
Microbial Consortium HJ-SH with Very High Degradation Efficiency of Phenanthrene


Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Culture Media
2.2. Isolation and Identification of a Phenanthrene-Degrading Microbial Consortium
2.3. Structural Comparison of Microbial Consortia HJ-SH and HJ-7
2.4. Phenanthrene Biodegradation by Single Strains and Consortia
2.5. Biodegradation of Biphenyl, Naphthalene and n-Hexadecane
2.6. Analysis of the Degradation Ratio
2.7. Statistical Analysis
3. Results
3.1. Isolation and Characterization of a Phenanthrene-Degrading Microbial Consortium
3.2. Structural Comparison of Microbial Consortia HJ-SH and HJ-7
3.3. Characterization of Phenanthrene Biodegradation
3.3.1. Phenanthrene Degradation by HJ-7 and HJ-SH
3.3.2. Phenanthrene Degradation by Single-Strain and HJ-SH
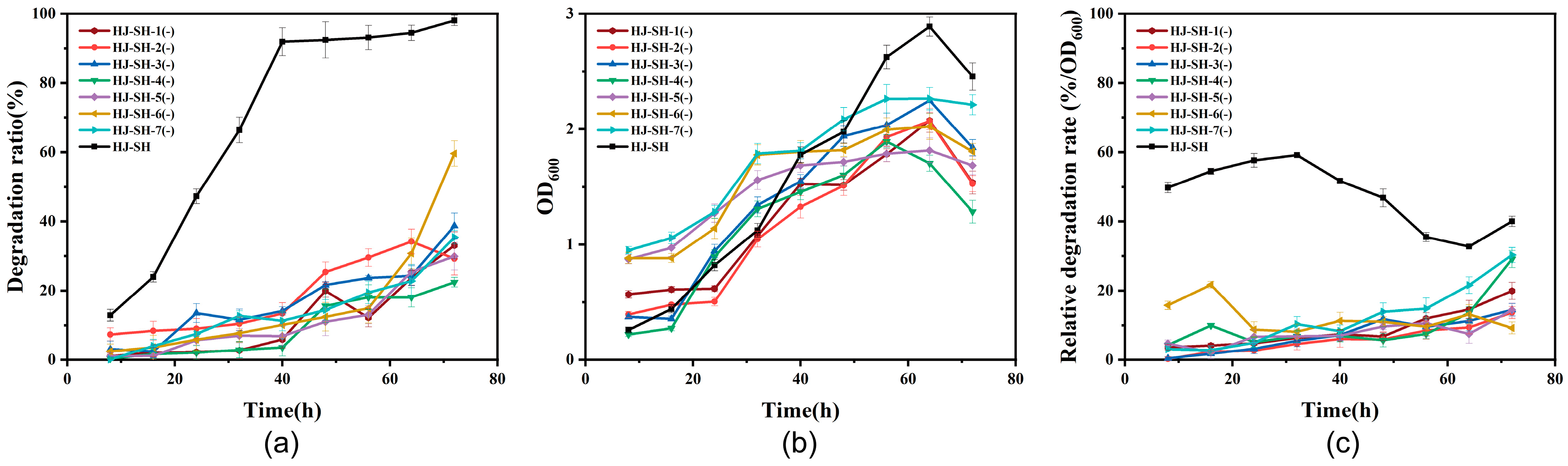
3.3.3. Phenanthrene Degradation by Six-Strains and HJ-SH
3.3.4. The Contribution of the Seven Strains
3.3.5. PHE Tolerance of the Consortium HJ-SH
3.4. Degradation of Other Organic Pollutants
4. Discussion

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain Name | The Ability to Degrade PHE | Reference |
|---|---|---|
| Coriolopsis byrsina | Degraded 99.90% of 20 mg/L PHE under in vitro conditions and 77.48% of 50 mg/L PHE under in vivo conditions. | [35] |
| Kocuria flava, Rhodococcus pyridinivorans | K. flava could degrade PHE with an efficiency of 55.13%, R. pyridinivorans exhibited 62.03% efficiency when the PHE concentration was 10 mg/L. | [65] |
| Serratia marcescens | Degraded 44% of 100 mg/L PHE after 3 days. | [66] |
| Pseudomonas sp. VB92 Bacillus sp. JK17 | Degraded 54.21% of 200 mg/L PHE in 10 days and 59.91% in 12 days. | [67] |
| Sphingobium xenophagum D43FB | Degraded 95% of 50 mg/L PHE after 5 days. | [60] |
| Cladosporium sp. CBMAI 1237 | Degraded 47% of 100 mg/L PHE and other PAHs after 21 days. | [61] |
| Pseudomonas sp. CGMCC2953-pK (engineered strain) | Degraded 98% of 100 mg/L PHE after 7 days. | [68] |
| Consortium YL (wild consortium) | Degraded 92.3% of 50 mg/L PHE after 21 days. | [69] |
| Halothermophilic consortium (engineered consortium) | Degraded 58% of 200 mg/L PHE after 42 days. | [70] |
| Artificial E. coli BL21 consortium (engineered consortium) | Degraded 90.66% of 100 mg/L PHE after 21 days. | [36] |
| Consortium HJ-SH (wild consortium) | Degraded 98.06% of 100 mg/L PHE after 3 days. | Present study |
| Consortium HJ-7 (artificial consortium without engineering) | Degraded 93.75% of 100 mg/L PHE after 3 days. | Present study |
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Li, H.; Qu, R.; Li, C.; Guo, W.; Han, X.; He, F.; Ma, Y.; Xing, B. Selective removal of polycyclic aromatic hydrocarbons (PAHs) from soil washing effluents using biochars produced at different pyrolytic temperatures. Bioresour. Technol. 2014, 163, 193–198. [Google Scholar] [CrossRef] [PubMed]
- Hadibarata, T.; Khudhair, A.B.; Kristanti, R.A.; Kamyab, H. Biodegradation of pyrene by Candida sp. S1 under high salinity conditions. Bioprocess Biosyst. Eng. 2017, 40, 1411–1418. [Google Scholar] [CrossRef] [PubMed]
- Gupta, G.; Kumar, V.; Pal, A.K. Biodegradation of Polycyclic Aromatic Hydrocarbons by Microbial Consortium: A Distinctive Approach for Decontamination of Soil. Soil Sediment Contam. 2016, 25, 597–623. [Google Scholar] [CrossRef]
- Moreda, J.M.; Arranz, A.; De Betono, S.F.; Cid, A.; Arranz, J.F. Chromatographic determination of aliphatic hydrocarbons and polyaromatic hydrocarbons (PAHs) in a sewage sludge. Sci. Total Environ. 1998, 220, 33–43. [Google Scholar] [CrossRef]
- Lipinska, A.; Wyszkowska, J.; Kucharski, J. Microbiological and Biochemical Activity in Soil Contaminated with Pyrene Subjected to Bioaugmentation. Water Air Soil Pollut. 2021, 232, 45. [Google Scholar] [CrossRef]
- Sutherland, J.B. Detoxification of polycyclic aromatic hydrocarbons by fungi. J. Ind. Microbiol. 1992, 9, 53–61. [Google Scholar] [CrossRef]
- Prak, D.J.L.; Pritchard, P.H. Solubilization of polycyclic aromatic hydrocarbon mixtures in micellar nonionic surfactant solutions. Water Res. 2002, 36, 3463–3472. [Google Scholar] [CrossRef]
- Seo, J.-S.; Keum, Y.-S.; Li, Q.X. Bacterial Degradation of Aromatic Compounds. Int. J. Environ. Res. Public Health 2009, 6, 278–309. [Google Scholar] [CrossRef]
- Singh, P.; Tiwary, B.N. Optimization of conditions for polycyclic aromatic hydrocarbons (PAHs) degradation by Pseudomonas stutzeri P2 isolated from Chirimiri coal mines. Biocatal. Agric. Biotechnol. 2017, 10, 20–29. [Google Scholar] [CrossRef]
- Wang, X.S.; Chen, M.Q.; Zheng, X. Polycyclic aromatic hydrocarbons (PAHs) in Xuzhou urban street dust: Concentration and sources. Environ. Earth Sci. 2017, 76, 576. [Google Scholar] [CrossRef]
- Lara-Moreno, A.; Morillo, E.; Merchan, F.; Villaverde, J. A comprehensive feasibility study of effectiveness and environmental impact of PAH bioremediation using an indigenous microbial degrader consortium and a novel strain Stenotrophomonas maltophilia CPHE1 isolated from an industrial polluted soil. J. Environ. Manag. 2021, 289, 112512. [Google Scholar] [CrossRef] [PubMed]
- Bellas, J.; Saco-Alvarez, L.; Nieto, O.; Beiras, R. Ecotoxicological evaluation of polycyclic aromatic hydrocarbons using marine invertebrate embryo-larval bioassays. Mar. Pollut. Bull. 2008, 57, 493–502. [Google Scholar] [CrossRef] [PubMed]
- Bekki, K.; Takigami, H.; Suzuki, G.; Tang, N.; Hayakawa, K. Evaluation of Toxic Activities of Polycyclic Aromatic Hydrocarbon Derivatives Using In Vitro Bioassays. J. Health Sci. 2009, 55, 601–610. [Google Scholar] [CrossRef]
- O’Connor, D.; Hou, D.; Ok, Y.S.; Mulder, J.; Duan, L.; Wu, Q.; Wang, S.; Tack, F.M.G.; Rinklebe, J. Mercury speciation, transformation, and transportation in soils, atmospheric flux, and implications for risk management: A critical review. Environ. Int. 2019, 126, 747–761. [Google Scholar] [CrossRef]
- Laothamteep, N.; Kawano, H.; Vejarano, F.; Suzuki-Minakuchi, C.; Shintani, M.; Nojiri, H.; Pinyakong, O. Effects of environmental factors and coexisting substrates on PAH degradation and transcriptomic responses of the defined bacterial consortium OPK. Environ. Pollut. 2021, 277, 116769. [Google Scholar] [CrossRef]
- Trapido, M. Polycyclic aromatic hydrocarbons in Estonian soil: Contamination and profiles. Environ. Pollut. 1999, 105, 67–74. [Google Scholar] [CrossRef]
- Li, X.; Peng, D.; Zhang, Y.; Ju, D.; Guan, C. Klebsiella sp. PD3, a phenanthrene (PHE)-degrading strain with plant growth promoting properties enhances the PHE degradation and stress tolerance in rice plants. Ecotoxicol. Environ. Saf. 2020, 201, 110804. [Google Scholar] [CrossRef]
- Vasudevan, V.; Gayathri, K.V.; Krishnan, M.E.G. Bioremediation of a pentacyclic PAH, Dibenz(a,h)Anthracene- A long road to trip with bacteria, fungi, autotrophic eukaryotes and surprises. Chemosphere 2018, 202, 387–399. [Google Scholar] [CrossRef]
- Wang, P.; Zhang, Y.; Jin, J.; Wang, T.; Wang, J.; Jiang, B. A high-efficiency phenanthrene-degrading Diaphorobacter sp. isolated from PAH-contaminated river sediment. Sci. Total Environ. 2020, 746, 140455. [Google Scholar] [CrossRef]
- Corgie, S.C.; Beguiristain, T.; Leyval, C. Differential composition of bacterial communities as influenced by phenanthrene and dibenzo a,h anthracene in the rhizosphere of ryegrass (Lolium perenne L.). Biodegradation 2006, 17, 511–521. [Google Scholar] [CrossRef]
- Jaafarzadeh, N.; Jorfi, S.; Mirzaiy, A.; Ghaedrahmat, Z. Bovine Manure as a Rich Source for Isolation of Halo-Tolerant Bacterial Strains Capable of PAHs Biodegradation in Slurry Bioreactor. Biointerface Res. Appl. Chem. 2021, 11, 14964–14973. [Google Scholar] [CrossRef]
- Mijaylova Nacheva, P.; Esquivel Sotelo, A. Removal of naphthalene and phenanthrene using aerobic membrane bioreactor. Biodegradation 2016, 27, 83–93. [Google Scholar] [CrossRef] [PubMed]
- Yang, K.; Wang, X.; Zhu, L.; Xing, B. Competitive sorption of pyrene, phenanthrene, and naphthalene on multiwalled carbon nanotubes. Environ. Sci. Technol. 2006, 40, 5804–5810. [Google Scholar] [CrossRef] [PubMed]
- Kuppusamy, S.; Thavamani, P.; Venkateswarlu, K.; Lee, Y.B.; Naidu, R.; Megharaj, M. Remediation approaches for polycyclic aromatic hydrocarbons (PAHs) contaminated soils: Technological constraints, emerging trends and future directions. Chemosphere 2017, 168, 944–968. [Google Scholar] [CrossRef] [PubMed]
- Gitipour, S.; Sorial, G.A.; Ghasemi, S.; Bazyari, M. Treatment technologies for PAH-contaminated sites: A critical review. Environ. Monit. Assess. 2018, 190, 546. [Google Scholar] [CrossRef]
- Wolejko, E.; Wydro, U.; Loboda, T. The Ways to Increase Efficiency of Soil Bioremediation. Ecol. Chem. Eng. S-Chem. I Inz. Ekol. S 2016, 23, 155–174. [Google Scholar] [CrossRef]
- Dadrasnia, A.; Wei, K.S.C.; Shahsavari, N.; Azirun, M.S.; Ismail, S. Biosorption Potential of Bacillus salmalaya Strain 139SI for Removal of Cr(VI) from Aqueous Solution. Int. J. Environ. Res. Public Health 2015, 12, 15321–15338. [Google Scholar] [CrossRef]
- Zhao, H.-P.; Wu, Q.-S.; Wang, L.; Zhao, X.-T.; Gao, H.-W. Degradation of phenanthrene by bacterial strain isolated from soil in oil refinery fields in Shanghai China. J. Hazard. Mater. 2009, 164, 863–869. [Google Scholar] [CrossRef]
- Liao, X.; Luo, J.; Cassidy, D.P.; Li, Y.; Tao, H.; Zhao, Y. Biodegradation of phenanthrene at high concentrations by Acidovorax sp. JG5 and its functional genomic analysis. J. Chem. Technol. Biotechnol. 2021, 96, 3142–3151. [Google Scholar] [CrossRef]
- Moody, J.D.; Freeman, J.P.; Fu, P.P.; Cerniglia, C.E. Degradation of benzo a pyrene by Mycobacterium vanbaalenii PYR-1. Appl. Environ. Microbiol. 2004, 70, 340–345. [Google Scholar] [CrossRef]
- Goswami, L.; Manikandan, N.A.; Dolman, B.; Pakshirajan, K.; Pugazhenthi, G. Biological treatment of wastewater containing a mixture of polycyclic aromatic hydrocarbons using the oleaginous bacterium Rhodococcus opacus. J. Clean. Prod. 2018, 196, 1282–1291. [Google Scholar] [CrossRef]
- Chebbi, A.; Hentati, D.; Zaghden, H.; Baccar, N.; Rezgui, F.; Chalbi, M.; Sayadi, S.; Chamkha, M. Polycyclic aromatic hydrocarbon degradation and biosurfactant production by a newly isolated Pseudomonas sp. strain from used motor oil-contaminated soil. Int. Biodeterior. Biodegrad. 2017, 122, 128–140. [Google Scholar] [CrossRef]
- Medic, A.; Ljesevic, M.; Inui, H.; Beskoski, V.; Kojic, I.; Stojanovic, K.; Karaddzic, I. Efficient biodegradation of petroleum n-alkanes and polycyclic aromatic hydrocarbons by polyextremophilic Pseudomonas aeruginosa san ai with multidegradative capacity. RSC Adv. 2020, 10, 14060–14070. [Google Scholar] [CrossRef] [PubMed]
- Boonchan, S.; Britz, M.L.; Stanley, G.A. Degradation and mineralization of high-molecular-weight polycyclic aromatic hydrocarbons by defined fungal-bacterial cocultures. Appl. Environ. Microbiol. 2000, 66, 1007–1019. [Google Scholar] [CrossRef]
- Mehetre, G.T.; Dastager, S.G.; Dharne, M.S. Biodegradation of mixed polycyclic aromatic hydrocarbons by pure and mixed cultures of biosurfactant producing thermophilic and thermo-tolerant bacteria. Sci. Total Environ. 2019, 679, 52–60. [Google Scholar] [CrossRef]
- Zhang, G.; Yang, X.; Zhao, Z.; Xu, T.; Jia, X. Artificial Consortium of Three E. coli BL21 Strains with Synergistic Functional Modules for Complete Phenanthrene Degradation. ACS Synth. Biol. 2022, 11, 162–175. [Google Scholar] [CrossRef]
- Luo, C.; Hu, X.; Bao, M.; Sun, X.; Li, F.; Li, Y.; Liu, W.; Yang, Y. Efficient biodegradation of phenanthrene using Pseudomonas stutzeri LSH-PAH1 with the addition of sophorolipids: Alleviation of biotoxicity and cometabolism studies. Environ. Pollut. 2022, 301, 119011. [Google Scholar] [CrossRef]
- Zhang, Z.; Guo, H.; Sun, J.; Gong, X.; Wang, C.; Wang, H. Anaerobic phenanthrene biodegradation by a newly isolated sulfate-reducer, strain PheS1, and exploration of the biotransformation pathway. Sci. Total Environ. 2021, 797, 149148. [Google Scholar] [CrossRef]
- Xu, T.; Liu, T.; Jiang, D.; Yuan, Z.; Jia, X. Attainment and characterization of a microbial consortium that efficiently degrades biphenyl and related substances. Biochem. Eng. J. 2021, 173, 108073. [Google Scholar] [CrossRef]
- Bankole, P.O.; Semple, K.T.; Jeon, B.-H.; Govindwar, S.P. Biodegradation of fluorene by the newly isolated marine-derived fungus, Mucor irregularis strain bpo1 using response surface methodology. Ecotoxicol. Environ. Saf. 2021, 208, 111619. [Google Scholar] [CrossRef]
- Ahmed, R.Z.; Ahmed, N. Effect of yeast extract on fluoranthene degradation and aromatic ring dioxygenase expressing bacterial community structure of a fluoranthene degrading bacterial consortium. Int. Biodeterior. Biodegrad. 2014, 88, 56–61. [Google Scholar] [CrossRef]
- Wang, H.; Lou, J.; Gu, H.; Luo, X.; Yang, L.; Wu, L.; Liu, Y.; Wu, J.; Xu, J. Efficient biodegradation of phenanthrene by a novel strain Massilia sp WF1 isolated from a PAH-contaminated soil. Environ. Sci. Pollut. Res. 2016, 23, 13378–13388. [Google Scholar] [CrossRef]
- Xiao, K.; Wang, R.; Liu, C.; Wang, M.; Zhang, J.; Dong, W.; Pan, T. Influence of Triton X-100 and beta-cyclodextrin on the bioavailability and biodegradation of crystalline phenanthrene covered with biofilms. Process. Biochem. 2021, 102, 173–179. [Google Scholar] [CrossRef]
- Ma, J.; Xu, L.; Jia, L. Degradation of polycyclic aromatic hydrocarbons by Pseudomonas sp JM2 isolated from active sewage sludge of chemical plant. J. Environ. Sci. 2012, 24, 2141–2148. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Li, C.; Huang, L.; He, Y.; Zhao, T.; Han, B.; Jia, X. Combination of a crude oil-degrading bacterial consortium under the guidance of strain tolerance and a pilot-scale degradation test. Chin. J. Chem. Eng. 2017, 25, 1838–1846. [Google Scholar] [CrossRef]
- Steen, A.D.; Crits-Christoph, A.; Carini, P.; DeAngelis, K.M.; Fierer, N.; Lloyd, K.G.; Thrash, J.C. High proportions of bacteria and archaea across most biomes remain uncultured. Isme J. 2019, 13, 3126–3130. [Google Scholar] [CrossRef] [PubMed]
- Albertsen, M.; Hugenholtz, P.; Skarshewski, A.; Nielsen, K.L.; Tyson, G.W.; Nielsen, P.H. Genome sequences of rare, uncultured bacteria obtained by differential coverage binning of multiple metagenomes. Nat. Biotechnol. 2013, 31, 533–538. [Google Scholar] [CrossRef]
- Nayfach, S.; Shi, Z.J.; Seshadri, R.; Pollard, K.S.; Kyrpides, N.C. New insights from uncultivated genomes of the global human gut microbiome. Nature 2019, 568, 505–510. [Google Scholar] [CrossRef]
- Xie, Y.; Gu, Z.; Herath, H.M.S.K.; Gu, M.; He, C.; Wang, F.; Jiang, X.; Zhang, J.; Zhang, Y. Evaluation of bacterial biodegradation and accumulation of phenanthrene in the presence of humic acid. Chemosphere 2017, 184, 482–488. [Google Scholar] [CrossRef]
- Mishra, A.; Rathour, R.; Singh, R.; Kumari, T.; Thakur, I.S. Degradation and detoxification of phenanthrene by actinobacteriumZhihengliuellasp. ISTPL4. Environ. Sci. Pollut. Res. 2020, 27, 27256–27267. [Google Scholar] [CrossRef]
- Zhou, L.; Sheng, X.; Zhang, S.; Liu, J. Screening of a phenanthrene-degrading bacterium and its degradation conditions. Ying Yong Sheng Tai Xue Bao = J. Appl. Ecol. 2005, 16, 2399–2402. [Google Scholar]
- Volpe, A.; Del Moro, G.; Rossetti, S.; Tandoi, V.; Lopez, A. Enhanced bioremediation of methyl tert-butyl ether (MTBE) by microbial consortia obtained from contaminated aquifer material. Chemosphere 2009, 75, 149–155. [Google Scholar] [CrossRef]
- Zhang, Q.; He, J.; Tian, M.; Mao, Z.; Tang, L.; Zhang, J.; Zhang, H. Enhancement of methane production from cassava residues by biological pretreatment using a constructed microbial consortium. Bioresour. Technol. 2011, 102, 8899–8906. [Google Scholar] [CrossRef]
- Pedetta, A.; Pouyte, K.; Seitz, M.K.H.; Babay, P.A.; Espinosa, M.; Costagliola, M.; Studdert, C.A.; Peressutti, S.R. Phenanthrene degradation and strategies to improve its bioavailability to microorganisms isolated from brackish sediments. Int. Biodeterior. Biodegrad. 2013, 84, 161–167. [Google Scholar] [CrossRef]
- Sangkharak, K.; Choonut, A.; Rakkan, T.; Prasertsan, P. The Degradation of Phenanthrene, Pyrene, and Fluoranthene and Its Conversion into Medium-Chain-Length Polyhydroxyalkanoate by Novel Polycyclic Aromatic Hydrocarbon-Degrading Bacteria. Curr. Microbiol. 2020, 77, 897–909. [Google Scholar] [CrossRef] [PubMed]
- Masakorala, K.; Yao, J.; Cai, M.; Chandankere, R.; Yuan, H.; Chen, H. Isolation and characterization of a novel phenanthrene (PHE) degrading strain Psuedomonas sp. USTB-RU from petroleum contaminated soil. J. Hazard. Mater. 2013, 263, 493–500. [Google Scholar] [CrossRef] [PubMed]
- Marcoux, J.; Deziel, E.; Villemur, R.; Lepine, F.; Bisaillon, J.G.; Beaudet, R. Optimization of high-molecular-weight polycyclic aromatic hydrocarbons’ degradation in a two-liquid-phase bioreactor. J. Appl. Microbiol. 2000, 88, 655–662. [Google Scholar] [CrossRef] [PubMed]
- Kumar, M.; Leon, V.; De Sisto Materano, A.; Ilzins, O.A.; Galindo-Castro, I.; Fuenmayor, S.L. Polycyclic aromatic hydrocarbon degradation by biosurfactant-producing Pseudomonas sp IR1. Z. Fur Naturforschung Sect. C-A J. Biosci. 2006, 61, 203–212. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.; Zhang, Y.; Hesham, A.E.-L.; Liu, R.; Yang, M. Cell surface properties of five polycyclic aromatic compound-degrading yeast strains. Appl. Microbiol. Biotechnol. 2010, 86, 1933–1939. [Google Scholar] [CrossRef]
- Gran-Scheuch, A.; Fuentes, E.; Bravo, D.M.; Cristobal Jimenez, J.; Perez-Donoso, J.M. Isolation and Characterization of Phenanthrene Degrading Bacteria from Diesel Fuel-Contaminated Antarctic Soils. Front. Microbiol. 2017, 8, 1634. [Google Scholar] [CrossRef]
- Birolli, W.G.; Santos, D.d.A.; Alvarenga, N.; Garcia, A.C.F.S.; Romao, L.P.C.; Porto, A.L.M. Biodegradation of anthracene and several PAHs by the marine-derived fungus Cladosporium sp. CBMAI 1237. Mar. Pollut. Bull. 2018, 129, 525–533. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Chen, R.F.; Shiaris, M.P. Metabolism of naphthalene, fluorene, and phenanthrene: Preliminary characterization of a cloned. J. Bacteriol. 1994, 176, 2158–2164. [Google Scholar] [CrossRef] [PubMed]
- Kiyohara, H. Phenanthrene-degrading phenotype of Alcaligenes faecalis AFK2. Appl. Environ. Microbiol. 1982, 43, 458–461. [Google Scholar] [CrossRef] [PubMed]
- Wei, H.; Wang, L.; Hassan, M.; Xie, B. Succession of the functional microbial communities and the metabolic functions in maize straw composting process. Bioresour. Technol. 2018, 256, 333–341. [Google Scholar] [CrossRef] [PubMed]
- Sakshi; Singh, S.K.; Haritash, A.K. Catabolic enzyme activities during biodegradation of three-ring PAHs by novel DTU-1Y and DTU-7P strains isolated from petroleum-contaminated soil. Arch. Microbiol. 2021, 203, 3101–3110. [Google Scholar] [CrossRef] [PubMed]
- Pandey, A.K.; Chaudhary, P.; Singh, S.B.; Arora, A.; Kumar, K.; Chaudhry, S.; Nain, L. Deciphering the traits associated with PAH degradation by a novel Serratia marcesencs L-11 strain. J. Environ. Sci. Health Part A-Toxic/Hazard. Subst. Environ. Eng. 2012, 47, 755–765. [Google Scholar] [CrossRef]
- Xu, F.; Feng, G.; Ma, W.; Wang, J.; Li, M.; Wang, Y.; Hu, S. Study on the Screening and Degradation Characteristics of Highly Efficient Petroleum Degrading Bacteria. Biotechnol. Bull. 2010, 221–226. [Google Scholar]
- Zhou, Y.; Wei, J.; Shao, N.; Wei, D. Construction of a genetically engineered microorganism for phenanthrene biodegradation. J. Basic Microbiol. 2013, 53, 188–194. [Google Scholar] [CrossRef]
- Lin, Y.; Cai, L.-X. PAH-degrading microbial consortium and its pyrene-degrading plasmids from mangrove sediment samples in Huian, China. Mar. Pollut. Bull. 2008, 57, 703–706. [Google Scholar] [CrossRef]
- Pugazhendi, A.; Qari, H.; Basahi, J.M.A.-B.; Godon, J.J.; Dhavamani, J. Role of a halothermophilic bacterial consortium for the biodegradation of PAHs and the treatment of petroleum wastewater at extreme conditions. Int. Biodeterior. Biodegrad. 2017, 121, 44–54. [Google Scholar] [CrossRef]







| Strain Combination | Source | Name |
|---|---|---|
| Microbial consortium | PHE-contaminated soil | HJ-SH |
| Single-strain pure cultures | Seven single-strains isolated from HJ-SH | SH-1, SH-2, SH-3, SH-4, SH-5, SH-6, SH-7 |
| Six-strain co-cultures | Remove SH-1 from HJ-SH | HJ-SH-1 (-) |
| Remove SH-2 from HJ-SH | HJ-SH-2 (-) | |
| Remove SH-3 from HJ-SH | HJ-SH-3 (-) | |
| Remove SH-4 from HJ-SH | HJ-SH-4(-) | |
| Remove SH-5 from HJ-SH | HJ-SH-5(-) | |
| Remove SH-6 from HJ-SH | HJ-SH-6(-) | |
| Remove SH-7 from HJ-SH | HJ-SH-7(-) | |
| Seven-strain co-culture | Seven strains isolated from HJ-SH | HJ-7 |
| Strains | SH-4 | SH-7 | SH-3 | SH-1 | SH-6 | SH-5 | SH-2 |
|---|---|---|---|---|---|---|---|
| Contribution of HJ-SH | 0.500 | 0.342 | 0.271 | 0.238 | 0.197 | 0.181 | 0.177 |
| Contribution of HJ-7 | 0.530 | 0.367 | 0.290 | 0.255 | 0.222 | 0.193 | 0.189 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, R.; Zhao, Z.; Xu, T.; Jia, X. Microbial Consortium HJ-SH with Very High Degradation Efficiency of Phenanthrene. Microorganisms 2023, 11, 2383. https://doi.org/10.3390/microorganisms11102383
Chen R, Zhao Z, Xu T, Jia X. Microbial Consortium HJ-SH with Very High Degradation Efficiency of Phenanthrene. Microorganisms. 2023; 11(10):2383. https://doi.org/10.3390/microorganisms11102383
Chicago/Turabian StyleChen, Rui, Zhenhua Zhao, Tao Xu, and Xiaoqiang Jia. 2023. "Microbial Consortium HJ-SH with Very High Degradation Efficiency of Phenanthrene" Microorganisms 11, no. 10: 2383. https://doi.org/10.3390/microorganisms11102383