lake (Solar lake, Sinai, Egypt). Appl. Environ. Microbiol. 2000, 66, 3269–3276. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.C.; Dong, H.L.; Zhang, G.X.; Yu, B.S.; Leah, R.C.; Matthew, W.F. Microbial diversity in water and sediment of lake Chaka, an athalassohaline lake in northwestern China. Appl. Environ. Microbiol. 2006, 72, 3832–3845. [Google Scholar] [CrossRef] [PubMed]
- Han, R.; Zhang, X.; Liu, J.; Long, Q.; Chen, L.; Liu, D.; Zhu, D. Microbial community structure and diversity within hypersaline Keke Salt Lake environments. Can. J. Microbiol. 2017, 63, 895–908. [Google Scholar] [CrossRef] [PubMed]
- Guan, T.W.; Lin, Y.J.; Ou, M.Y.; Chen, K.B. Isolation and diversity of sediment bacteria in the hypersaline aiding lake, China. PLoS ONE 2020, 15, e0236006. [Google Scholar] [CrossRef] [PubMed]
- Sorokin, D.Y.; van Pelt, S.; Tourova, T.P.; Evtushenko, L.I. Nitriliruptor alkaliphilus gen. nov. sp. nov. a deep-lineage haloalkaliphilic actinobacterium from soda lakes capable of growth on aliphatic nitriles, and proposal of Nitriliruptoraceae fam. nov. and Nitriliruptorales ord. nov. Int. J. Syst. Evol. Microbiol. 2009, 59, 248–253. [Google Scholar] [CrossRef] [PubMed]
- Tang, S.K.; Zhi, X.Y.; Li, W.J. Jiangellaceae. In Bergey’s Manual of Systematics of Archaea and Bacteria, 2nd ed.; Trujillo, M.E., Dedysh, S., DeVos, P., Hedlund, B., Kämpfer, P., Rainey, F.A., Whitman, W.B., Eds.; John Wiley & Sons, Inc: Hoboken, NJ, USA, 2015; p. 1. [Google Scholar]
- Tang, S.K.; Zhi, X.Y.; Wang, Y.; Shi, R.; Lou, K.; Xu, L.H.; Li, W.J. Haloactinopolyspora alba gen. nov., sp. nov., a halophilic filamentous actinomycete isolated from a salt lake, with proposal of Jiangellaceae fam. nov. and Jiangellineae subord. nov. Int. J. Syst. Evol. Microbiol. 2011, 61, 194–200. [Google Scholar] [CrossRef] [PubMed]
- Tang, S.K.; Zhi, X.Y.; Wang, Y.; Wu, J.Y.; Lee, J.C.; Kim, C.J.; Lou, K.; Xu, L.H.; Li, W.J. Haloactinobacterium album gen. nov., sp. nov., a halophilic actinobacterium, and proposal of Ruaniaceae fam. nov. Int. J. Syst. Evol. Microbiol. 2010, 60, 2113–2119. [Google Scholar] [CrossRef] [PubMed]
- Waditee-Sirisattha, R.; Kageyama, H.; Takabe, T. Halophilic microorganism resources and their applications in industrial and environmental biotechnology. AIMS Microbiol. 2016, 2, 42–54. [Google Scholar] [CrossRef]
- Kim, S.H.; Shin, Y.; Lee, S.H.; Oh, K.B.; Lee, S.K.; Shin, J.; Oh, D.C. Salternamides A–D from a Halophilic Streptomyces sp. Actinobacterium. J. Nat. Prod. 2015, 78, 836–843. [Google Scholar] [CrossRef]
- Kim, S.H.; Shin, Y.; Lee, S.K.; Shin, J.; Oh, D.C. Salternamide E from a saltern-derived marine actinomycete Streptomyces sp. Nat. Prod. Sci. 2015, 21, 273–277. [Google Scholar] [CrossRef]
- Kim, M.C.; Lee, J.H.; Shin, B.; Subedi, L.; Cha, J.W.; Park, J.S.; Oh, D.C.; Kim, S.Y.; Kwon, H.C. Salinazinones A and B: Pyrrolidinyl-oxazinones from solar saltern-derived Streptomyces sp. KMF-004. Org. Lett. 2015, 17, 5024–5027. [Google Scholar] [CrossRef]
- Kim, S.H.; Ha, T.K.; Oh, W.K.; Shin, J.; Oh, D.C. Antiviral indolosesquiterpenoid Xiamycins C–E from a halophilic actinomycete. J. Nat. Prod. 2016, 79, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.X.; Zhao, L.X.; Tang, S.K.; Jiang, C.L.; Duan, Y.; Shen, B. Erythronolides H and I, new erythromycin congeners from a new halophilic actinomycete Actinopolyspora sp. YIM90600. Org. Lett. 2009, 11, 1353–1356. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.X.; Huang, S.X.; Tang, S.K.; Jiang, C.L.; Duan, Y.; Beutler, J.A.; Henrich, C.J.; McMahon, J.B.; Schmid, T.; Blees, J.S.; et al. Actinopolysporins A-C and tubercidin as a Pdcd4 stabilizer from the halophilic actinomycete Actinopolyspora erythraea YIM 90600. J. Nat. Prod. 2011, 74, 1990–1995. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Shin, D.; Kim, S.H.; Park, W.; Shin, Y.; Kim, W.K.; Lee, S.K.; Oh, K.B.; Shin, J.; Oh, D.C. Borrelidins C–E: New antibacterial macrolides from a saltern-derived halophilic Nocardiopsis sp. Mar. Drugs 2017, 15, 166. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.; Zhang, X.; Hao, H.; Li, W.; Lu, C. Nocarbenzoxazoles A-G, benzoxazoles produced by halophilic Nocardiopsis lucentensis DSM 44048. J. Nat. Prod. 2015, 78, 2123–2127. [Google Scholar] [CrossRef] [PubMed]
- Sayed, A.M.; Hassan, M.H.A.; Alhadrami, H.A.; Hassan, H.M.; Goodfellow, M.; Rateb, M.E. Extreme environments: Microbiology leading to specialized metabolites. J. Appl. Microbiol. 2020, 128, 630–657. [Google Scholar] [CrossRef] [PubMed]
- Zheng, M.; Tang, J.; Liu, J.; Zhang, F. Chinese saline lakes. Hydrobiologia 1993, 267, 23–36. [Google Scholar]
- Qiu, J. China: The third pole. Nature 2008, 454, 393–396. [Google Scholar] [CrossRef]
- Wu, J.S.; Shen, Z.X.; Zhang, X.Z. Advances in grassland ecosystem research in Northern Tibetan plateau. Agric. Sci. Technol. 2009, 10, 148–152. [Google Scholar]
- Jia, Q.; Liu, X.; Wang, H.; Liu, S.; Luo, Y.; Chen, L.; Zheng, M. Bio-ecological resources of saline lakes in Tibet and their economic prospect. Sci. Technol. Rev. 2017, 35, 19–26. (In Chinese) [Google Scholar]
- Rice, L.B. Federal funding for the study of antimicrobial resistance in nosocomial pathogens: No ESKAPE. J. Infect. Dis. 2008, 197, 1079–1081. [Google Scholar] [CrossRef] [PubMed]
- Dewapriya, P.; Khalil, Z.G.; Prasad, P.; Salim, A.A.; Cruz-Morales, P.; Marcellin, E.; Capon, R.J. Talaropeptides A–D: Structure and biosynthesis of extensively N-methylated linear peptides from an australian marine tunicate-derived Talaromyces sp. Front. Chem. 2018, 6, 394. [Google Scholar] [CrossRef] [PubMed]
- Salim, A.A.; Khalil, Z.G.; Elbanna, A.H.; Wu, T.; Capon, R.J. Methods in microbial biodiscovery. Mar. Drugs 2021, 19, 503. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.Q.; Huang, X.L.; Huang, D.Y.; Hu, X.W.; Chen, J.L. A rapid method for extracting DNA from actinomycetes by Chelex-100. Biotechnol. Bull. 2010, 2, 123–125. (In Chinese) [Google Scholar]
- Lu, Q.P.; Huang, Y.M.; Liu, S.W.; Wu, G.; Yang, Q.; Liu, L.F.; Zhang, H.T.; Qi, Y.; Wang, T.; Jiang, Z.K.; et al. Metabolomics tools assisting classic screening methods in discovering new antibiotics from mangrove actinomycetia in Leizhou Peninsula. Mar. Drugs 2021, 19, 688. [Google Scholar] [CrossRef] [PubMed]
- Yoon, S.H.; Ha, S.M.; Kwon, S.; Lim, J.; Kim, Y.; Seo, H.; Chun, J. Introducing EzBioCloud: A taxonomically united database of 16S rRNA gene sequences and whole-genome assemblies. Int. J. Syst. Evol. Microbiol. 2017, 67, 1613–1617. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar]
- Sultanbawa, Y.; Cusack, A.; Currie, M.; Davis, C. An innovative microplate assay to facilitate the detection of antimicrobial activity in plant extracts. J. Rapid. Methods. Autom. Microbiol. 2009, 17, 519–534. [Google Scholar] [CrossRef]
- Baba, T.; Ara, T.; Hasegawa, M.; Takai, Y.; Okumura, Y.; Baba, M.; Datsenko, K.A.; Tomita, M.; Wanner, B.L.; Mori, H. Construction of Escherichia coli K-12 in-frame, single-gene knockout mutants: The Keio collection. Mol. Syst. Biol. 2006, 2, 2006.0008. [Google Scholar] [CrossRef] [PubMed]
- Osterman, I.A.; Komarova, E.S.; Shiryaev, D.I.; Korniltsev, I.A.; Khven, I.M.; Lukyanov, D.A.; Tashlitsky, V.N.; Serebryakova, M.V.; Efremenkova, O.V.; Ivanenkov, Y.A.; et al. Sorting out antibiotics’ mechanisms of action: A double fluorescent protein reporter for high-throughput screening of ribosome and DNA biosynthesis inhibitors. Antimicrob. Agents Chemother. 2016, 60, 7481–7489. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Carver, J.J.; Phelan, V.V.; Sanchez, L.M.; Garg, N.; Peng, Y.; Nguyen, D.D.; Watrous, J.; Kapono, C.A.; Luzzatto-Knaan, T.; et al. Sharing and community curation of mass spectrometry data with Global Natural Products Social Molecular Networking. Nat. Biotechnol. 2016, 34, 828–837. [Google Scholar] [CrossRef] [PubMed]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef] [PubMed]
- Van Santen, J.A.; Jacob, G.; Singh, A.L.; Aniebok, V.; Balunas, M.J.; Bunsko, D.; Neto, F.C.; Castaño-Espriu, L.; Chang, C.; Clark, T.N.; et al. The Natural Products Atlas: An open access knowledge base for microbial natural products discovery. ACS Cent. Sci. 2019, 5, 1824–1833. [Google Scholar] [CrossRef] [PubMed]
- Moumbock, A.F.A.; Gao, M.; Qaseem, A.; Li, J.; Kirchner, P.A.; Ndingkokhar, B.; Bekono, B.D.; Simoben, C.V.; Babiaka, S.B.; Malange, Y.I.; et al. StreptomeDB 3.0: An updated compendium of streptomycetes natural products. Nucleic. Acids Res. 2021, 49, D600–D604. [Google Scholar] [CrossRef] [PubMed]
- Marmur, J. A procedure for the isolation of deoxyribonucleic acid from microorganisms. J. Mol. Biol. 1961, 3, 208–218. [Google Scholar] [CrossRef]
- Li, R.; Li, Y.; Kristiansen, K.; Wang, J. SOAP: Short oligonucleotide alignment program. Bioinformatics 2008, 24, 713–714. [Google Scholar] [CrossRef]
- Tatusova, T.; DiCuccio, M.; Badretdin, A.; Chetvernin, V.; Nawrocki, E.P.; Zaslavsky, L.; Lomsadze, A.; Pruitt, K.D.; Borodovsky, M.; Ostell, J. NCBI prokaryotic genome annotation pipeline. Nucleic. Acids Res. 2016, 44, 6614–6624. [Google Scholar] [CrossRef]
- Blin, K.; Shaw, S.; Augustijn, H.E.; Reitz, Z.L.; Biermann, F.; Alanjary, M.; Fetter, A.; Terlouw, B.R.; Metcalf, W.W.; Helfrich, E.J.N.; et al. antiSMASH 7.0: New and improved predictions for detection, regulation, chemical structures and visualisation. Nucleic Acids Res. 2023, 51, gkad344. [Google Scholar] [CrossRef]
- Oren, A.; Garrity, G.M. Valid publication of the names of forty-two phyla of prokaryotes. Int. J. Syst. Evol. Microbiol. 2021, 71, 005056. [Google Scholar] [CrossRef] [PubMed]
- Vandamme, P.; Peeters, C.; Seth-Smith, H.M.B.; Schmid, H.; Cnockaert, M.; Egli, A.; Goldenberger, D. Description of Pseudoclavibacter triregionum sp. nov. from human blood and Pseudoclavibacter albus comb. nov., and revised classification of the genus Pseudoclavibacter: Proposal of Caespitibacter gen. nov., with Caespitibacter soli comb. nov. and Caespitibacter caeni comb. nov. Antonie Leeuwenhoek 2022, 115, 461–472. [Google Scholar] [PubMed]
- Khalil, Z.G.; Salim, A.A.; Capon, R.J. Microbioreactor techniques for the production and spectroscopic characterization of microbial peptides. Methods Mol. Biol. 2020, 2103, 303–322. [Google Scholar] [PubMed]
- Ivanenkov, Y.A.; Komarova, E.S.; Osterman, I.A.; Sergiev, P.V.; Yamidanov, R.S.; Deineka, E.V.; Terent’ev, V.A.; Fil’kov, G.I.; Sofronova, A.A.; Mazhuga, A.G.; et al. N-pyridyl-substituted carboxypiperidine amides: A new class of prokaryote translation inhibitors. Pharm. Chem. J. 2019, 53, 225–229. [Google Scholar] [CrossRef]
- Osterman, I.A.; Wieland, M.; Maviza, T.P.; Lashkevich, K.A.; Lukianov, D.A.; Komarova, E.S.; Zakalyukina, Y.V.; Buschauer, R.; Shiriaev, D.I.; Leyn, S.A.; et al. Tetracenomycin X inhibits translation by binding within the ribosomal exit tunnel. Nat. Chem. Biol. 2020, 16, 1071–1077. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Li, F.; Lu, Q.; Wu, G.; Jiang, Z.; Liu, S.; Habden, X.; Razumova, E.A.; Osterman, I.A.; Sergiev, P.V.; et al. Diversity, novelty, antimicrobial activity, and new antibiotics of cultivable endophytic actinobacteria isolated from psammophytes collected from Taklamakan Desert. J. Pharm. Anal. 2021, 11, 241–250. [Google Scholar] [CrossRef] [PubMed]
- Axenov-Gribanov, D.V.; Voytsekhovskaya, I.V.; Tokovenko, B.T.; Protasov, E.S.; Gamaiunov, S.V.; Rebets, Y.V.; Luzhetskyy, A.N.; Timofeyev, M.A. Actinobacteria isolated from an underground lake and moonmilk speleothem from the biggest conglomeratic karstic cave in siberia as sources of novel biologically active compounds. PLoS ONE 2016, 11, e0149216. [Google Scholar] [CrossRef] [PubMed]
- Ando, T.; Hirayama, K.; Takahashi, R.; Horino, I.; Etoh, Y.; Morioka, H.; Shibai, H.; Murai, A. The structures of anthracycline antibiotics, cosmomycins A and B. Agric. Biol. Chem. 1985, 49, 1207–1209. [Google Scholar]
- Ando, T.; Hirayama, K.; Takahashi, R.; Horino, I.; Etoh, Y.; Morioka, H.; Shibai, H.; Murai, A. Cosmomycin D, a new anthracycline antibiotic. Agric. Biol. Chem. 1985, 49, 259–262. [Google Scholar]
- Hirayama, K.; Akashi, S.; Ando, T.; Horino, I.; Etoh, Y.; Morioka, H.; Shibai, H.; Murai, A. Field desorption tandem mass spectrometry of anthracycline antibiotics, cosmomycin A, B, A’, B’, C and D. Biomed. Environ. Mass Spectrom. 1987, 14, 305–312. [Google Scholar] [CrossRef]
- Kelso, C.; Rojas, J.D.; Furlan, R.L.; Padilla, G.; Beck, J.L. Characterisation of anthracyclines from a cosmomycin D-producing species of Streptomyces by collisionally-activated dissociation and ion mobility mass spectrometry. Eur. J. Mass Spectrom. 2009, 15, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Chen, Y.L. Structural studies on rhodilunancins A and B. J. Antibiot. 1986, 39, 430–436. [Google Scholar] [CrossRef] [PubMed]
- Morioka, H.; Etoh, Y.; Horino, I.; Takezawa, M.; Ando, T.; Hirayama, K.; Kano, H.; Shibai, H. Production and isolation of cosmomycins A, B, C and D: New differentiation inducers of Friend cell F5-5. Agric. Biol. Chem. 1985, 49, 1951–1958. [Google Scholar]
- Waluyo, D.; Prabandari, E.E.; Pramisandi, A.; Hidayati, D.N.; Chrisnayanti, E.; Puspitasari, D.J.; Dewi, D.; Suryani; Kristiningrum; Oktaviani, A.N.; et al. Exploring natural microbial resources for the discovery of anti-malarial compounds. Parasitol. Int. 2021, 85, 102432. [Google Scholar] [CrossRef] [PubMed]
- Séquin, U.; Bedford, C.T.; Chung, S.K.; Scott, A.I. The structure of the antibiotic hedamycin I. Chemical, physical and spectral properties. Helv. Chim. Acta 1977, 60, 896–906. [Google Scholar] [CrossRef]
- Séquin, U. The structure of the antibiotic hedamycin-II: Comparison of hedamycin and kidamycin. Tetrahedron 1978, 34, 761–767. [Google Scholar] [CrossRef]
- Kanda, N. A new antitumor antibiotic, kidamycin. I. Isolation, purification and properties of kidamycin. J. Antibiot. 1971, 24, 599–606. [Google Scholar] [CrossRef] [PubMed]
- Kanda, N.; Kono, M.; Asano, K. A new antitumor antibiotic, kidamycin. II. Experimental treatment of cancer with kidamycin. J. Antibiot. 1972, 25, 553–556. [Google Scholar] [CrossRef]
- Kanda, N. A new antitumor antibiotic, kidamycin. III. Preparation and properties of acetyl kidamycin. J. Antibiot. 1972, 25, 557–560. [Google Scholar] [CrossRef]
- Ui, H.; Ishiyama, A.; Sekiguchi, H.; Namatame, M.; Nishihara, A.; Takahashi, Y.; Shiomi, K.; Otoguro, K.; Omura, S. Selective and potent in vitro antimalarial activities found in four microbial metabolites. J. Antibiot. 2007, 60, 220–222. [Google Scholar] [CrossRef]
- Hamao, U.; Tomio, T.; Masa, H.; Hiroshi, N.; Tsutomu, S.; Takeshi, U.; Masaya, I. Anthracycline Derivatives and Their Use as Medicaments. Patent EP167935A2, 15 January 1986. [Google Scholar]
- Werner, A.; Gerd, B.H.; Wolfram, F.H.; Gerhard, H.; Peter, K.H. Microbiological Production of Anthracycline Derivatives. Patent DE3415544A1, 21 November 1985. [Google Scholar]
- Shimosaka, A.; Hayakawa, Y.; Nakagawa, M.; Furihata, K.; Seto, H.; Otake, N. Isolation of new anthracycline antibiotics, A447 C and D. J. Antibiot. 1987, 40, 116–121. [Google Scholar] [CrossRef] [PubMed]
- Hans, B.; Hartwig, G. β-Rhodomycin. Tetrahedron Lett. 1975, 11, 831–834. [Google Scholar]
- Tomoko, T.; Misako, T.; Yuzuru, E.; Kazunari, H.; Hiroshiro, S. Manufacture of Antitumor Cosmocarsins. Japan Patent JP61212595A, 20 September 1986. [Google Scholar]
- Byrne, K.M.; Gonda, S.K.; Hilton, B.D. Largomycin FII chromophore component 4, a new pluramycin antibiotic. J. Antibiot. 1985, 38, 1040–1049. [Google Scholar] [CrossRef] [PubMed]
- Abe, N.; Enoki, N.; Nakakita, Y.; Uchida, H.; Nakamura, T.; Munekata, M. Novel antitumor antibiotics, saptomycins. II. Isolation, physico-chemical properties and structure elucidation. J. Antibiot. 1993, 46, 1536–1549. [Google Scholar] [CrossRef] [PubMed]
- O’Keefe, B.M.; Mans, D.M.; Kaelin, D.E., Jr.; Martin, S.F. Studies toward the syntheses of pluramycin natural products. The first total synthesis of isokidamycin. Tetrahedron 2011, 67, 6524–6538. [Google Scholar] [CrossRef]
- Kondo, S.; Miyamoto, M.; Naganawa, H.; Takeuchi, T.; Umezawa, H. Structures of pluramycin A and neopluramycin. J. Antibiot. 1977, 30, 1143–1145. [Google Scholar] [CrossRef]
- Nadig, H.; Sèquin, U. Isolation and structure elucidation of some components of the antitumor antibiotic mixture ‘Rubiflavin’. Helv. Chim. Acta 1987, 70, 1217–1228. [Google Scholar] [CrossRef]
- Hu, A.; Yao, T.; Jiao, N.; Liu, Y.; Yang, Z.; Liu, X. Community structures of ammonia-oxidising archaea and bacteria in high-altitude lakes on the Tibetan Plateau. Freshw. Biol. 2010, 55, 2375–2390. [Google Scholar] [CrossRef]
- Liu, Y.; Priscu, J.C.; Yao, T.; Vick-Majors, T.J.; Michaud, A.B.; Jiao, N.; Hou, J.; Tian, L.; Hu, A.; Chen, Z.Q. A comparison of pelagic, littoral, and riverine bacterial assemblages in Lake Bangongco, Tibetan Plateau. FEMS Microbiol. Ecol. 2014, 89, 211–221. [Google Scholar] [CrossRef]
- Zeng, F.; Zhu, Y.; Zhang, D.; Zhao, Z.; Li, Q.; Ma, P.; Zhang, G.; Wang, Y.; Wu, S.; Guo, S.; et al. Metagenomic analysis of the soil microbial composition and salt tolerance mechanism in Yuncheng Salt Lake, Shanxi Province. Front. Microbiol. 2022, 13, 1004556. [Google Scholar] [CrossRef]
- Zhang, G.J.; He, J.Q. Populations of moderately-halophilic actinomycetes in the Nam Co salt lake. Arid Zone Res. 2013, 30, 856–859. (In Chinese) [Google Scholar]
- Guan, T.W. Actinomycetes of China Salt Lakes, 1st ed.; China Light Industry Press: Beijing, China, 2017; pp. 54–58. (In Chinese) [Google Scholar]
- Pan, W.; Lin, J.; Wang, X.; Guo, Y.; Chu, Y.; Liu, C. Isolation, Identification and Antimicrobial Activity Determination of Actinobacteria from the Lakes in Tibet. Biotechnol. Bull. 2020, 36, 97–103. (In Chinese) [Google Scholar]
- Xiong, J.; Liu, Y.; Lin, X.; Zhang, H.; Zeng, J.; Hou, J.; Yang, Y.; Yao, T.; Knight, R.; Chu, H. Geographic distance and pH drive bacterial distribution in alkaline lake sediments across Tibetan Plateau. Environ. Microbiol. 2012, 14, 2457–2466. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Z.P.; Liu, Y.; Miao, L.L.; Wang, F.; Chu, L.M.; Wang, J.L.; Liu, Z.P. Prokaryotic community structure driven by salinity and ionic concentrations in plateau lakes of the Tibetan Plateau. Appl. Environ. Microbiol. 2016, 82, 1846–1858. [Google Scholar] [CrossRef] [PubMed]
- Yue, X.J.; Yu, L.Y.; Li, Q.P.; Wei, Y.Z.; Guan, Y.; Zhang, Y.Q. Study of methods to isolate viable but non-culturable microorganisms from natural environment. Microbiology 2006, 33, 77–81. (In Chinese) [Google Scholar]
- Zhang, Y.Q.; Liu, H.Y.; Chen, J.; Yuan, L.J.; Sun, W.; Zhang, L.X.; Zhang, Y.Q.; Yu, L.Y.; Li, W.J. Diversity of culturable actinobacteria from Qinghai-Tibet plateau, China. Antonie Leeuwenhoek 2010, 98, 213–223. [Google Scholar] [CrossRef] [PubMed]
- Bússolo, T.B.; Souza, C.M.D.; Souza, N.A.A.D.; Furlaneto, M.C.; Bona, E.; Furlaneto-Maia, L. Soybean flour as a substrate to obtain Enterococcus durans bacteriocins. Ciência Agrotecnol. 2022, 46, e008022. [Google Scholar] [CrossRef]
- Furlan, R.L.; Watt, S.J.; Garrido, L.M.; Amarante-Mendes, G.P.; Nur-e-alam, M.; Rohr, J.; Braña, A.; Mendez, C.; Salas, J.A.; Sheil, M.M.; et al. DNA-binding properties of cosmomycin D, an anthracycline with two trisaccharide chains. J. Antibiot. 2004, 57, 647–654. [Google Scholar] [CrossRef]
- Hansen, M.; Yun, S.; Hurley, L. Hedamycin intercalates the DNA helix and, through carbohydrate-mediated recognition in the minor groove, directs N7-alkylation of guanine in the major groove in a sequence-specific manner. Chem. Biol. 1995, 2, 229–240. [Google Scholar] [CrossRef]
- Pavlopoulos, S.; Bicknell, W.; Wickham, G.; Craik, D.J. Characterization of the sequential non-covalent and covalent interactions of the antitumour antibiotic hedamycin with double stranded DNA by NMR spectroscopy. J. Mol. Recognit. 1999, 12, 346–354. [Google Scholar] [CrossRef]
- Parker, K.A.; Koh, Y. Methodology for the regiospecific synthesis of bis C-Aryl glycosides. Models for Kidamycins. J. Am. Chem. Soc. 1994, 116, 11149–11150. [Google Scholar] [CrossRef]
- Hulst, M.B.; Grocholski, T.; Neefjes, J.J.C.; van Wezel, G.P.; Metsä-Ketelä, M. Anthracyclines: Biosynthesis, engineering and clinical applications. Nat. Prod. Rep. 2022, 39, 814–841. [Google Scholar] [CrossRef] [PubMed]
- Tenconi, E.; Rigali, S. Self-resistance mechanisms to DNA-damaging antitumor antibiotics in actinobacteria. Curr. Opin. Microbiol. 2018, 45, 100–108. [Google Scholar] [CrossRef] [PubMed]
- Tenconi, E.; Traxler, M.; Tellatin, D.; van Wezel, G.P.; Rigali, S. Prodiginines postpone the onset of sporulation in Streptomyces coelicolor. Antibiotics 2020, 9, 847. [Google Scholar] [CrossRef] [PubMed]
- Tuli, H.S.; Chaudhary, P.; Beniwal, V.; Sharma, A.K. Microbial pigments as natural color sources: Current trends and future perspectives. J. Food. Sci. Technol. 2015, 52, 4669–4678. [Google Scholar] [CrossRef] [PubMed]
- Celedón, R.S.; Díaz, L.B. Natural pigments of bacterial origin and their possible biomedical applications. Microorganisms 2021, 9, 739. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, S.-W.; Zhai, X.-X.; Liu, D.; Liu, Y.-Y.; Sui, L.-Y.; Luo, K.-K.; Yang, Q.; Li, F.-N.; Nikandrova, A.A.; Imamutdinova, A.N.; et al. Bioprospecting of Actinobacterial Diversity and Antibacterial Secondary Metabolites from the Sediments of Four Saline Lakes on the Northern Tibetan Plateau. Microorganisms 2023, 11, 2475. https://doi.org/10.3390/microorganisms11102475
Liu S-W, Zhai X-X, Liu D, Liu Y-Y, Sui L-Y, Luo K-K, Yang Q, Li F-N, Nikandrova AA, Imamutdinova AN, et al. Bioprospecting of Actinobacterial Diversity and Antibacterial Secondary Metabolites from the Sediments of Four Saline Lakes on the Northern Tibetan Plateau. Microorganisms. 2023; 11(10):2475. https://doi.org/10.3390/microorganisms11102475
Chicago/Turabian StyleLiu, Shao-Wei, Xiao-Xu Zhai, Di Liu, Yu-Yu Liu, Li-Ying Sui, Ke-Ke Luo, Qin Yang, Fei-Na Li, Arina A. Nikandrova, Arina N. Imamutdinova, and et al. 2023. "Bioprospecting of Actinobacterial Diversity and Antibacterial Secondary Metabolites from the Sediments of Four Saline Lakes on the Northern Tibetan Plateau" Microorganisms 11, no. 10: 2475. https://doi.org/10.3390/microorganisms11102475