
Long-Term Monitoring of Bioaerosols in an Environment without UV and Desiccation Stress, an Example from the Cave Postojnska Jama, Slovenia
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Description of the Study Site
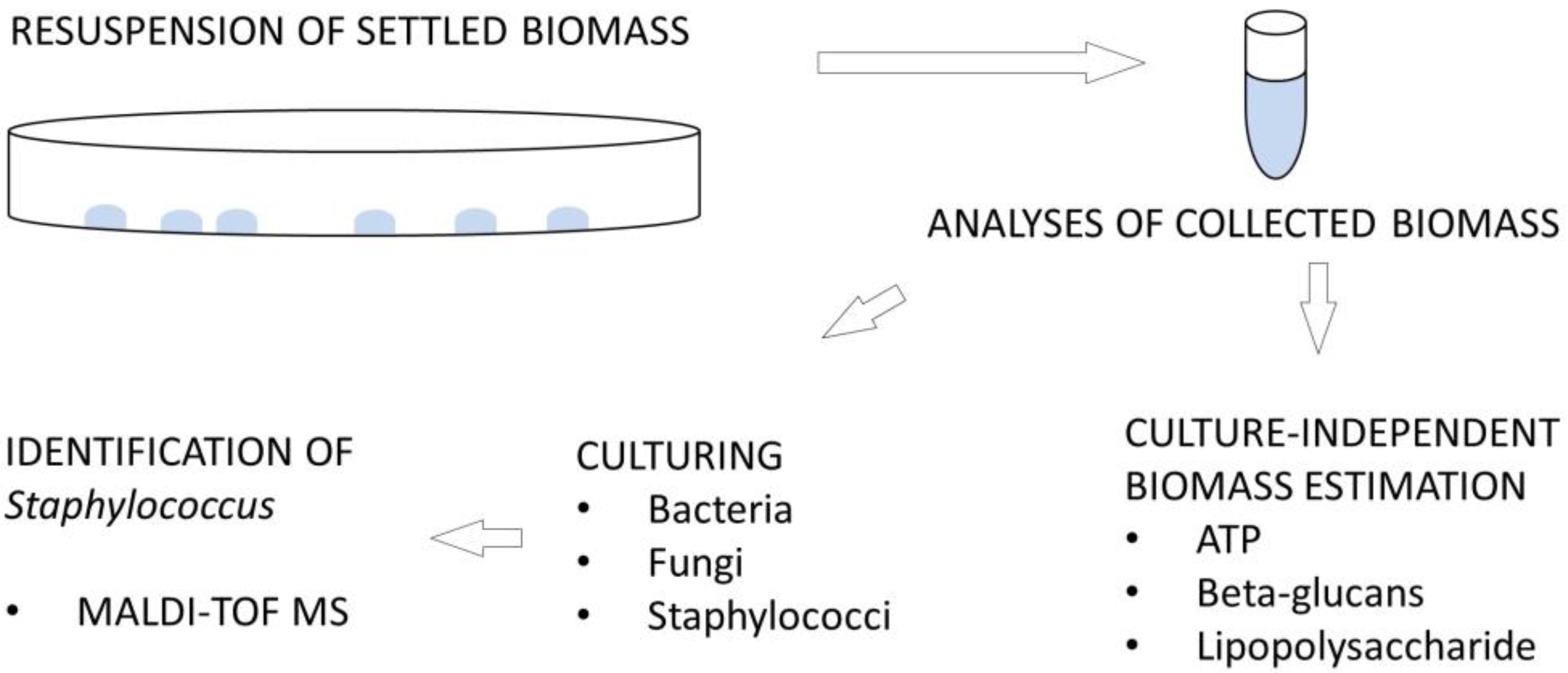
2.2. Preparation of Samples
2.3. Estimation of Microbial Biomass and Air Velocity
2.4. Identification of Staphylococcus
2.5. Statistical Analyses
3. Results
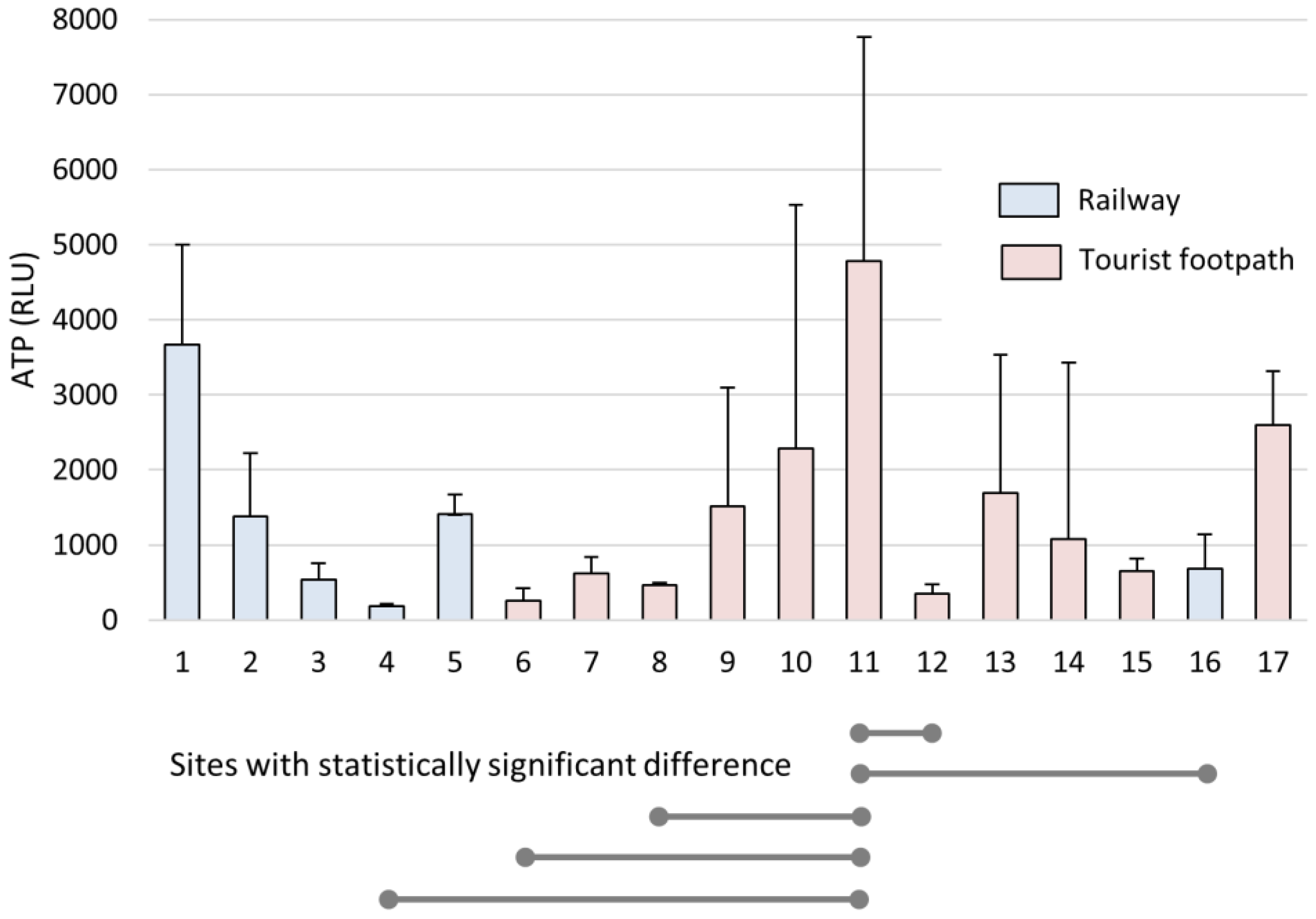
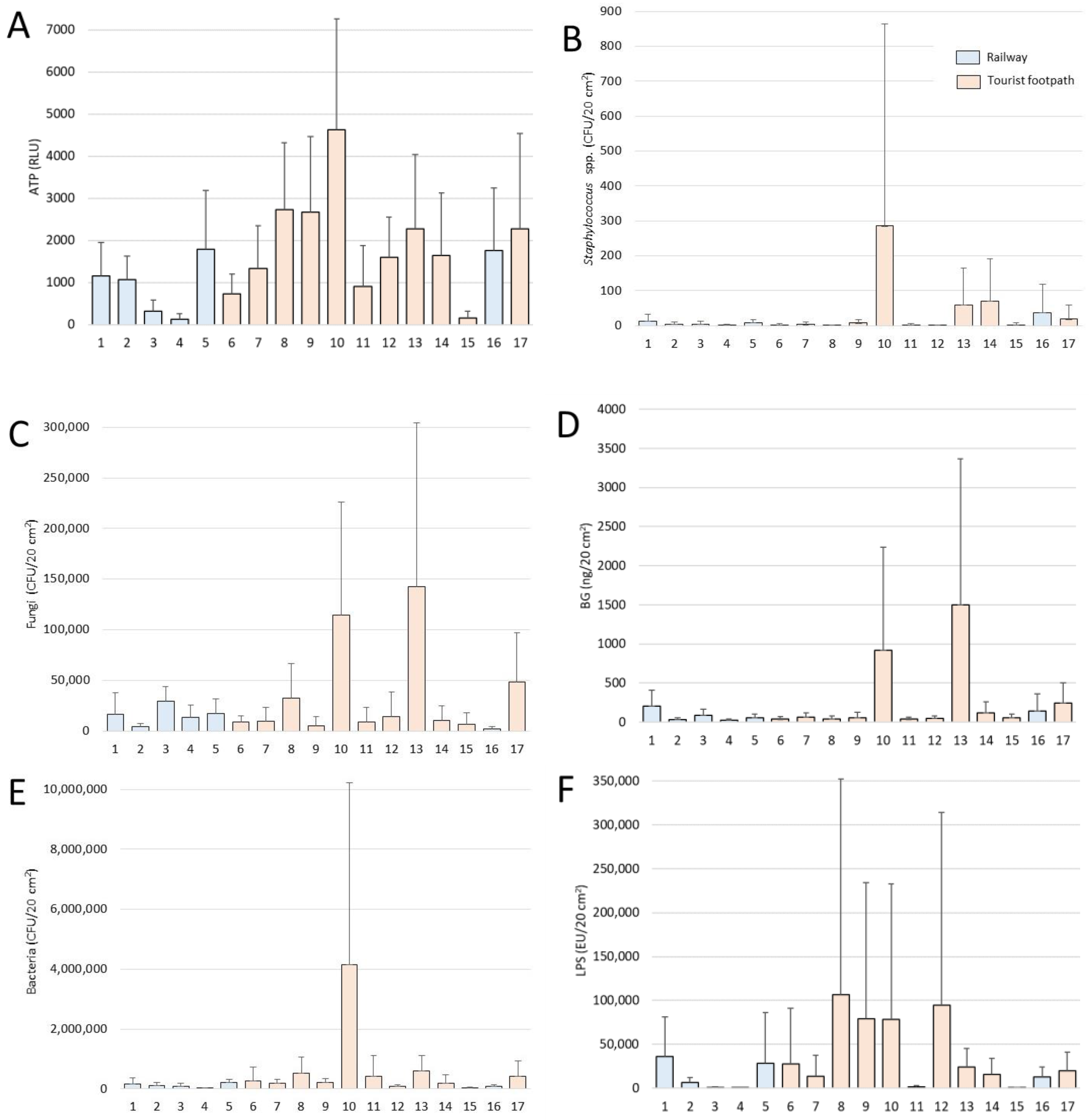
3.1. Bioaerosols in the Cave Air
3.2. Staphylococcus as an Indicator of the Human Microbiome in Cave Air
4. Discussion
4.1. Characteristics and Dynamics of Settled Bioaerosols
4.2. Airborne Staphylococci, Viability and Potential Impact on Humans and Caves
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brimblecombe, P. Air Composition and Chemistry, 2nd ed.; Cambridge University Press: Cambridge, UK, 1996. [Google Scholar]
- Chen, X.; Kumari, D.; Achal, V. A review on airborne microbes: The characteristics of sources, pathogenicity and geography. Atmosphere 2020, 11, 919. [Google Scholar] [CrossRef]
- Kim, K.; Kabir, E.; Jahan, S. Airborne bioaerosols and their impact on human health. J. Environ. Sci. 2018, 67, 23–35. [Google Scholar] [CrossRef] [PubMed]
- Campbell, B.; Al Kouba, J.; Timbrell, V.; Noor, M.; Massel, K.; Gilding, E.; Angel, N.; Kemish, B.; Hugenholtz, P.; Godwin, I.; et al. Tracking seasonal changes in diversity of pollen allergen exposure: Targeted metabarcoding of a subtropical aerobiome. Sci. Total Environ. 2020, 747, 2020747. [Google Scholar] [CrossRef] [PubMed]
- Masotti, F.; Cattaneo, S.; Stuknyte, M.; De Noni, I. Airborne contamination in the food industry: An update on monitoring and disinfection techniques of air. Trends Food Sci. Technol. 2019, 90, 147–156. [Google Scholar] [CrossRef]
- Mehta, Y.; Gupta, A.; Todi, S.; Myatra, S.; Samaddar, D.; Patil, V.; Bhattacharya, P.; Ramasubban, S. Guidelines for prevention of hospital acquired infections. Indian J. Crit. Care Med. 2014, 18, 149–163. [Google Scholar]
- Alwis, K.; Mandryk, J.; Hocking, A. Exposure to biohazards in wood dust: Bacteria, fungi, endotoxins, and (1 3)-beta-D-glucans. Appl. Occup. Environ. Hyg. 1999, 14, 598–608. [Google Scholar] [CrossRef]
- Douwes, J. (1→3)-beta-D-glucans and respiratory health: A review of the scientific evidence. Indoor Air 2005, 15, 160–169. [Google Scholar] [CrossRef]
- Stopinšek, S.; Ihan, A.; Salobir, B.; Terčelj, M.; Simčič, S. Fungal cell wall agents and bacterial lipopolysaccharide in organic dust as possible risk factors for pulmonary sarcoidosis. J. Occup. Med. Toxicol. 2016, 11, 1–7. [Google Scholar] [CrossRef] [Green Version]
- WHO. WHO Guidelines for Indoor Air Quality: Dampness and mould; WHO Regional Office Europe: Copenhagen, Denmark, 2009. [Google Scholar]
- Rylander, R.; Lin, R. (1→3)-beta-D-glucan—Relationship to indoor air-related symptoms, allergy and asthma. Toxicology 2000, 152, 47–52. [Google Scholar] [CrossRef]
- Young, S.; Castranova, V. Toxicology of 1→3-Beta-Glucans: Glucans as a Marker for Fungal Exposure; CRC Press: Boca Raton, FL, USA, 2005. [Google Scholar]
- Garcia-Vello, P.; Di Lorenzo, F.; Zucchetta, D.; Zamyatina, A.; De Castro, C.; Molinaro, A. Lipopolysaccharide lipid A: A promising molecule for new immunity-based therapies and antibiotics. Pharmacol. Therapeut. 2022, 230, 107970. [Google Scholar] [CrossRef]
- Kanchongkittiphon, W.; Mendell, M.; Gaffin, J.; Wang, G.; Phipatanakul, W. Indoor Environmental Exposures and Exacerbation of Asthma: An Update to the 2000 Review by the Institute of Medicine. Environ. Health Perspect. 2015, 123, 6–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mulec, J.; Vaupotič, J.; Walochnik, J. Prokaryotic and eukaryotic airborne microorganisms as tracers of microclimatic changes in the underground (Postojna Cave, Slovenia). Microb. Ecol. 2012, 64, 654–667. [Google Scholar] [CrossRef] [PubMed]
- Gillieson, D. Caves: Processes, Development, and Management, 2nd ed.; John Wiley & Sons: Hoboken, NJ, USA, 2021. [Google Scholar]
- Mulec, J.; Oarga-Mulec, A.; Šturm, S.; Tomazin, R.; Matos, T. Spacio-temporal distribution and tourist impact on airborne bacteria in a cave (Škocjan Caves, Slovenia). Diversity 2017, 9, 28. [Google Scholar] [CrossRef] [Green Version]
- Tomazin, R.; Simčič, S.; Matos, T.; Kopitar, A.; Stopinšek, S.; Mauko Pranjić, A.; Zalar Serjun, V.; Mulec, J. Vpliv turizma na kakovost zraka v Postojnski jami in Škocjanskih jamah. In 10 Baničevi Dnevi, Proceedings of Mikrobiologija v Javnem Zdravstvu, Postojna, Slovenija, 23–24 November 2018; Petrovec, M., Ed.; Sekcija za Klinično Mikrobiologijo in Bolnišnične Okužbe SZD: Ljubljana, Slovenija, 2018; pp. 2015–2116. [Google Scholar]
- Cogen, A.; Nizet, V.; Gallo, R. Skin microbiota: A source of disease or defence? Brit. J. Dermatol. 2008, 158, 442–455. [Google Scholar] [CrossRef] [Green Version]
- O’Sullivan, J.; Rea, M.; O’Connor, P.; Hill, C.; Ross, R. Human skin microbiota is a rich source of bacteriocin-producing staphylococci that kill human pathogens. FEMS Microbiol. Ecol. 2019, 95, fly241. [Google Scholar] [CrossRef] [Green Version]
- Parlet, C.; Brown, M.; Horswill, A. Commensal staphylococci influence Staphylococcus aureus skin colonization and disease. Trends Microbiol. 2019, 27, 497–507. [Google Scholar] [CrossRef]
- Griffin, D.; Gray, M.; Lyles, M.; Northup, D. The transport of nonindigenous microorganisms into caves by human visitation: A case study at Carlsbad Caverns National Park. Geomicrobiol. J. 2014, 31, 175–185. [Google Scholar] [CrossRef]
- Bercea, S.; Năstase-Bucur, R.; Mirea, I.; Mantoiu, D.; Kenesz, M.; Petculescu, A.; Baricz, A.; Andrei, A.; Banciu, H.; Papp, B.; et al. Novel approach to microbiological air monitoring in show caves. Aerobiologia 2018, 34, 445–468. [Google Scholar] [CrossRef]
- Borda, D.; Năstase-Bucur, R.; Spînu, M.; Uricariu, R.; Mulec, J. Aerosolized microbes from organic rich materials: Case study of bat guano from caves in Romania. J. Cave Karst Stud. 2014, 76, 114–126. [Google Scholar] [CrossRef]
- Mulec, J.; Oarga-Mulec, A. ATP luminescence assay as a bioburden estimator of biomass accumulation in caves. Int. J. Speleol. 2016, 45, 207–208. [Google Scholar] [CrossRef] [Green Version]
- Mulec, J.; Petrič, M.; Koželj, A.; Brun, C.; Batagelj, E.; Hladnik, A.; Holko, L. A multiparameter analysis of environmental gradients related to hydrological conditions in a binary karst system (underground course of the Pivka River, Slovenia). Acta Carsol. 2019, 48, 313–327. [Google Scholar] [CrossRef]
- Mulec, J. Human impact on underground cultural and natural heritage sites, biological parameters of monitoring and remediation actions for insensitive surfaces: Case of Slovenian show caves. J. Nat. Conserv. 2014, 22, 132–141. [Google Scholar] [CrossRef]
- Muri, G.; Jovičić, A.; Mihevc, A. Source assessment of deposited particles in a Slovenian show cave (Postojnska Jama): Evidence of long-lasting anthropogenic impact. Int. J. Speleol. 2013, 42, 225–233. [Google Scholar] [CrossRef] [Green Version]
- Prelovšek, M.; Šebela, S.; Turk, J. Carbon dioxide in Postojna Cave (Slovenia): Spatial distribution, seasonal dynamics and evaluation of plausible sources and sinks. Environ. Earth Sci. 2018, 77, 289. [Google Scholar] [CrossRef]
- Skok, S.; Kogovšek, B.; Tomazin, R.; Šturm, S.; Ambrožič Avguštin, J.; Mulec, J. Antimicrobial-resistant Escherichia coli from karst waters, surfaces and bat guano in Slovenian caves. Acta Carsol. 2020, 49, 265–279. [Google Scholar]
- Šebela, S.; Prelovšek, M.; Turk, J. Impact of peak period visits on the Postojna Cave (Slovenia) microclimate. Theor. Appl. Climatol. 2013, 111, 51–64. [Google Scholar] [CrossRef]
- Šebela, S.; Turk, J. Natural and anthropogenic influences on the year-round temperature dynamics of air and water in Postojna show cave, Slovenia. Tourism Manag. 2014, 40, 233–243. [Google Scholar] [CrossRef]
- Peberdy, J. Fungal cell walls—A review. In Biochemistry of Cell Walls and Membranes in Fungi; Kuhn, P.J., Trinci, A.P.J., Jung, M.J., Goosey, M.W., Copping, L.G., Eds.; Springer: Berlin/Heidelberg, Germany, 1990; pp. 5–30. [Google Scholar]
- Dredge, J.; Fairchild, I.J.; Harrison, R.M.; Fernandez-Cortes, A.; Sanchez-Moral, S.; Jurado, V.; Gunn, J.; Smith, A.; Spötl, C.; Mattey, D.; et al. Cave aerosols: Distribution and contribution to speleothem geochemistry. Quat. Sci. Rev. 2013, 63, 23–41. [Google Scholar] [CrossRef] [Green Version]
- Dredge, J.; Fairchild, I.; Harrison, R.; Gunn, J. Processes affecting aerosol concentrations in Gough’s Cave, Cheddar Gorge, UK. Cave Karst Sci. 2014, 41, 42–47. [Google Scholar]
- Wang, W.; Ma, X.; Ma, Y.; Mao, L.; Wu, F.; Ma, X.; An, L.; Feng, H. Seasonal dynamics of airborne fungi in different caves of the Mogao Grottoes, Dunhuang, China. Int. Biodeter. Biodegr. 2010, 64, 461–466. [Google Scholar] [CrossRef]
- Wojkowski, J.; Andreychouk, V.; Fraczek, K. Airborne microorganisms of hypogenic maze caves based on the example of the Zoloushka Cave, Ukraine-Moldova. Rocz. Och. Śr. 2019, 21, 1116–1135. [Google Scholar]
- Leplat, J.; Francois, A.; Touron, S.; Frouin, M.; Portais, J.; Bousta, F. Aerobiological behavior of Paleolithic rock art sites in Dordogne (France): A comparative study in protected sites ranging from rock shelters to caves, with and without public access. Aerobiologia 2020, 36, 355–374. [Google Scholar] [CrossRef]
- Lee, K.; Park, S.; Kim, C. Effects of radon on soil microbial community and their growth. Environ. Eng. Res. 2020, 25, 29–35. [Google Scholar] [CrossRef] [Green Version]
- Gregorič, A.; Vaupotič, J.; Šebela, S. The role of cave ventilation in governing cave air temperature and radon levels (Postojna Cave, Slovenia). Int. J. Climatol. 2014, 34, 1488–1500. [Google Scholar] [CrossRef]
- Akira, S.; Uematsu, S.; Takeuchi, O. Pathogen recognition and innate immunity. Cell 2006, 124, 783–801. [Google Scholar] [CrossRef] [Green Version]
- Singh, A.; Singh, A.; Pandit, T. Respiratory diseases among agricultural industry workers in India: A cross-sectional epidemiological study. Ann. Agr. Env. Med. 1999, 6, 115–126. [Google Scholar]
- Krysinska-Traczyk, E.; Pande, B.; Skorska, C.; Sitkowska, J.; Prazmo, Z.; Cholewa, G.; Dutkiewicz, J. Exposure of Indian agricultural workers to airborne microorganisms, dust and endotoxin during handling of various plant products. Ann. Agr. Env. Med. 2005, 12, 269–275. [Google Scholar]
- Rylander, R. Health effects of organic dusts in the farm environment. Am. J. Ind. Med. 1986, 10, 206–240. [Google Scholar]
- Rylander, R. The role of endotoxin for reactions after exposure to cotton dust. Am. J. Ind. Med. 1987, 12, 687–697. [Google Scholar] [CrossRef]
- Malmros, P.; Sigsgaard, T.; Bach, B. Occupational health problems due to garbage sorting. Waste Manag. Res. 1992, 10, 227–234. [Google Scholar] [CrossRef]
- Gorny, R.; Dutkiewicz, J. Bacterial and fungal aerosols in indoor environment in Central and Eastern European countries. Ann. Agr. Env. Med. 2002, 9, 17–23. [Google Scholar]
- Laitinen, S.; Kangas, J.; Husman, K.; Susitaival, P. Evaluation of exposure to airborne bacterial endotoxins and peptidoglycans in selected work environments. Ann. Agr. Env. Med. 2001, 8, 213–219. [Google Scholar]
- DECOS. Endotoxins: Health Based Recommended Exposure Limit; A Report of the Health Council of the Netherlands; Health Council of the Netherlands: Rijswijk, The Netherlands, 1998. [Google Scholar]
- Becker, K.; Pagnier, I.; Schuhen, B.; Wenzelburger, F.; Friedrich, A.; Kipp, F.; Peters, G.; von Eiff, C. Does nasal cocolonization by methicillin-resistant coagulase-negative staphylococci and methicillin-susceptible Staphylococcus aureus strains occur frequently enough to represent a risk of false-positive methicillin-resistant S. aureus determinations by molecular methods? J. Clin. Microbiol. 2006, 44, 229–231. [Google Scholar] [PubMed] [Green Version]
- Coughenour, C.; Stevens, V.; Stetzenbach, L. An evaluation of methicillin-resistant Staphylococcus aureus survival on five environmental surfaces. Microb. Drug Resist. 2011, 17, 457–461. [Google Scholar] [CrossRef] [PubMed]
- Zarpellon, M.; Gales, A.; Sasaki, A.; Selhorst, G.; Menegucci, T.; Cardoso, C.; Garcia, L.; Tognim, M. Survival of vancomycin-intermediate Staphylococcus aureus on hospital surfaces. J. Hosp. Infect. 2015, 90, 347–350. [Google Scholar] [CrossRef] [PubMed]
- Neely, A.; Maley, M. Survival of enterococci and staphylococci on hospital fabrics and plastic. J. Clin. Microbiol. 2000, 38, 724–726. [Google Scholar] [CrossRef] [Green Version]
- Rich, M. Staphylococci in animals: Prevalence, identification and antimicrobial susceptibility, with an emphasis on methicillin-resistant Staphylococcus aureus. Brit. J. Biomed. Sci. 2005, 62, 98–105. [Google Scholar] [CrossRef]
- Becker, K.; Both, A.; Weisselberg, S.; Heilmann, C.; Rohde, H. Emergence of coagulase-negative staphylococci. Expert Rev. Anti-Infect. Ther. 2020, 18, 349–366. [Google Scholar] [CrossRef]
- Kaspar, U.; Kriegeskorte, A.; Schubert, T.; Peters, G.; Rudack, C.; Pieper, D.; Wos-Oxley, M.; Becker, K. The culturome of the human nose habitats reveals individual bacterial fingerprint patterns. Environ. Microbiol. 2016, 18, 2130–2142. [Google Scholar] [CrossRef]
- Nguyen, T.; Park, M.; Otto, M. Host response to Staphylococcus epidermidis colonization and infections. Front. Cell. Infect. Microbiol. 2017, 7, 90. [Google Scholar] [CrossRef] [Green Version]
- Hovelius, B.; Mardh, P. Staphylococcus saprophyticus as a common cause of urinary-tract infections. Rev. Infect. Dis. 1984, 6, 328–337. [Google Scholar] [CrossRef] [PubMed]
- Tong, S.; Davis, J.; Eichenberger, E.; Holland, T.; Fowler, V. Staphylococcus aureus infections: Epidemiology, pathophysiology, clinical manifestations, and management. Clin. Microbiol. Rev. 2015, 28, 603–661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taylor, T.; Unakal, C. Staphylococcus aureus. Available online: https://www.ncbi.nlm.nih.gov/books/NBK441868/ (accessed on 15 March 2022).





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ATP | BG | LPS | Fungi | Bacteria | |
|---|---|---|---|---|---|
| ATP | 1.000 | 0.413 | 0.269 | 0.398 | 0.537 |
| BG | 0.413 | 1.000 | –0.010 | 0.863 | 0.580 |
| LPS | 0.269 | –0.010 | 1.000 | 0.188 | 0.044 |
| Fungi | 0.398 | 0.863 | 0.188 | 1.000 | 0.588 |
| Bacteria | 0.537 | 0.580 | 0.044 | 0.588 | 1.000 |
| Species | Site | Risk Group (Country) |
|---|---|---|
| S. arlettae | 1, 13 | – |
| S. aureus | 16 | 2 (US, NZ, BE, CA, EU, CH, UK) |
| S. chonii | 13, 14 | – |
| S. epidermidis | 4 | 2 (BE, DE, CH) |
| S. equorum | 5, 10, 13, 14, 15 | – |
| S. equorum ssp. equorum | 13 | – |
| S. haemolyticus | 10 | 2 (DE, CH) |
| S. pasteuri | 10 | 2 (DE, CH) |
| S. saprophyticus | 1, 3, 4, 6, 7, 13, 14, 15, 17 | 2 (BE, CA, DE, CH) |
| S. sciuri | 6, 11 | – |
| S. warneri | 2, 5, 10, 12 | – |
| S. xylosus | 1 | – |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mulec, J.; Skok, S.; Tomazin, R.; Letić, J.; Pliberšek, T.; Stopinšek, S.; Simčič, S. Long-Term Monitoring of Bioaerosols in an Environment without UV and Desiccation Stress, an Example from the Cave Postojnska Jama, Slovenia. Microorganisms 2023, 11, 809. https://doi.org/10.3390/microorganisms11030809
Mulec J, Skok S, Tomazin R, Letić J, Pliberšek T, Stopinšek S, Simčič S. Long-Term Monitoring of Bioaerosols in an Environment without UV and Desiccation Stress, an Example from the Cave Postojnska Jama, Slovenia. Microorganisms. 2023; 11(3):809. https://doi.org/10.3390/microorganisms11030809
Chicago/Turabian StyleMulec, Janez, Sara Skok, Rok Tomazin, Jasmina Letić, Tadej Pliberšek, Sanja Stopinšek, and Saša Simčič. 2023. "Long-Term Monitoring of Bioaerosols in an Environment without UV and Desiccation Stress, an Example from the Cave Postojnska Jama, Slovenia" Microorganisms 11, no. 3: 809. https://doi.org/10.3390/microorganisms11030809