1. Introduction
The Mars spacecraft Pathfinder, Spirit, Opportunity, Phoenix, Curiosity, and InSight were rated as either Category IVa or IVc (Phoenix) missions, and as such, their bioburdens were limited to ≤5 × 10
5 total spores per vehicle or ≤300 spores/m
2 at launch [
1]. Published data indicate that the bioburdens estimated at launch for these spacecraft met or exceeded the guidelines (i.e., lower bioburdens than required [
2,
3]). However, spacecraft bioburden models [
4,
5,
6] suggest that total spacecraft bioburdens of all microbiota are likely to be up to three orders of magnitude higher than the planetary protection guidelines required for spore-forming species.
After launch, Mars spacecraft spend 6–8 months in a cruise phase in which space biocidal factors begin to inactivate bioburdens on external and internal surfaces [
6,
7]. It is likely that 1–3 orders of magnitude reductions in viable bioburdens can be achieved during the Earth–Mars cruise phase [
7]. However, this would leave many individual viable cells, spores, and cell–spore aggregates to persist through the entry, descent, and landing (EDL) profiles of spacecraft.
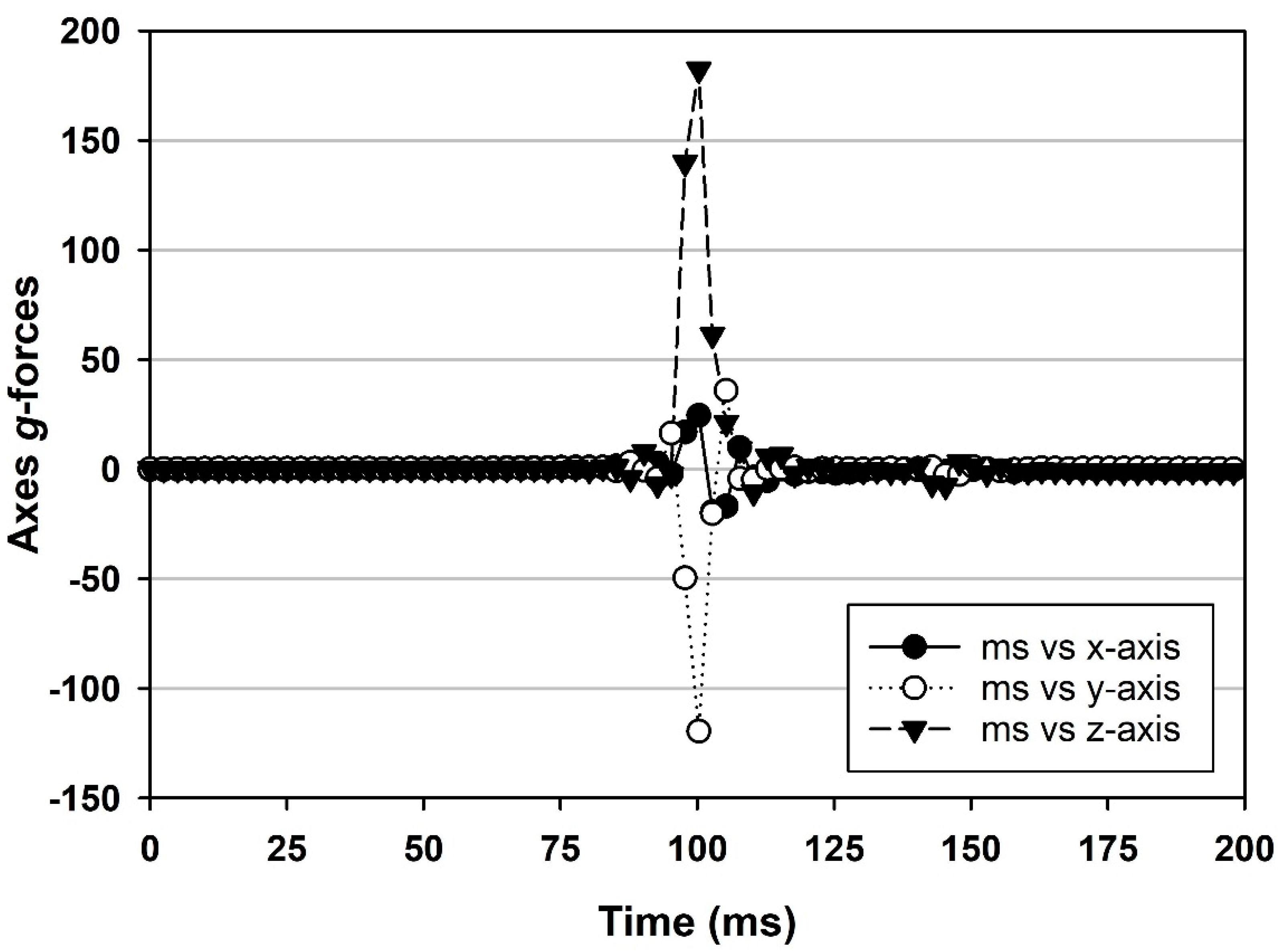
Spacecraft EDL scenarios are typically divided into landings with air-bag delivery systems (e.g., Pathfinder, Spirit, Opportunity rovers) or landings with active descent engine braking at the surface (e.g., Phoenix, Curiosity, InSight, Perseverance spacecraft). The air-bag delivery systems create high-
g spikes as the spacecraft bounces along the landing ellipse. For example, the Pathfinder rover experienced 14 bounces, with the first bounce creating a 15.5
g dynamic shock compressional impact (henceforth called shock-impacts) to the landed systems [
8]. All subsequent bounces created a diminishing series of loads that averaged approx. 8–10
g’s. In contrast, active descent landing configurations may have a high-
g phase during landing, but it is generally observed during the hypersonic phase just prior to parachute deployment. For example, the Phoenix lander experienced a slow build-up of
g-loads to 9.2
g’s during its hypersonic phase [
9]. However, in the off-nominal landings of spacecraft (e.g., Beagle 2, Mars Polar Lander) and in the nominal impacts of jettisoned subsystems like the Perseverance aeroshell (
Figure 1) or Opportunity heat shield (
Figure 2), the
g-forces can be high enough to cause the disintegration of spacecraft components.
The goals of the current study were to determine if
g-force thresholds exist for dislodging
Bacillus spp. spores from aluminum coupons under shock impact or vibrational
g-loading conditions. Vibrational
g-loads can be encountered during the Earth launch or EDL of planetary spacecraft. If shock-impacts or vibrational-
g loads experienced by spacecraft during launch or if EDL cause the release of microbiota from surfaces, the dislodged cells/spores may have consequences for predicting the forward contamination risks to Special Regions [
10] on Mars.
The nomenclature of Hazel et al. [
11] is adopted here for differentiating
static compressional g-forces (i.e., high pressures imposed on microbiota through devices that create and then hold the samples at elevated pressures (not tested here)) and
dynamic shock compressional g-forces (i.e., very-short-duration high-
g events). We further simplify the second term to ‘shock-impacts’ for the discussions below. Lastly, we use the term ‘
threshold’ here in its generic form defined as ‘…a level, point, or value above which something is true or will take place…’ (Merriam-Webster Dictionary (online at
https://www.merriam-webster.com/dictionary/threshold (accessed on 17 September 2023)).
4. Discussion
Modeling the forward contamination risk of the Martian terrain requires a variety of factors to be defined and characterized. Typical predictions rely almost exclusively on characterizing the launched bioburdens on spacecraft surfaces prior to launch [
1]. However, if >99.99% of the bioburdens on Mars rovers and landers remain on the spacecraft surfaces, then the forward contamination of the local terrain is likely to be extremely low. Estimates on how much bioburden can be dislodged from spacecraft during Earth-launch and Mars EDL profiles are limited, and dislodged spores may be a dominant factor in predicting the risks to Special Regions on Mars. One example is a study in which descent engine exhaust was shown to scour landing struts on the Phoenix lander [
15]; thus, it would have removed and dispersed many viable cells/spores remaining on external surfaces at landing.
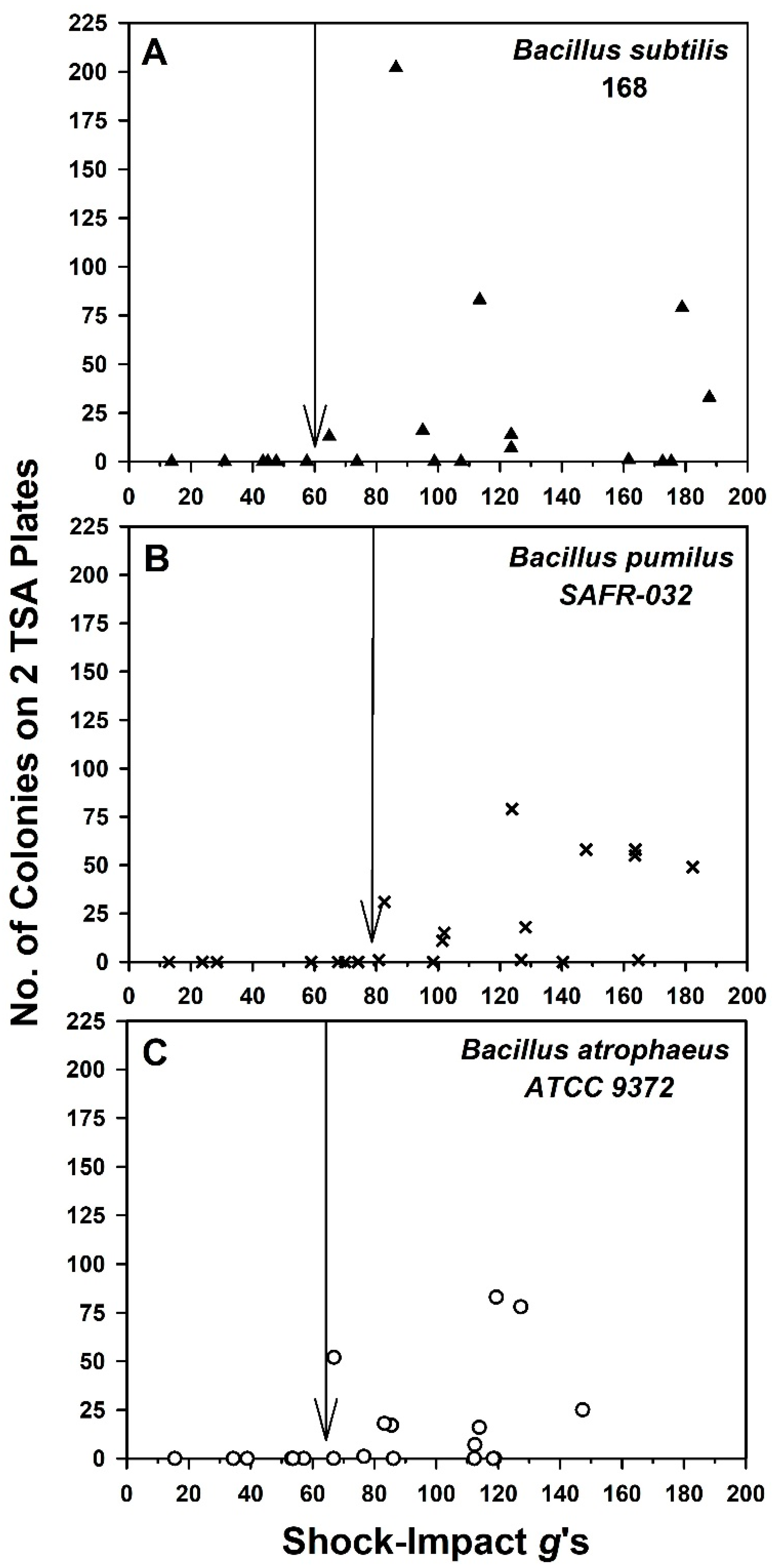
The experiments described here report for the first time that high g-loading on spacecraft can dislodge bacterial spores from analog spacecraft surfaces. The research considered two possible g-loading scenarios: high-g shock-impacts and vibrational g-loading. High-g shock-impact events would be applicable for the off-nominal impacts of spacecraft or the expected impacts of jettisoned subsystems (e.g., backshells or heat shields of rovers). Vibrational g-loading would occur in both Earth-launch and Mars EDL profiles.
For the bacteria tested here, 60-
g thresholds were observed for
B. subtilis 168 and
B. atrophaeus 9372 spores on ChemFilm-coated aluminum coupons during shock-impact events. The high-
g threshold for dislodging spores was slightly higher at 80
g’s for
B. pumilus SAFR-032. However, what was remarkable was the observation that the numbers of dislodged spores ranged from single digits to 10s or low-100s of spores for each impact event, even up to 190
g’s for the three
Bacillus spp. tested (
Figure 6). Recall that the population densities of spores on coupons were ~2 × 10
6 at T = 0. Thus, the vast majority of spores remained firmly attached to ChemFilm-coated surfaces with all three
Bacillus spp. as shock-impact forces were increased up to 190
g’s.
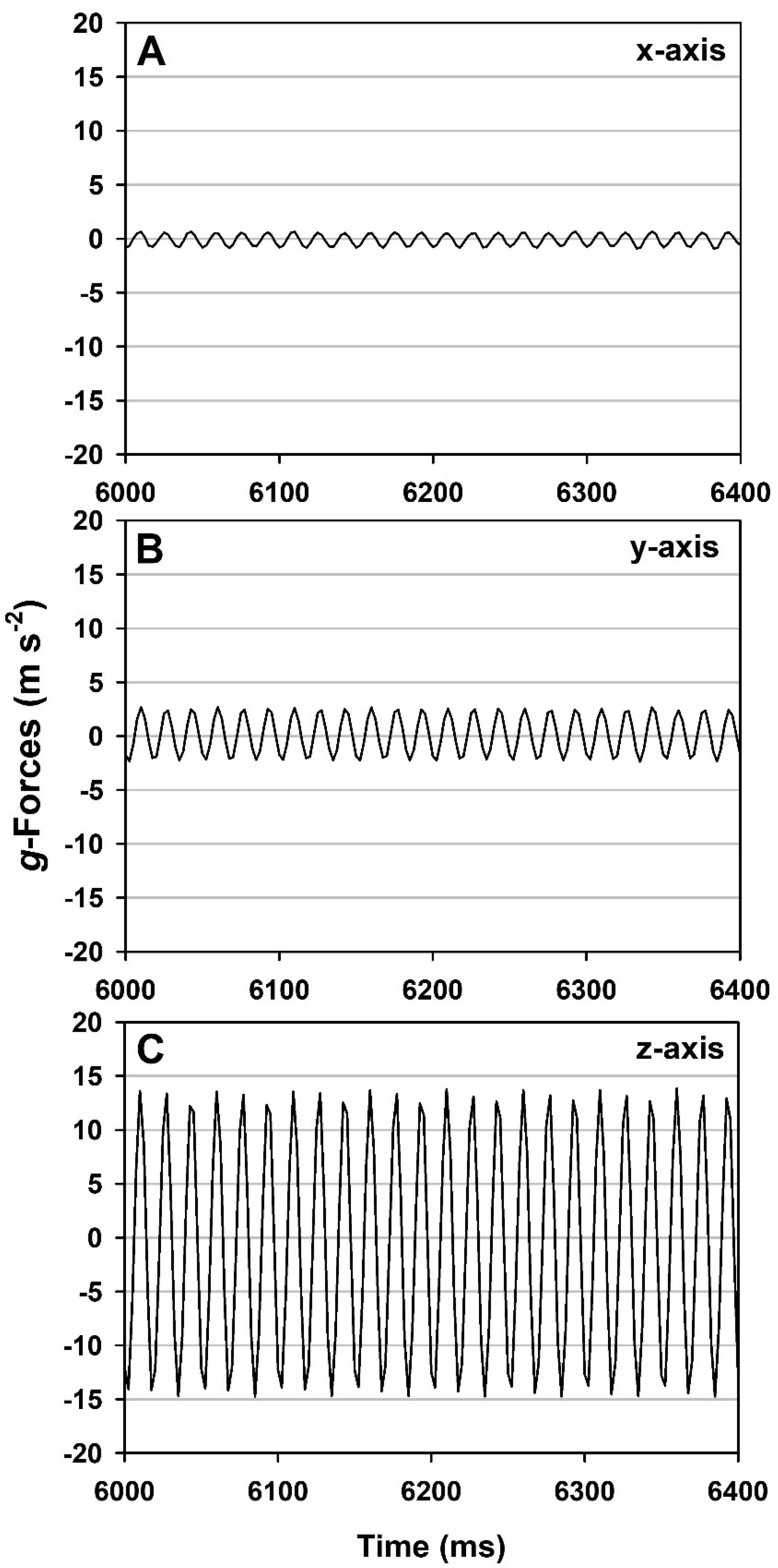
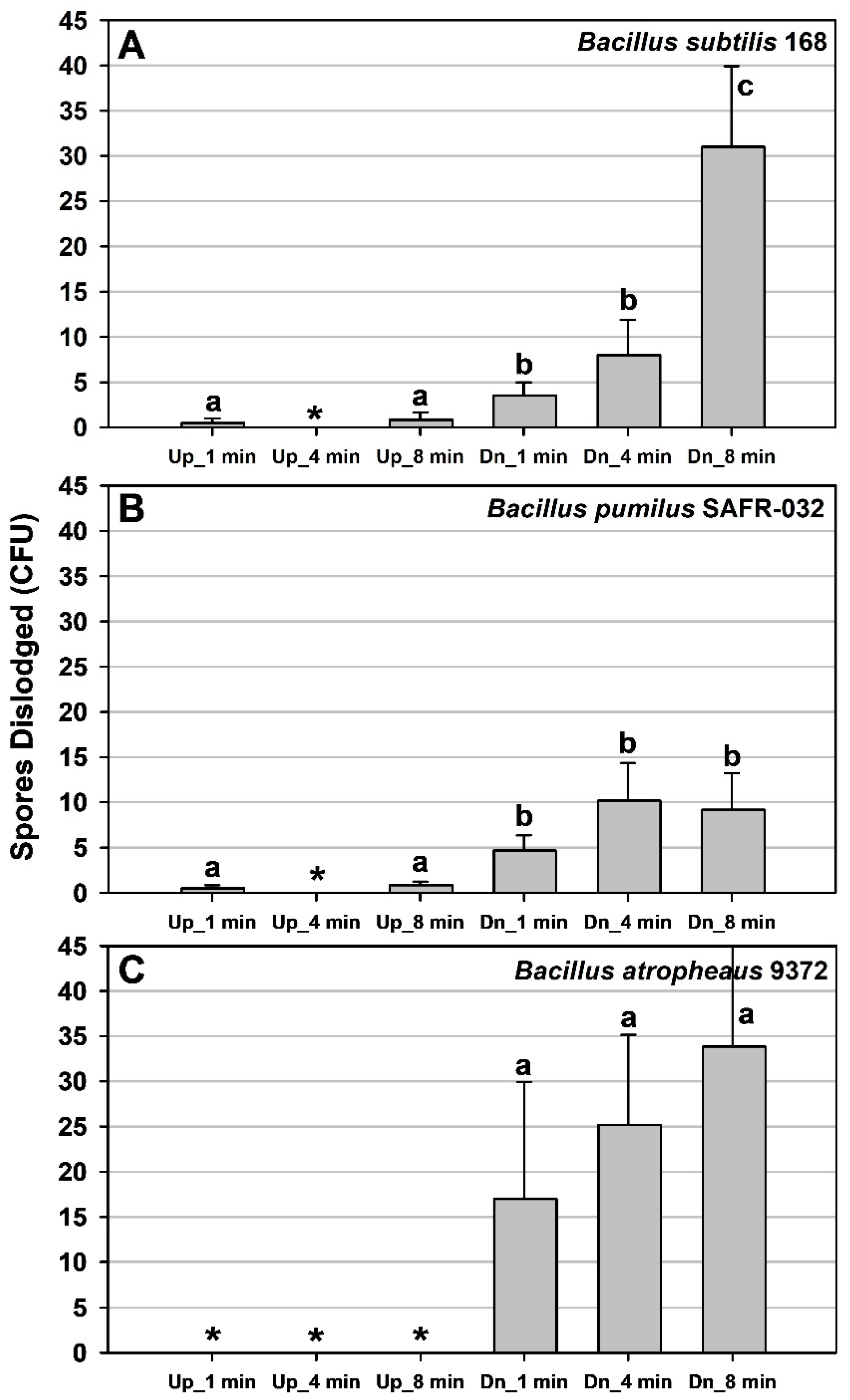
In contrast, continuous vibrational
g-loading of the same
Bacillus spp. and hardware produced dislodged spores at lower
g-forces (z-axis) of up to ~12–14
g’s if the vibrations were extended for 1 to 8 min and the MSH units were inverted on the vibration table (
Figure 7). Spores dislodged during the three-axes vibrations (inverted MSH units) increased with longer timesteps for all three
Bacillus spp. tested. Surprisingly, very few to zero spores of all three
Bacillus spp. were dislodged if the ChemFilm-coated coupons were in the upright configuration. The results suggest that vibrational
g-loading is more effective in dislodging spores when the surfaces are concomitantly inverted relative to Earth’s planetary
g-vector.
Most launch vehicles incur three-axes vibrations for approx. 8 min during launch to low-Earth orbit (LEO). However, only a few spores per MSH unit were reported at the upper range of time under vibrational
g-loading. For example, approx. 30 and 35 spores were dislodged for
B. subtilis 168 and
B. atrophaeus 9372, respectively, per coupon on samples exposed to three-axes vibrations for 8 min (i.e.,
Down MSH units) (
Figure 7A,C). In contrast, only approx. 10 spores per MSH unit were dislodged for
B. pumilus SAFR-032 when the coupons were exposed to vibrations at 4 and 8 min (i.e.,
Down MSH units) (
Figure 7B). These results suggest that spores of
B. pumilus adhere more strongly to Class 1A ChemFilm aluminum compared to the other two species, which is consistent with earlier research on spore attachment to aluminum surfaces [
12].
The three key assumptions in these experiments were as follows: (1) bioburdens on Mars spacecraft are attached as low-density single cells/spores or small aggregates of the same [
16]; (2) that ChemFilm-coated aluminum coupons were good proxies for the average material of a Mars spacecraft [
13,
17]; and (3) the
g-loads in these experiments were representative of both the launch and EDL profiles of Mars spacecraft (see below). Furthermore, it is unlikely that the shock-impacts of vibrational
g-loads tested here directly impacted microbial survival because numerous studies have shown reasonable rates of microbial survival up to 57–78 GPa [
18,
19]; i.e., significantly higher
g-loads for shock impact events than tested here.
Furthermore, ChemFilm-coated (Class 1A) aluminum coupons were used here as the test material for both
g-loading assays. ChemFilm was selected based on previous work with multiple analog spacecraft components as support materials for microbial assays [
13,
17]. However, it is plausible that spore adhesion might diverge from these results (e.g.,
Figure 6 and
Figure 7) if other spacecraft materials or microbial species are tested. For example, four forces may be involved in the attachment of particles (including microbes) to aluminum and include contact potential, electrostatic, van der Waals, and water capillary forces [
20]. In addition, secreted cell/spore mucilage might contribute to adhesion to spacecraft materials. It is likely that diverse spacecraft components and materials will vary widely with respect to these properties, which would alter how particles—including bacteria—might adhere to surfaces.
New experiments that examine other spacecraft materials and microbial species are required to fully characterize how launch and EDL g-loading can affect spore removal from planetary spacecraft. The results presented herein should be viewed as preliminary data on how terrestrial microorganisms can adhere to planetary spacecraft during prelaunch processing and how the same bioburdens might be dislodged during launch and EDL events.
Lastly, if the data in
Figure 6 and
Figure 7 were replotted using the common approach of representing the removal of particles from surfaces as fractional terms (i.e., called ‘
particle removal fractions’) [
21,
22], the results would indicate that the particle removal fractions would be between 10
−4 (low 100s of recovered spores) and 10
−6 (low single digits). These values are extremely low compared to assays that seek to identify a ‘
threshold velocity or
shear stress’, in which 50% of particles are removed at a given velocity in a shearing fluid [
22]. Here we sought the thresholds (i.e., generic definition) at which single digits and 10s to low-100s of spores were initially dislodged from spacecraft surfaces. We call attention to these divergent definitions to avoid confusion.
4.1. Implications for Spacecraft during Launch
Possible effects of shock-impact and vibrational
g-loads for launch are derived from data presented in the Delta II [
23] and Atlas V [
24] user guides. Pathfinder, Spirit, Opportunity, and Phoenix spacecraft were launched on Delta II rockets, and Insight and Perseverance were launched on Atlas V rockets. We assume here that the data given in these two user guides from United Launch Alliance (ULA) are averages of these missions.
First, spacecraft that weigh approx. 1500 kg in a Delta II launch configuration (i.e., the assumed average weight of the spacecraft listed above for the Delta II) will encounter a steady-state acceleration of approx. 7
g’s during launch at the peak acceleration of the spacecraft (i.e., at the end of first-stage main engine cutoff (MECO) [
23]). However, the acceleration
g-forces are not shock-impact
g-loads; they build up slowly as the Delta II pushes through decreasing air resistance during launch. Furthermore, during Delta II launches, spacecraft typically encounter a variable vibrational
g-loading environment up to 2.2
g’s (120 Hz) in the thrust vector and up to 0.85
g (120 Hz) in lateral vectors [
23].
The launch performance of the Atlas V vehicle [
24] is similar to that of the Delta II given above. The Atlas V experiences a very slow increase in acceleration up to 5
g’s for the thrust vector and up to 2
g’s for lateral vectors during launch, but the
g’s are not shock-impact forces. In addition, the Atlas V payloads can experience low vibrational
g-forces between 50 and 100 Hz that peak at approx. 1
g during launch.
The vibrational
g-load frequencies tested here (12–14
g’s at 77 Hz;
Figure 5) were in the same ranges of the Delta II and Atlas V vibrational frequencies but much more severe in the
g-loads (i.e., 12–14
g’s here versus no more than 2.2
g’s in the thrust vector of a Delta II). First, our results suggest that
Bacillus spores are unlikely to be dislodged from payload surfaces during launch on Delta II or Atlas V rockets. One exception to this conclusion might be unique staging events that might have unusually strong shock-impact forces with engine cutoffs and second- or third-stage booster ignitions. The model AS200 Retsch vibration table used here (
Figure 3B) was very limited in its ability to adjust the
g-forces in the assays. Thus, future vibrational
g-loads at approx. 2.5
g’s and up to 120 Hz (i.e., similar to the Delta II and Atlas V launch vehicles) should be tested to determine if lower
g-forces but higher frequencies result in the dislodgment of bacterial spores from spacecraft surfaces.
4.2. Implications for Mars Spacecraft during EDL
There are two basic architectures for landing spacecraft on Mars: air-bag bounce landings or active descent engine landings. The former can experience shock-impact forces during the bounces, but the latter approach is intended to gently land vehicles on the terrain.
The best data for the air-bag bounce EDL approach were found for the Pathfinder spacecraft [
8]. The Pathfinder vehicle bounced 14 times on the Martian terrain before coming to rest. The initial bounce reached 15.5
g’s, the subsequent seven bounces peaked between 10 and 12
g’s, and the final six bounces were approx. 5
g’s. The shock impact
g-force thresholds for dislodging spores described here were not reached on any of the Pathfinder bounces. Thus, it is unlikely that
Bacillus spores that might have been present on internal spacecraft surfaces were dislodged during the Pathfinder EDL profile.
In addition, all spacecraft entering the Martian atmosphere slowly build-up
g-loads along the entry vector during the hypersonic phase as atmospheric density increases. The peak
g-loads were 16, 12.5, and 9.2 for the Mars spacecraft Pathfinder, Curiosity, and Phoenix, respectively [
8,
9,
25]. However, these
g-loads built up slowly and were not shock-impact events. Thus, for nominal EDL profiles, spores are unlikely to be dislodged from Mars spacecraft, even if air-bag-bounced landings are employed.
This leaves us to consider either off-nominal landings (e.g., Beagle 2 and Mars Polar Lander) or the impacts of jettisoned subsystems like the Perseverance backshell (
Figure 1) or the Opportunity heat shield (
Figure 2). In preliminary analysis of the impact
g-loads for the Perseverance backshell, the structure likely hit the terrain at approx. 62
g’s (A. Chen, JPL-Cal-tech, personal communications), consistent with the thresholds—described here—for dislodging
B. subtilis and
B. atrophaeus spores from aluminum coupons (
Figure 6). Examination of the image in
Figure 1 clearly shows that the impact forces were adequate to crack and flatten the Perseverance backshell. We conclude here that the forces were also adequate to likely dislodge low numbers of spores from the structures. Furthermore, the Opportunity heat shield (
Figure 2) broke up into two major components upon impact. Both examples of these expected impacts of jettisoned subsystems clearly show numerous pieces and parts strewn around the impact sites.
5. Conclusions
Mars spacecraft that successfully complete a nominal EDL profile are unlikely to experience g-loads that will dislodge spores into the Martian atmosphere or terrains. The exceptions to this rule might be jettisoned subsystems (e.g., backshells and heat shields described above) that are released above the Martian surface and have no active means of slowing down before impact, as well as landing struts and pads exposed to active descent engines. In the latter cases, bacterial spores are likely dislodged from spacecraft surfaces and may be immediately caught up in the swirling winds at the landing sites. In addition, bacterial spores could be released directly to the Martian regolith below the crashed components. Further research is suggested in order to characterize whether dislodged spores would survive long enough in the winds or regolith to encounter benign conditions and permit the growth and cellular replication of terrestrial microorganisms on Mars.
Furthermore, the tests presented here were done in Earth lab atmospheric conditions of 1013 mbar, 45% RH, room temperature between 22 and 24 °C, and with a gas composition of 78% to 21% for pN2 and pO2 gases. These conditions are not present on the Martian surface. Thus, additional research is suggested in order to repeat and extend these experiments into measuring the g-forces required to dislodge spores under simulated Martian conditions. The equipment to run these experiments under Martian atmospheric conditions was not available.
Lastly, future research is suggested in order to evaluate the adhesion and dislodgement of vegetative cells of bacteria and archaea—plus spores of eukaryotes like fungi—that might be present as bioburdens on spacecraft surfaces. Our initial results, presented here, suggest that some components hitting the Martian terrain could release spores if the shock-impact g-forces are greater than 60 g’s, but further study is warranted.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}






