

Novel Insights on the Bacterial and Archaeal Diversity of the Panarea Shallow-Water Hydrothermal Vent Field
 , , , , , ,
, , , , , , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling Sites and Sampling Strategy
2.2. DNA Extraction and Purification and Downstream Molecular Analyses
2.3. Bioinformatic and Statistical Analyses
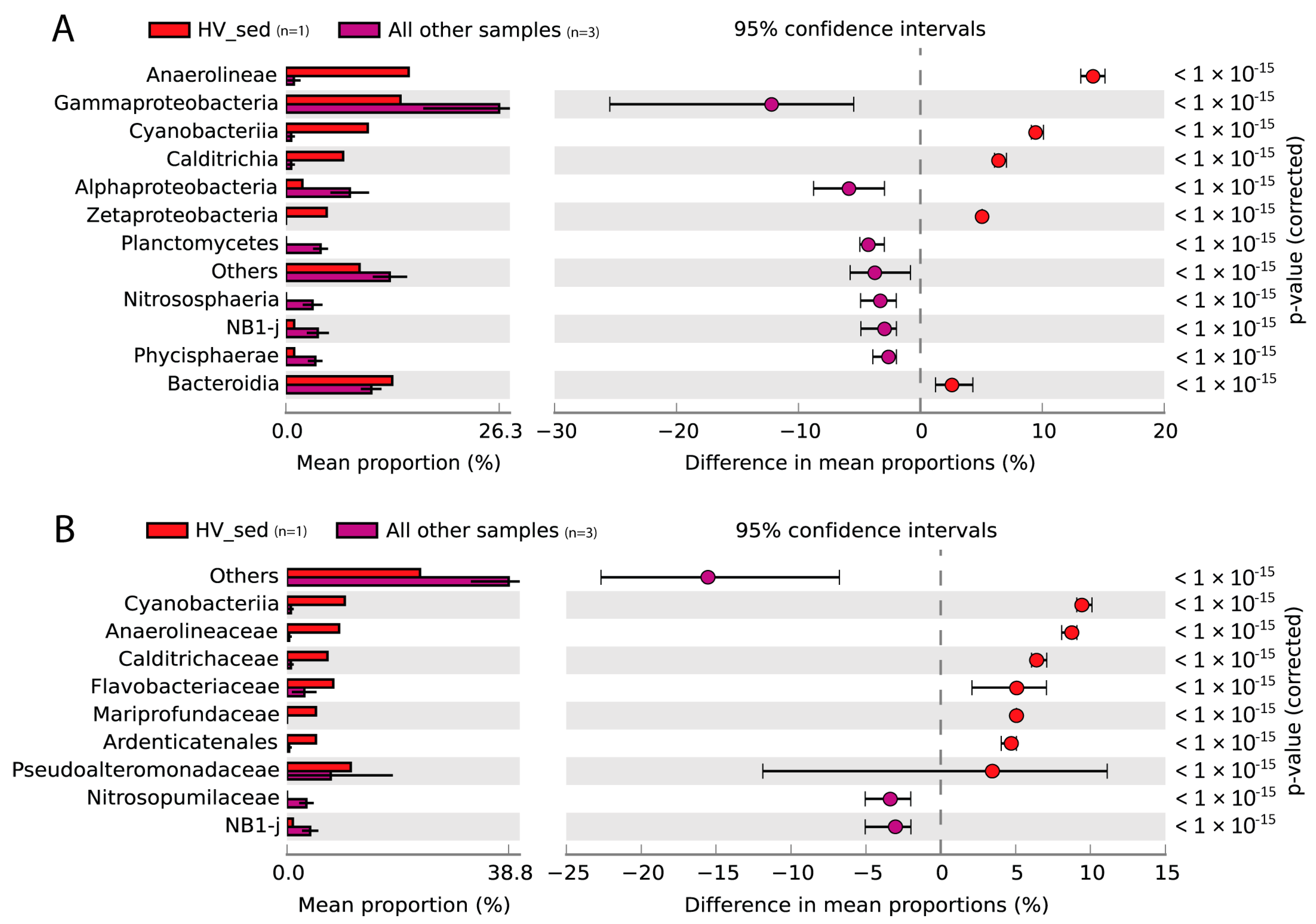
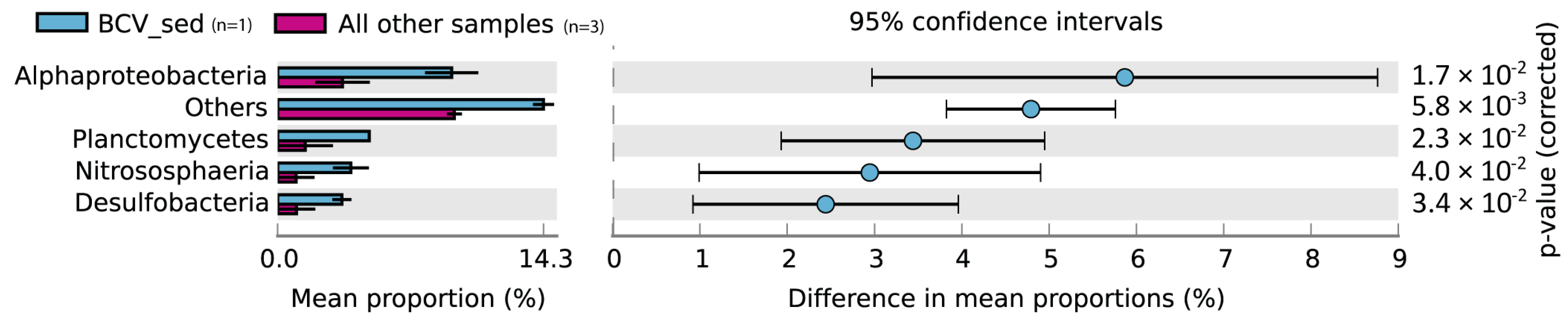
3. Results and Discussion
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ristova, P.; Pichler, T.; Friedrich, M.W.; Bühring, S.I. Bacterial diversity and biogeochemistry of two marine shallow-water hydrothermal systems off dominica (Lesser Antilles). Front. Microbiol. 2017, 8, 2400. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Tang, K.; Zhang, L.; Zhao, Z.; Xie, X.; Chen, C.T.A.; Wang, D.; Jiao, N.; Zhang, Y. Coupled carbon, sulfur, and nitrogen cycles mediated by microorganisms in the water column of a shallow-water hydrothermal ecosystem. Front. Microbiol. 2018, 9, 2718. [Google Scholar] [CrossRef] [PubMed]
- Tarasov, V.G.; Gebruk, A.V.; Mironov, A.N.; Moskalev, L.I. Deep-sea and shallow-water hydrothermal vent communities: Two different phenomena? Chem. Geol. 2005, 224, 5–39. [Google Scholar] [CrossRef]
- Dick, G.J. The microbiomes of deep-sea hydrothermal vents: Distributed globally, shaped locally. Nat. Rev. Microbiol. 2019, 17, 271–283. [Google Scholar] [CrossRef]
- Laffoley, D.; Baxter, J.M.; Thevenon, F.; Oliver, J. The Significance and Management of Natural Carbon Stores in the Open Ocean; Full Report; IUCN: Gland, Switzerland, 2014; p. 124. [Google Scholar]
- Rastelli, E.; Corinaldesi, C.; Dell’Anno, A.; Tangherlini, M.; Martorelli, E.; Ingrassia, M.; Chiocci, F.L.; Lo Martire, M.; Danovaro, R. High potential for temperate viruses to drive carbon cycling in chemoautotrophy-dominated shallow-water hydrothermal vents. Environ. Microbiol. 2017, 19, 4432–4446. [Google Scholar] [CrossRef]
- Bellec, L.; Cambon-Bonavita, M.A.; Durand, L.; Aube, J.; Gayet, N.; Sandulli, R.; Brandily, C.; Zeppilli, D. Microbial communities of the shallow-water hydrothermal vent near Naples, Italy, and chemosynthetic symbionts associated with a free-living marine nematode. Front. Microbiol. 2020, 11, 2023. [Google Scholar] [CrossRef]
- Arcadi, E.; Rastelli, E.; Tangherlini, M.; Rizzo, C.; Mancuso, M.; Sanfilippo, M.; Esposito, V.; Andaloro, F.; Romeo, T. Shallow-Water Hydrothermal Vents as Natural Accelerators of Bacterial Antibiotic Resistance in Marine Coastal Areas. Microorganisms 2022, 10, 479. [Google Scholar] [CrossRef]
- Sievert, S.M.; Bühring, S.I.; Gulmann, L.K.; Hinrichs, K.U.; Pop Ristova, P.; Gomez-Saez, G.V. Fluid flow stimulates chemoautotrophy in hydrothermally influenced coastal sediments. Commun. Earth Environ. 2022, 3, 96. [Google Scholar] [CrossRef]
- Tarasov, V.G. Effects of shallow-water hydrothermal venting on biological communities of coastal marine ecosystems of the Western Pacific. Adv. Mar. Biol. 2006, 50, 267–421. [Google Scholar]
- Hirayama, H.; Sunamura, M.; Takai, K.; Nunoura, T.; Noguchi, T.; Oida, H.; Furushima, Y.; Yamamoto, H.; Oomori, T.; Horikoshi, K. Culture-dependent and-independent characterization of microbial communities associated with a shallow submarine hydrothermal system occurring within a coral reef off Taketomi Island, Japan. Appl. Environ. Microbiol. 2007, 73, 7642–7656. [Google Scholar] [CrossRef]
- Maugeri, T.L.; Lentini, V.; Gugliandolo, C.; Cousin, S.; Stackebrandt, E. Microbial diversity at a hot, shallow-sea hydrothermal vent in the southern Tyrrhenian Sea (Italy). Geomicrobiol. J. 2010, 27, 380–390. [Google Scholar] [CrossRef]
- Maugeri, T.L.; Gugliandolo, C.; Lentini, V. Diversity of prokaryotes at a shallow submarine vent of Panarea Island (Italy) by high-throughput sequencing. AAPP Atti Accad. 2013, 91. [Google Scholar] [CrossRef]
- Wang, L.; Cheung, M.K.; Kwan, H.S.; Hwang, J.S.; Wong, C.K. Microbial diversity in shallow-water hydrothermal sediments of Kueishan Island, Taiwan as revealed by pyrosequencing. J. Basic Microbiol. 2015, 55, 1308–1318. [Google Scholar] [CrossRef] [PubMed]
- Rajasabapathy, R.; Mohandass, C.; Bettencourt, R.; Colaço, A.; Goulart, J.; Meena, R.M. Bacterial diversity at a shallow-water hydrothermal vent (Espalamaca) in Azores Island. Curr. Sci. 2018, 115, 2110–2121. [Google Scholar] [CrossRef]
- Gugliandolo, C.; Maugeri, T.L. Phylogenetic diversity of archaea in shallow hydrothermal vents of Eolian Islands, Italy. Diversity 2019, 11, 156. [Google Scholar] [CrossRef]
- Waite, D.W.; Vanwonterghem, I.; Rinke, C.; Parks, D.H.; Zhang, Y.; Takai, K.; Sievert, S.M.; Simon, J.; Campbell, B.J.; Hanson, T.E.; et al. Comparative genomic analysis of the class Epsilonproteobacteria and proposed reclassification to Epsilonbacteraeota (phyl. nov.). Front. Microbiol. 2017, 8, 682. [Google Scholar] [CrossRef]
- Chen, X.; Tang, K.; Zhang, M.; Liu, S.; Chen, M.; Zhan, P.; Fan, W.; Chen, C.T.; Zhang, Y. Genome-centric insight into metabolically active microbial population in shallow-sea hydrothermal vents. Microbiome 2022, 10, 170. [Google Scholar] [CrossRef]
- Gugliandolo, C.; Lentini, V.; Bunk, B.; Overmann, J.; Italiano, F.; Maugeri, T.L. Changes in prokaryotic community composition accompanying a pronounced temperature shift of a shallow marine thermal brine pool (Panarea Island, Italy). Extremophiles 2015, 19, 547–559. [Google Scholar] [CrossRef]
- Price, R.; Boyd, E.S.; Hoehler, T.M.; Wehrmann, L.M.; Bogason, E.; Valtýsson, H.Þ.; Örlygsson, J.; Gautason, B.; Amend, J.P. Alkaline vents and steep Na+ gradients from ridge-flank basalts—Implications for the origin and evolution of life. Geology 2017, 45, 1135–1138. [Google Scholar] [CrossRef]
- Sciutteri, V.; Smedile, F.; Vizzini, S.; Mazzola, A.; Vetriani, C. Microbial biofilms along a geochemical gradient at the shallow-water hydrothermal system of Vulcano island, Mediterranean Sea. Front. Microbiol. 2022, 13, 840205. [Google Scholar] [CrossRef] [PubMed]
- Patwardhan, S.; Smedile, F.; Giovannelli, D.; Vetriani, C. Metaproteogenomic profiling of chemosynthetic microbial biofilms reveals metabolic flexibility during colonization of a shallow-water gas vent. Front. Microbiol. 2021, 12, 638300. [Google Scholar] [CrossRef] [PubMed]
- Lentini, V.; Gugliandolo, C.; Bunk, B.; Overmann, J.; Maugeri, T.L. Diversity of prokaryotic community at a shallow marine hydrothermal site elucidated by Illumina sequencing technology. Curr. Microbiol. 2014, 69, 457–466. [Google Scholar] [CrossRef] [PubMed]
- Italiano, F. Hydrothermal fluids vented at shallow depths at the Aeolian islands: Relationships with volcanic and geothermal systems. Res. Shallow Mar. Fresh Water Syst. 2009, 22, 55–60. [Google Scholar]
- Aliani, S.; Bortoluzzi, G.; Caramanna, G.; Raffa, F. Seawater dynamics and environmental settings after November 2002 gas eruption off Bottaro (Panarea, Aeolian Islands, Mediterranean Sea). Cont. Shelf Res. 2010, 30, 1338–1348. [Google Scholar] [CrossRef]
- Stefanni, S.; Stanković, D.; Borme, D.; de Olazabal, A.; Juretić, T.; Pallavicini, A.; Tirelli, V. Multi-marker metabarcoding approach to study mesozooplankton at basin scale. Sci. Rep. 2018, 8, 12085. [Google Scholar] [CrossRef]
- Parada, A.E.; Needham, D.M.; Fuhrman, J.A. Every base matters: Assessing small subunit rRNA primers for marine microbiomes with mock communities, time series and global field samples. Environ. Microbiol. 2016, 18, 1403–1414. [Google Scholar] [CrossRef]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Tangherlini, M.; Corinaldesi, C.; Ape, F.; Greco, S.; Romeo, T.; Andaloro, F.; Danovaro, R. Ocean acidification induces changes in virus–host relationships in Mediterranean benthic ecosystems. Microorganisms 2021, 9, 769. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2012, 41, D590–D596. [Google Scholar] [CrossRef]
- Parks, D.H.; Tyson, G.W.; Hugenholtz, P.; Beiko, R.G. STAMP: Statistical analysis of taxonomic and functional profiles. Bioinformatics 2014, 30, 3123–3124. [Google Scholar] [CrossRef]
- Luna, G.M.; Corinaldesi, C.; Rastelli, E.; Danovaro, R. Patterns and drivers of bacterial α-and β-diversity across vertical profiles from surface to subsurface sediments. Environ. Microbiol. Rep. 2013, 5, 731–739. [Google Scholar] [CrossRef] [PubMed]
- Tangherlini, M.; Corinaldesi, C.; Rastelli, E.; Musco, L.; Armiento, G.; Danovaro, R.; Dell’Anno, A. Chemical contamination can promote turnover diversity of benthic prokaryotic assemblages: The case study of the Bagnoli-Coroglio bay (southern Tyrrhenian Sea). Mar. Environ. Res. 2020, 160, 105040. [Google Scholar] [CrossRef]
- Rastelli, E.; Corinaldesi, C.; Dell’Anno, A.; Tangherlini, M.; Lo Martire, M.; Nishizawa, M.; Nomaki, H.; Nunoura, T.; Danovaro, R. Drivers of bacterial α-and β-diversity patterns and functioning in subsurface hadal sediments. Front. Microbiol. 2019, 10, 2609. [Google Scholar] [CrossRef]
- Clarke, K.R.; Gorley, R.N. PRIMER v6: User Manual/Tutorial; PRIMER-E: Plymouth, UK, 2006; 192 pp. [Google Scholar]
- McArdle, B.H.; Anderson, M.J. Fitting multivariate models to community data: A comment on distance-based redundancy analysis. Ecology 2001, 82, 290–297. [Google Scholar] [CrossRef]
- Tang, K.; Zhang, Y.; Lin, D.; Han, Y.; Chen, C.T.A.; Wang, D.; Lin, Y.S.; Sun, J.; Zheng, Q.; Jiao, N. Cultivation-independent and cultivation-dependent analysis of microbes in the shallow-sea hydrothermal system off Kueishantao Island, Taiwan: Unmasking heterotrophic bacterial diversity and functional capacity. Front. Microbiol. 2018, 9, 279. [Google Scholar] [CrossRef] [PubMed]
- Paerl, H.W. Marine plankton. In Ecology of Cyanobacteria II: Their Diversity in Space and Time; Springer: Dordrecht, The Netherlands, 2012; pp. 127–153. [Google Scholar]
- Giovannoni, S.J. SAR11 bacteria: The most abundant plankton in the oceans. Ann. Rev. Mar. Sci. 2017, 9, 231–255. [Google Scholar] [CrossRef] [PubMed]
- Roda-Garcia, J.J.; Haro-Moreno, J.M.; Huschet, L.A.; Rodriguez-Valera, F.; López-Pérez, M. Phylogenomics of SAR116 Clade Reveals Two Subclades with Different Evolutionary Trajectories and an Important Role in the Ocean Sulfur Cycle. Msystems 2021, 6, e00944-21. [Google Scholar] [CrossRef]
- Ferrer-González, F.X.; Widner, B.; Holderman, N.R.; Glushka, J.; Edison, A.S.; Kujawinski, E.B.; Moran, M.A. Resource partitioning of phytoplankton metabolites that support bacterial heterotrophy. ISME J. 2021, 15, 762–773. [Google Scholar] [CrossRef] [PubMed]
- Bonch-Osmolovskaya, E.A.; Kublanov, I.V. Calditrichaceae. In Bergey’s Manual of Systematics of Archaea and Bacteria; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2021; pp. 1–3. [Google Scholar]
- Le Moine Bauer, S.; Lu, G.S.; Goulaouic, S.; Puzenat, V.; Schouw, A.; Barreyre, T.; Pawlowsky-Glahn, V.; Egozcue, J.J.; Martelat, J.E.; Escartin, J.; et al. Structure and metabolic potential of the prokaryotic communities from the hydrothermal system of Paleochori Bay, Milos, Greece. Front. Microbiol. 2023, 13, 5084. [Google Scholar] [CrossRef]
- McIlroy, S.J.; Kirkegaard, R.H.; Dueholm, M.S.; Fernando, E.; Karst, S.M.; Albertsen, M.; Nielsen, P.H. Culture-independent analyses reveal novel anaerolineaceae as abundant primary fermenters in anaerobic digesters treating waste activated sludge. Front. Microbiol. 2017, 8, 1134. [Google Scholar] [CrossRef]
- Yamada, T.; Sekiguchi, Y. Anaerolineaceae. In Bergey’s Manual of Systematics of Archaea and Bacteria; Kim, S.B., Goodfellow, M., Eds.; John Wiley & Sons, Ltd.: Chichester, UK, 2018; pp. 1–5. [Google Scholar]
- Silva, J.B.; Centurion, V.B.; Duarte, A.W.; Galazzi, R.M.; Arruda, M.A.; Sartoratto, A.; Rosa, L.H.; Oliveira, V.M. Unravelling the genetic potential for hydrocarbon degradation in the sediment microbiome of Antarctic islands. FEMS Microbiol. Ecol. 2023, 99, 143. [Google Scholar] [CrossRef] [PubMed]
- Kawaichi, S.; Ito, N.; Kamikawa, R.; Sugawara, T.; Yoshida, T.; Sako, Y. Ardenticatena maritima gen. nov., sp. nov., a ferric iron-and nitrate-reducing bacterium of the phylum ‘Chloroflexi’isolated from an iron-rich coastal hydrothermal field, and description of Ardenticatenia classis nov. Int. J. Syst. Evol. Microbiol. 2013, 63, 2992–3002. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; St John, E.; Anantharaman, K.; Reysenbach, A.L. Global patterns of diversity and metabolism of microbial communities in deep-sea hydrothermal vent deposits. Microbiome 2022, 10, 241. [Google Scholar] [CrossRef] [PubMed]
- Zemskaya, T.; Konstantinova, N.; Shubenkova, O.; Pogodaeva, T.; Ivanov, V.; Bukin, S.; Khabuev, A.; Khlystov, O.; Vilkin, G.; Lomakina, A. Microbial Communities of Ferromanganese Sedimentary Layers and Nodules of Lake Baikal (Bolshoy Ushkany Island). Diversity 2022, 14, 868. [Google Scholar] [CrossRef]
- Sieland, R.; Steinbrückner, D.; Hamel, M.; Merkel, B.; Schipek, M. Geochemical investigations and gas quantification of submarine fluid discharges in the hydrothermal system of Panarea (Aeolian Islands, Italy). In Proceedings of the 1st International Workshop Research in Shallow Marine and Freshwater Systems, Freiberg, Germany, 14–16 May 2009. [Google Scholar]
- McAllister, S.M.; Vandzura, R.; Keffer, J.L.; Polson, S.W.; Chan, C.S. Aerobic and anaerobic iron oxidizers together drive denitrification and carbon cycling at marine iron-rich hydrothermal vents. ISME J. 2021, 15, 1271–1286. [Google Scholar] [CrossRef]
- Langwig, M.V.; De Anda, V.; Dombrowski, N.; Seitz, K.W.; Rambo, I.M.; Greening, C.; Teske, A.P.; Baker, B.J. Large-scale protein level comparison of Deltaproteobacteria reveals cohesive metabolic groups. ISME J. 2022, 16, 307–320. [Google Scholar] [CrossRef]
- Zhong, Y.W.; Zhou, P.; Cheng, H.; Xu, X.W.; Wu, Y.H. Genome sequence of five Zetaproteobacteria metagenome-assembled genomes recovered from hydrothermal vent Longqi, Southwest Indian Ridge. Mar. Genom. 2022, 63, 100936. [Google Scholar] [CrossRef]
- Antranikian, G.; Suleiman, M.; Schäfers, C.; Adams, M.W.; Bartolucci, S.; Blamey, J.M.; Birkeland, N.K.; Bonch-Osmolovskaya, E.; da Costa, M.S.; Cowan, D.; et al. Diversity of bacteria and archaea from two shallow marine hydrothermal vents from Vulcano Island. Extremophiles 2017, 21, 733–742. [Google Scholar] [CrossRef]
- Parrilli, E.; Tedesco, P.; Fondi, M.; Tutino, M.L.; Giudice, A.L.; de Pascale, D.; Fani, R. The art of adapting to extreme environments: The model system Pseudoalteromonas. Phys. Life Rev. 2021, 36, 137–161. [Google Scholar] [CrossRef]
- Twing, K.I.; Ward, L.M.; Kane, Z.K.; Sanders, A.; Price, R.E.; Pendleton, H.L.; Giovannelli, D.; Brazelton, W.J.; McGlynn, S.E. Microbial ecology of a shallow alkaline hydrothermal vent: Strýtan Hydrothermal Field, Eyjafördur, northern Iceland. Front. Microbiol. 2022, 13, 3278. [Google Scholar] [CrossRef]
- Kirchman, D.L. The ecology of Cytophaga–Flavobacteria in aquatic environments. FEMS Microbiol. Ecol. 2002, 39, 91–100. [Google Scholar] [CrossRef] [PubMed]
- Gavriilidou, A.; Gutleben, J.; Versluis, D.; Forgiarini, F.; van Passel, M.W.; Ingham, C.J.; Smidt, H.; Sipkema, D. Comparative genomic analysis of Flavobacteriaceae: Insights into carbohydrate metabolism, gliding motility and secondary metabolite biosynthesis. BMC Genom. 2020, 21, 569. [Google Scholar] [CrossRef] [PubMed]
- Levin, L.A.; Baco, A.R.; Bowden, D.A.; Colaco, A.; Cordes, E.E.; Cunha, M.R.; Demopoulos, A.W.; Gobin, J.; Grupe, B.M.; Le, J.; et al. Hydrothermal vents and methane seeps: Rethinking the sphere of influence. Front. Mar. Sci. 2016, 3, 72. [Google Scholar] [CrossRef]
- Ashford, O.S.; Guan, S.; Capone, D.; Rigney, K.; Rowley, K.; Orphan, V.; Mullin, S.W.; Dawson, K.S.; Cortés, J.; Rouse, G.W.; et al. A chemosynthetic ecotone—“chemotone”—In the sediments surrounding deep-sea methane seeps. Limnol. Oceanogr. 2021, 66, 1687–1702. [Google Scholar] [CrossRef]
- Giovannelli, D.; d’Errico, G.; Manini, E.; Yakimov, M.; Vetriani, C. Diversity and phylogenetic analyses of bacteria from a shallow-water hydrothermal vent in Milos Island (Greece). Front. Microbiol. 2013, 4, 184. [Google Scholar] [CrossRef] [PubMed]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arcadi, E.; Buschi, E.; Rastelli, E.; Tangherlini, M.; De Luca, P.; Esposito, V.; Calogero, R.; Andaloro, F.; Romeo, T.; Danovaro, R. Novel Insights on the Bacterial and Archaeal Diversity of the Panarea Shallow-Water Hydrothermal Vent Field. Microorganisms 2023, 11, 2464. https://doi.org/10.3390/microorganisms11102464
Arcadi E, Buschi E, Rastelli E, Tangherlini M, De Luca P, Esposito V, Calogero R, Andaloro F, Romeo T, Danovaro R. Novel Insights on the Bacterial and Archaeal Diversity of the Panarea Shallow-Water Hydrothermal Vent Field. Microorganisms. 2023; 11(10):2464. https://doi.org/10.3390/microorganisms11102464
Chicago/Turabian StyleArcadi, Erika, Emanuela Buschi, Eugenio Rastelli, Michael Tangherlini, Pasquale De Luca, Valentina Esposito, Rosario Calogero, Franco Andaloro, Teresa Romeo, and Roberto Danovaro. 2023. "Novel Insights on the Bacterial and Archaeal Diversity of the Panarea Shallow-Water Hydrothermal Vent Field" Microorganisms 11, no. 10: 2464. https://doi.org/10.3390/microorganisms11102464