
The Influence of Hydrodynamic Conditions in a Laboratory-Scale Bioreactor on Pseudomonas aeruginosa Metabolite Production
 , , , , , and
, , , , , and 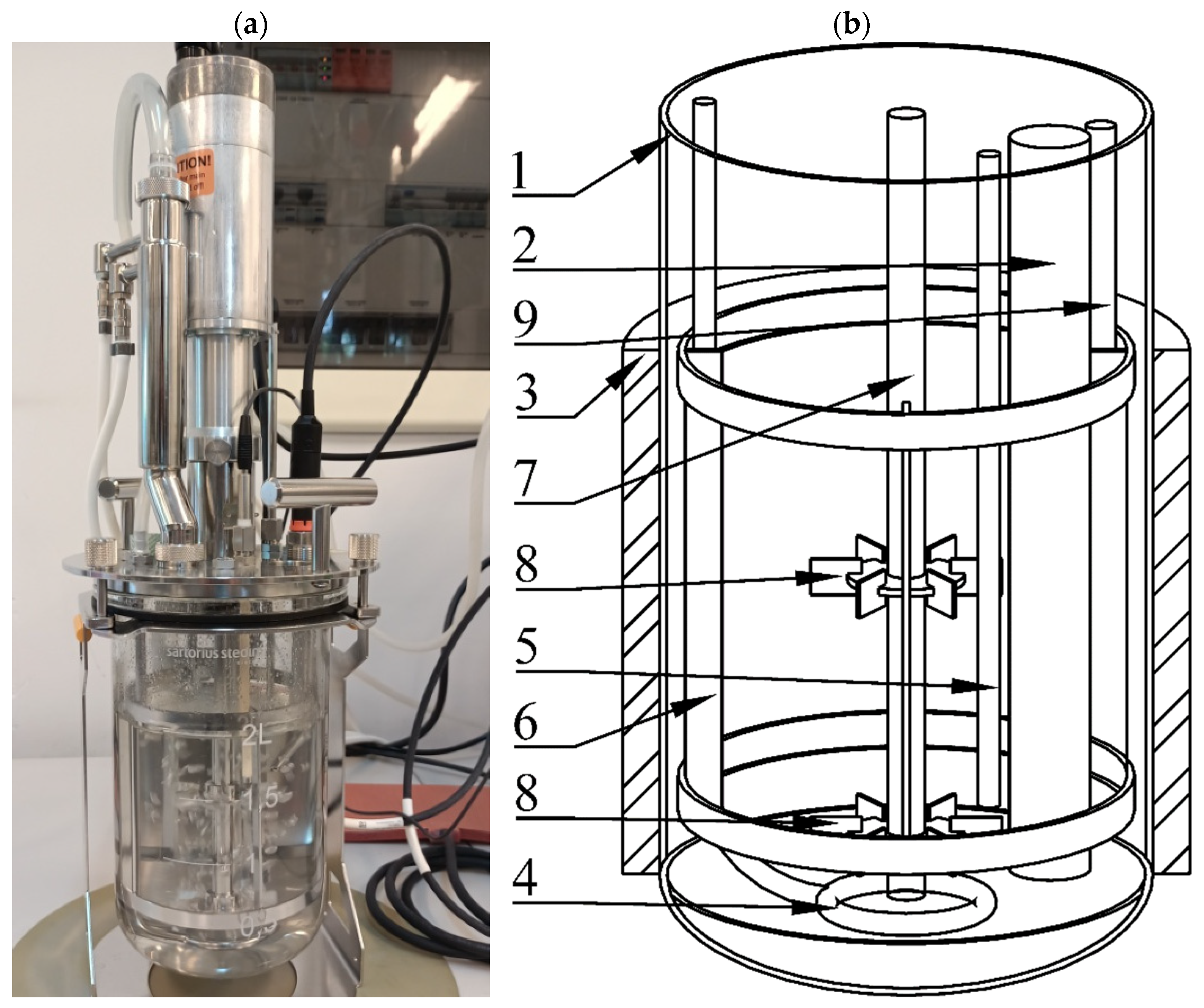{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Highlights
- This study demonstrated that the process production of Pseudomonas aeruginosa metabolites might be successfully carried out by using a batch bioreactor. This process is dependent on the hydrodynamic conditions. Therefore, we tested and analyzed the fluid behaviour for gassed and ungassed conditions with the application of the various types of hydrodynamic criteria (e.g., power consumption, mixing time, mixing energy, the volumetric gas-liquid mass transfer coefficient). The obtained hydrodynamic parameters might be successfully used in the bioprocessing of the different substances with the application of the tested bioreactor.
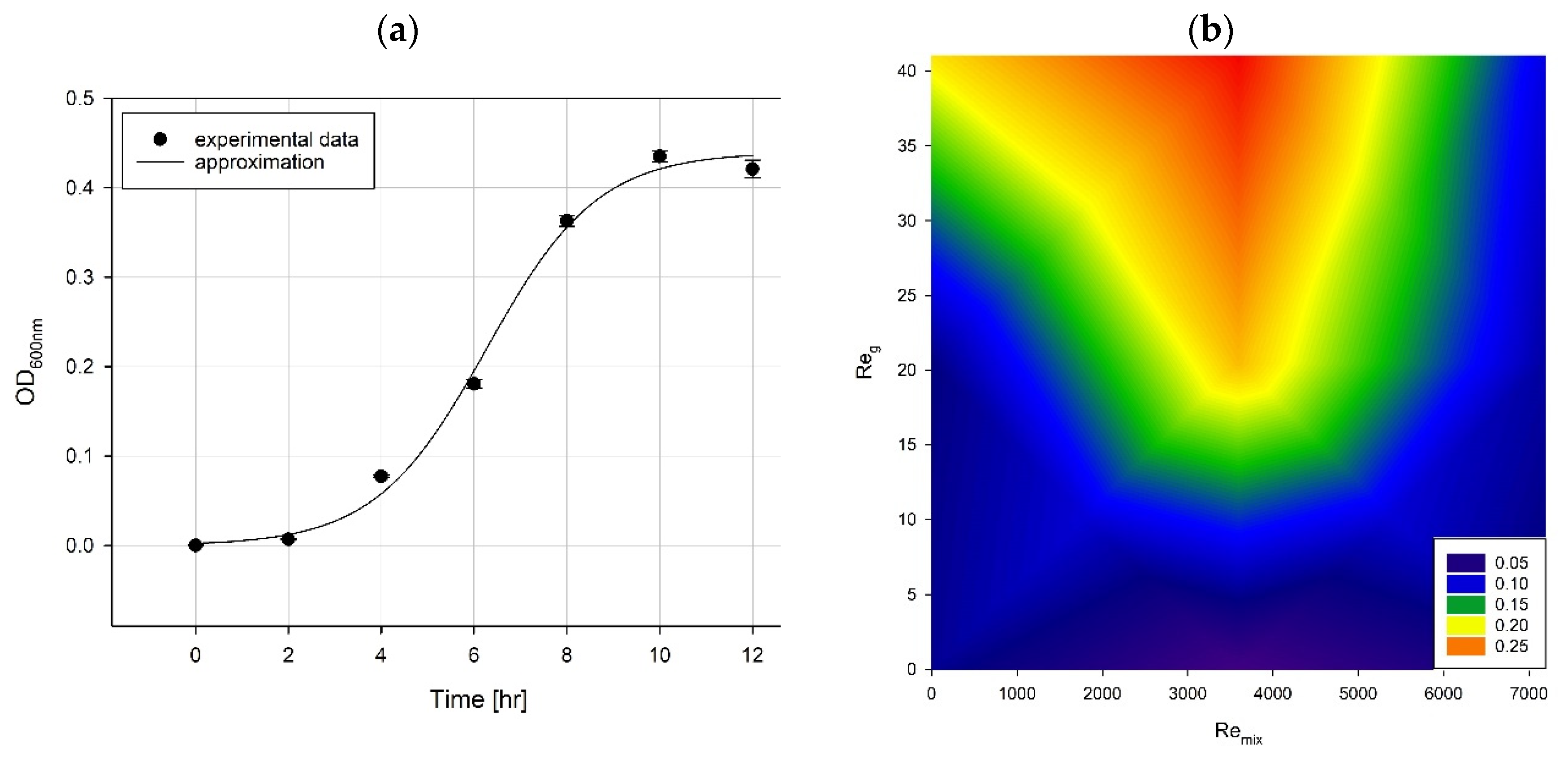
- The growth and metabolite production of Pseudomonas aeruginosa was analyzed under selected hydrodynamic conditions. We found that the highest oxygen mass transfer is connected with the most intensive pyocyanin production. The rhamnolipids were produced under low mixing conditions and moderate oxygen mass transfer.
- The obtained data showed that the process production of Pseudomonas aeruginosa metabolites is possible with the use of a batch bioreactor. It should be emphasized that the hydrodynamic conditions in the used mixing system have strongly influenced the obtained product. Therefore, the hydrodynamic analysis for bioprocessing or process production of bioproducts with the application of the mixing systems is required.
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Setup
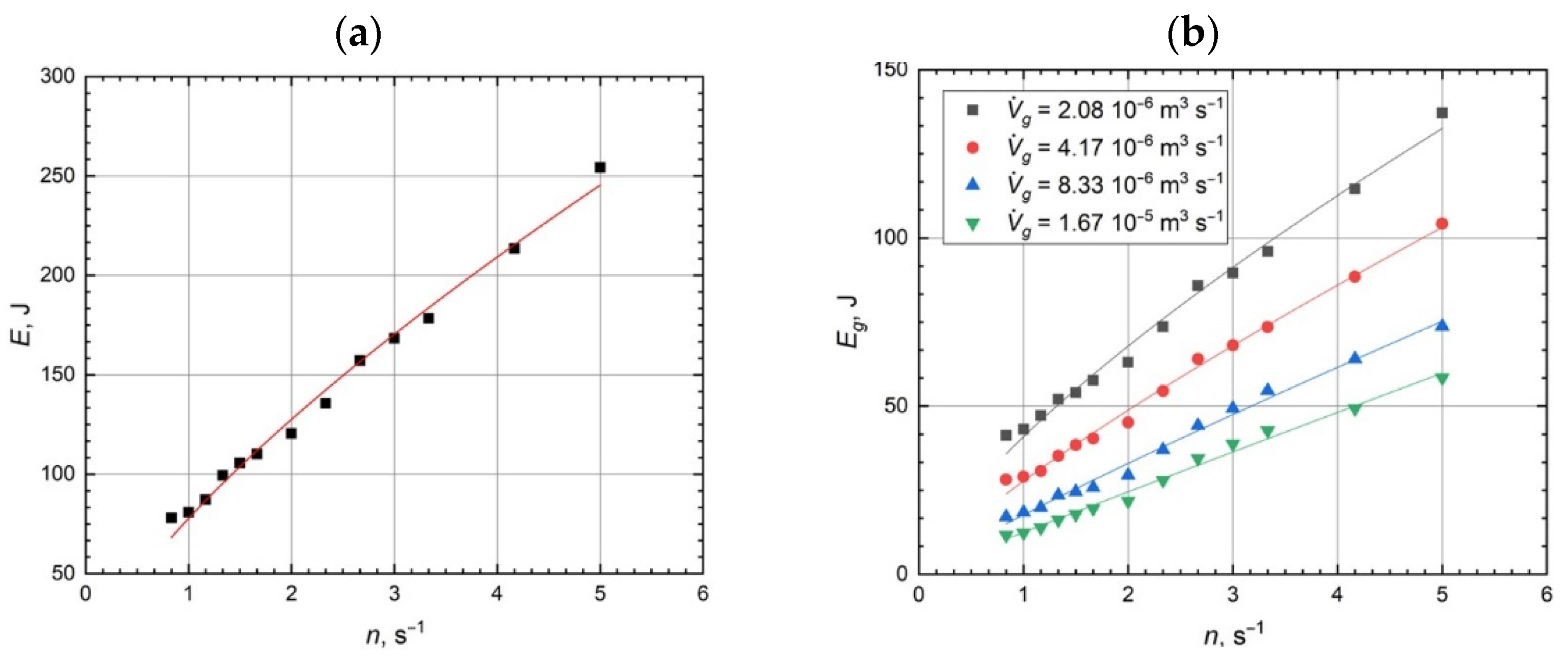
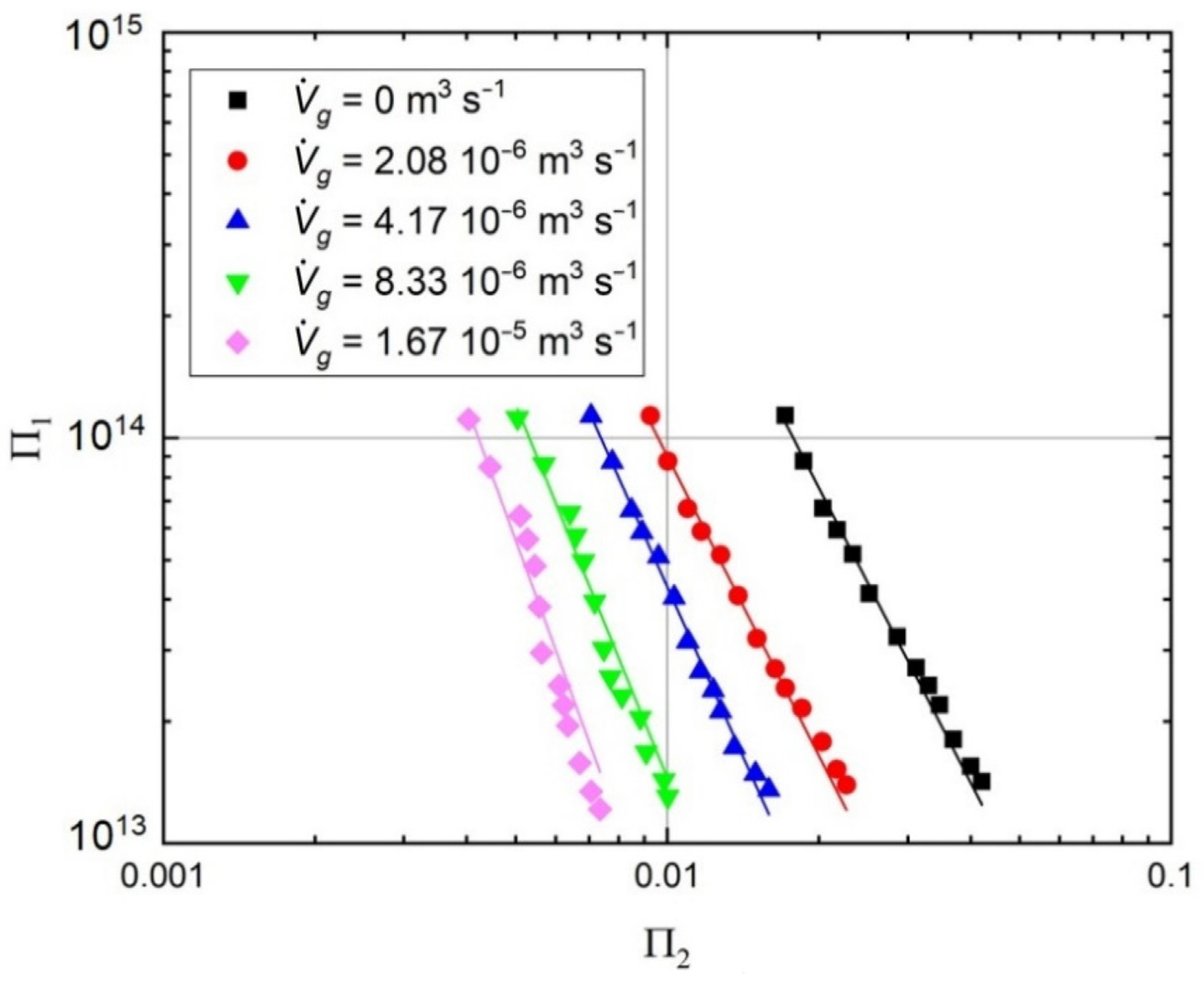
2.2. Hydrodynamic Conditions, Mixing Efficiency, and Mass Transfer Characteristics for the Tested Bioreactor
2.3. Growth and Viability Measurements
2.4. Pyocyanin and Rhamnolipid Quantification
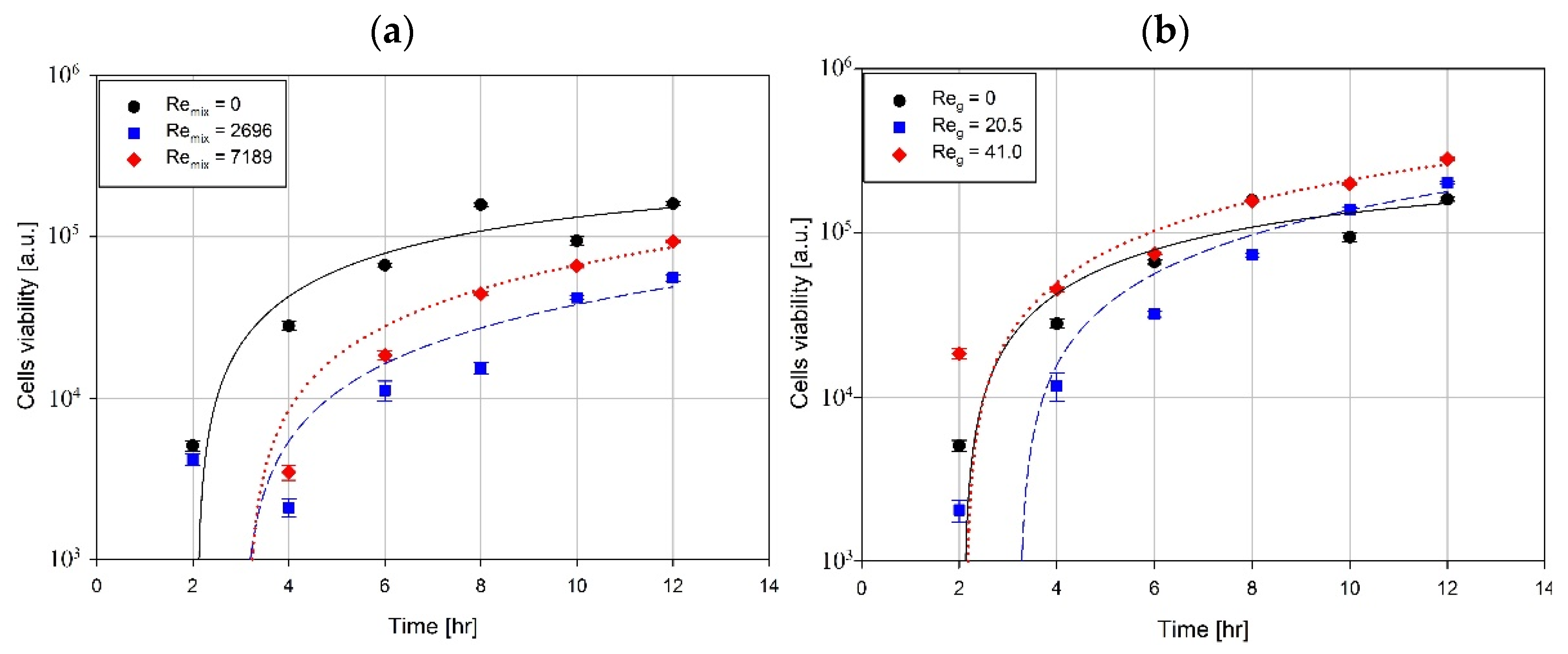
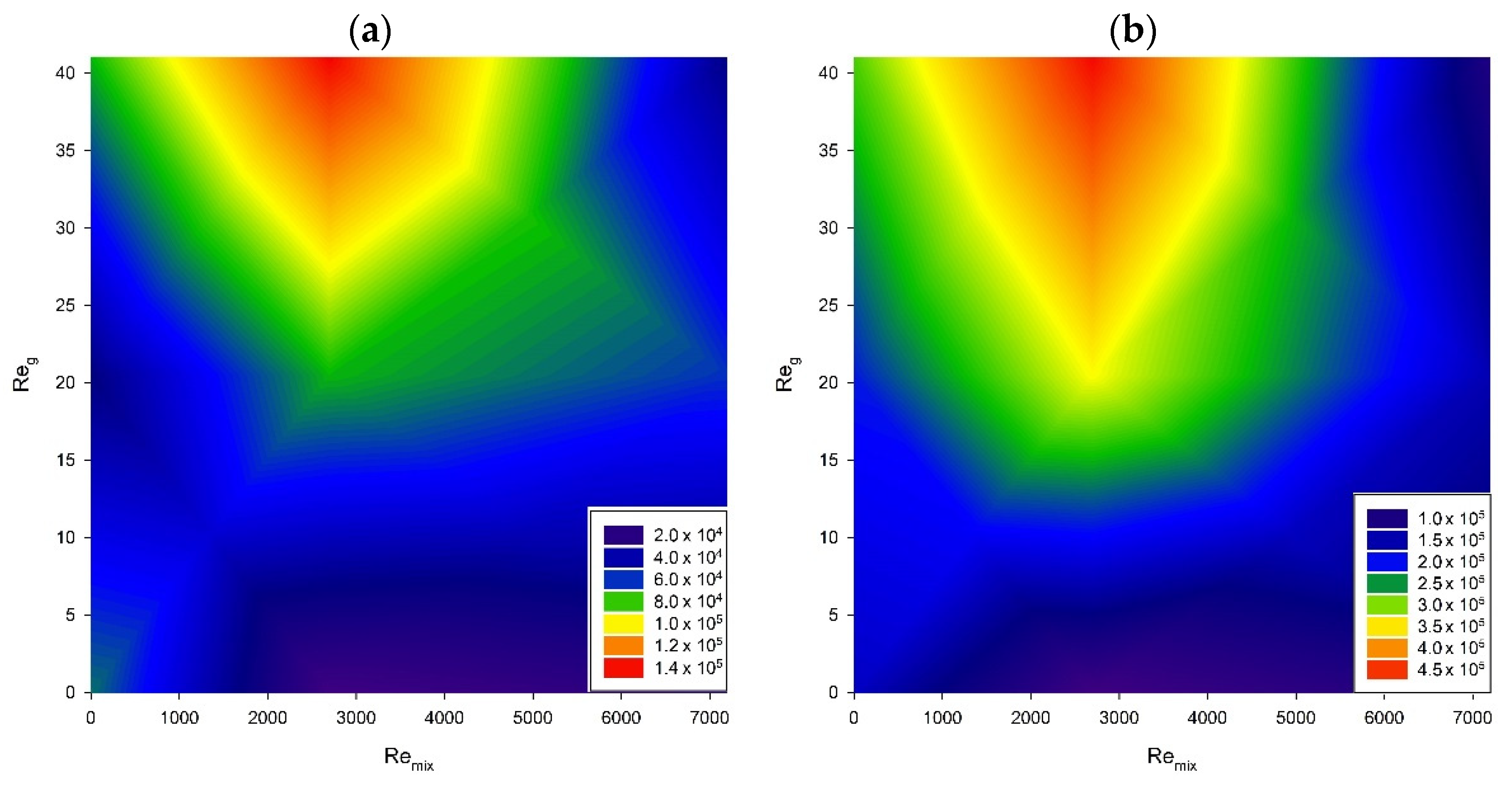
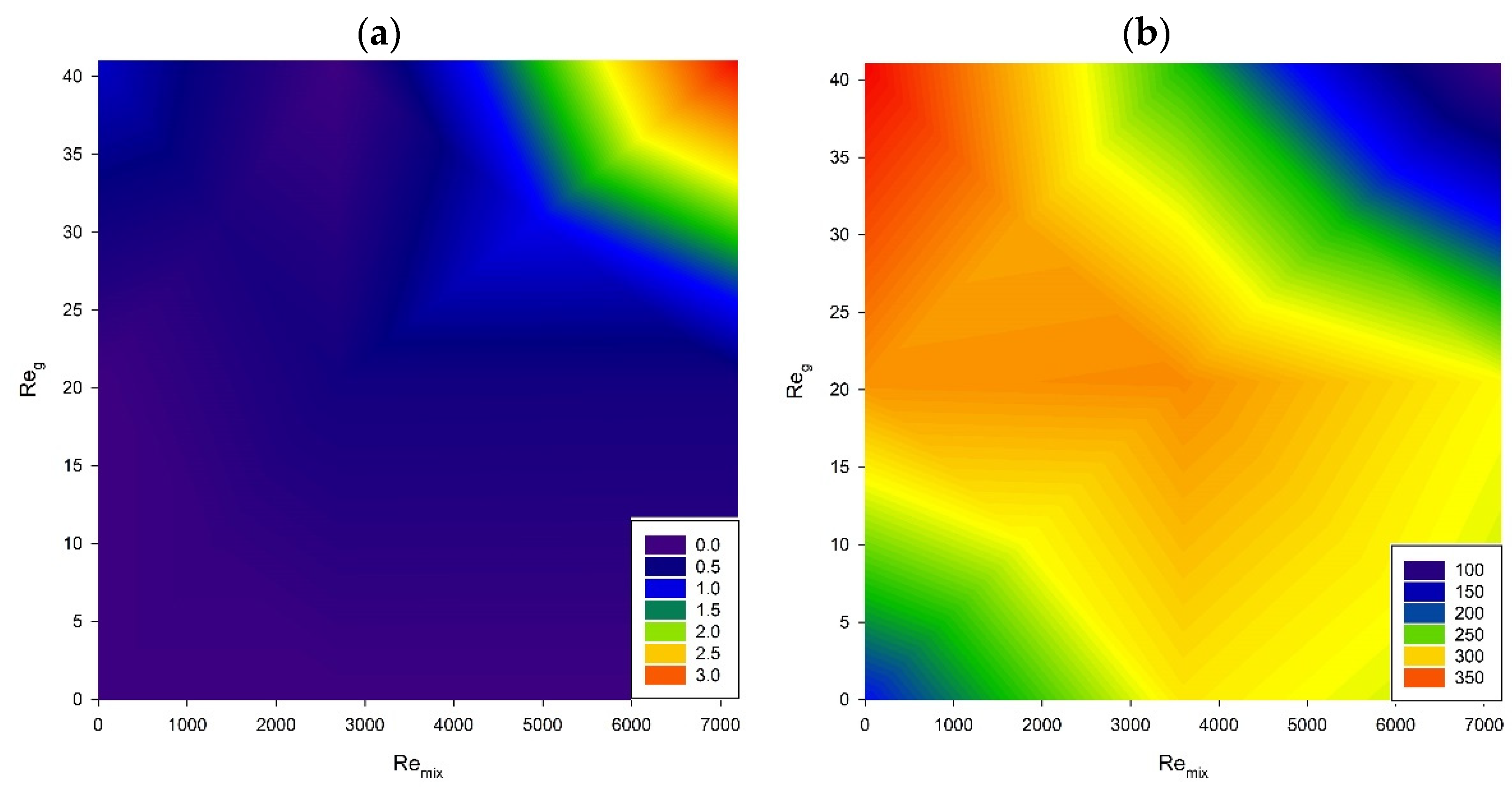
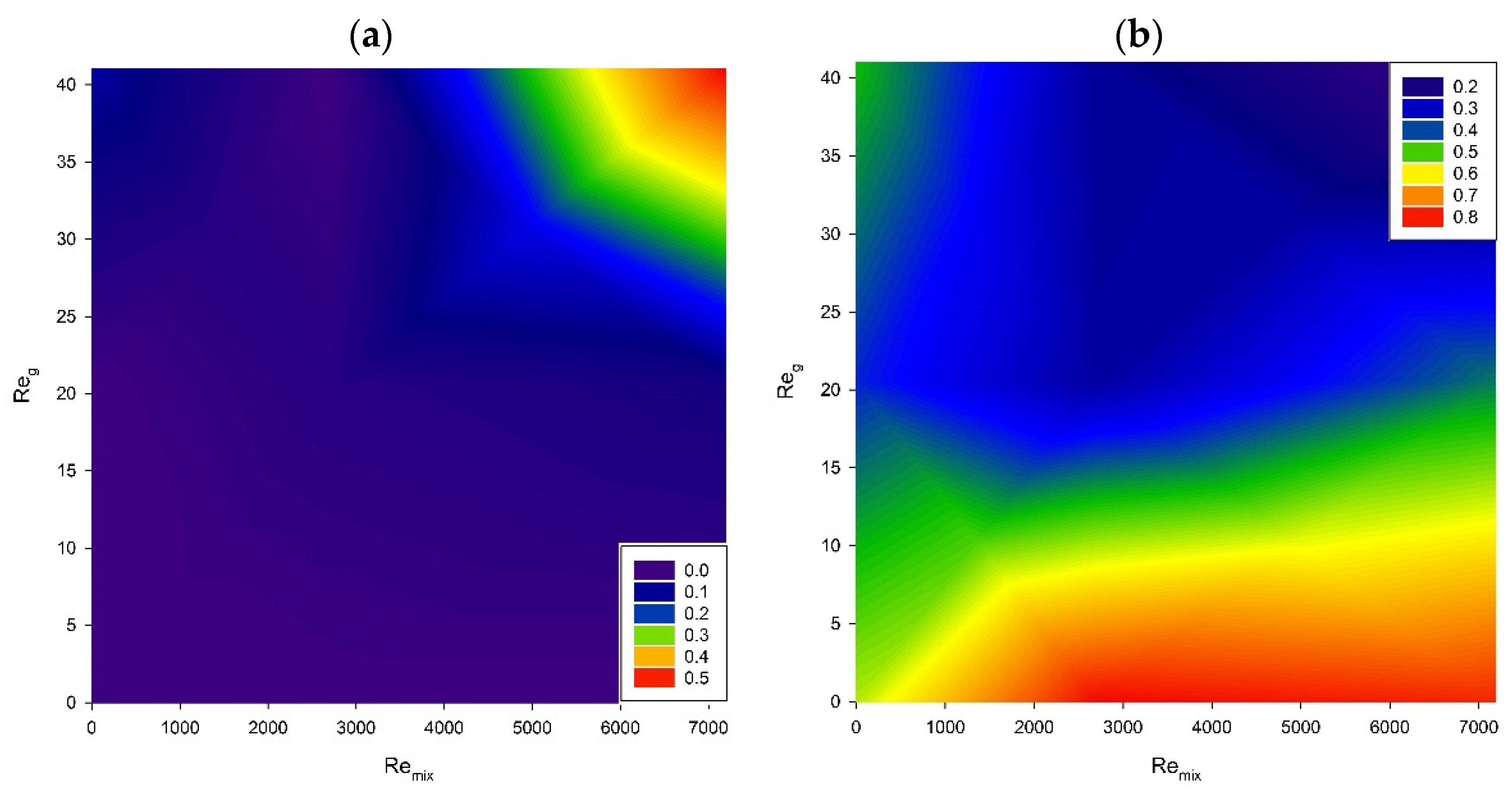
3. Results and Discussion
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Diano, A.; Bekker-Jensen, S.; Dynesen, J.; Nielsen, J. Polyol synthesis in Aspergillus niger: Influence of oxygen availability, carbon and nitrogen sources on the metabolism. Biotechnol. Bioeng. 2006, 94, 899–908. [Google Scholar] [CrossRef] [PubMed]
- Talwalkar, A.; Kailasapathy, K. Metabolic and Biochemical Responses of Probiotic Bacteria to Oxygen. J. Dairy Sci. 2003, 86, 2537–2546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salmon, J.-M. Interactions between yeast, oxygen and polyphenols during alcoholic fermentations: Practical implications. LWT Food Sci. Technol. 2006, 39, 959–965. [Google Scholar] [CrossRef]
- Li, A.; Pfelzer, N.; Zuijderwijk, R.; Brickwedde, A.; van Zeijl, C.; Punt, P. Reduced by-product formation and modified oxygen availability improve itaconic acid production in Aspergillus niger. Appl. Microbiol. Biotechnol. 2013, 97, 3901–3911. [Google Scholar] [CrossRef]
- Riedel, T.E.; Berelson, W.M.; Nealson, K.H.; Finkel, S.E. Oxygen Consumption Rates of Bacteria under Nutrient-Limited Conditions. Appl. Environ. Microbiol. 2013, 79, 4921–4931. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Ochoa, F.; Gomez, E. Bioreactor scale-up and oxygen transfer rate in microbial processes: An overview. Biotechnol. Adv. 2009, 27, 153–176. [Google Scholar] [CrossRef]
- Gómez Sánchez, C.E.; Martínez-Trujillo, A.; Aguilar Osorio, G. Oxygen transfer coefficient and the kinetic parameters of exo-polygalacturonase production by Aspergillus flavipes FP-500 in shake flasks and bioreactor. Lett. Appl. Microbiol. 2012, 55, 444–452. [Google Scholar] [CrossRef]
- Kordas, M.; Konopacki, M.; Grygorcewicz, B.; Augustyniak, A.; Musik, D.; Wójcik, K.; Jędrzejczak-Silicka, M.; Rakoczy, R. Hydrodynamics and Mass Transfer Analysis in BioFlow® Bioreactor Systems. Processes 2020, 8, 1311. [Google Scholar] [CrossRef]
- Rakoczy, R.; Lechowska, J.; Kordas, M.; Konopacki, M.; Fijałkowski, K.; Drozd, R. Effects of a rotating magnetic field on gas-liquid mass transfer coefficient. Chem. Eng. J. 2017, 327, 608–617. [Google Scholar] [CrossRef]
- Konopacki, M.; Kordas, M.; Fijałkowski, K.; Rakoczy, R. Computational fluid dynamics and experimental studies of a new mixing element in a static mixer as a heat exchanger. Chem. Process. Eng. 2015, 36, 59–72. [Google Scholar] [CrossRef]
- Soerjawinata, W.; Schlegel, K.; Fuchs, N.; Schüffler, A.; Schirmeister, T.; Ulber, R.; Kampeis, P. Applicability of a single-use bioreactor compared to a glass bioreactor for the fermentation of filamentous fungi and evaluation of the reproducibility of growth in pellet form. Eng. Life Sci. 2021, 21, 324–339. [Google Scholar] [CrossRef] [PubMed]
- Hupfeld, J.; Gorr, I.H.; Schwald, C.; Beaucamp, N.; Wiechmann, K.; Kuentzer, K.; Huss, R.; Rieger, B.; Neubauer, M.; Wegmeyer, H. Modulation of mesenchymal stromal cell characteristics by microcarrier culture in bioreactors. Biotechnol. Bioeng. 2014, 111, 2290–2302. [Google Scholar] [CrossRef] [PubMed]
- Wyrobnik, T.A.; Ducci, A.; Micheletti, M. Advances in human mesenchymal stromal cell-based therapies—Towards an integrated biological and engineering approach. Stem Cell Res. 2020, 47, 101888. [Google Scholar] [CrossRef] [PubMed]
- Hegner, R.; Koch, C.; Riechert, V.; Harnisch, F. Microbiome-based carboxylic acids production: From serum bottles to bioreactors. RSC Adv. 2017, 7, 15362–15371. [Google Scholar] [CrossRef] [Green Version]
- Rahman, P.K.S.M.; Pasirayi, G.; Auger, V.; Ali, Z. Development of a simple and low cost microbioreactor for high-throughput bioprocessing. Biotechnol. Lett. 2009, 31, 209–214. [Google Scholar] [CrossRef] [Green Version]
- Patil, S.; Nikam, M.; Patil, H.; Anokhina, T.; Kochetkov, V.; Chaudhari, A. Bioactive pigment production by Pseudomonas spp. MCC 3145: Statistical media optimization, biochemical characterization, fungicidal and DNA intercalation-based cytostatic activity. Process. Biochem. 2017, 58, 298–305. [Google Scholar] [CrossRef]
- Amani, H. Application of a Dynamic Method for the Volumetric Mass Transfer Coefficient Determination in the Scale-Up of Rhamnolipid Biosurfactant Production. J. Surfactants Deterg. 2018, 21, 827–833. [Google Scholar] [CrossRef]
- Martin-Arjol, I.; Llacuna, J.L.; Manresa, Á. Yield and kinetic constants estimation in the production of hydroxy fatty acids from oleic acid in a bioreactor by Pseudomonas aeruginosa 42A2. Appl. Microbiol. Biotechnol. 2014, 98, 9609–9621. [Google Scholar] [CrossRef]
- Mahato, R.P.; Kumar, S.; Singh, P. Optimization of Growth Conditions to Produce Sustainable Polyhydroxyalkanoate Bioplastic by Pseudomonas aeruginosa EO1. Front. Microbiol. 2021, 12, 711588. [Google Scholar] [CrossRef]
- Bisht, D.; Yadav, S.K.; Darmwal, N.S. Optimization of immobilization conditions by conventional and statistical strategies for alkaline lipase production by Pseudomonas aeruginosa mutant cells: Scale-up at bench-scale bioreactor level. Turkish J. Biol. 2013, 37, 392–404. [Google Scholar] [CrossRef]
- Panichikkal, A.F.; Prakasan, P.; Kizhakkepowathial Nair, U.; Kulangara Valappil, M. Optimization of parameters for the production of biodiesel from rubber seed oil using onsite lipase by response surface methodology. 3 Biotech 2018, 8, 459. [Google Scholar] [CrossRef] [PubMed]
- Chug, R.; Mathur, S.; Kothari, S.L.; Harish; Gour, V.S. Maximizing EPS production from Pseudomonas aeruginosa and its application in Cr and Ni sequestration. Biochem. Biophys. Rep. 2021, 26, 100972. [Google Scholar] [CrossRef] [PubMed]
- Peruzzo, R.; Corrà, S.; Costa, R.; Brischigliaro, M.; Varanita, T.; Biasutto, L.; Rampazzo, C.; Ghezzi, D.; Leanza, L.; Zoratti, M.; et al. Exploiting pyocyanin to treat mitochondrial disease due to respiratory complex III dysfunction. Nat. Commun. 2021, 12, 2103. [Google Scholar] [CrossRef] [PubMed]
- Moayedi, A.; Nowroozi, J.; Akhavan Sepahy, A. Cytotoxic effect of pyocyanin on human pancreatic cancer cell line (Panc-1). Iran. J. Basic Med. Sci. 2018, 21, 794–799. [Google Scholar] [CrossRef]
- Zhao, J.; Wu, Y.; Alfred, A.T.; Wei, P.; Yang, S. Anticancer effects of pyocyanin on HepG2 human hepatoma cells. Lett. Appl. Microbiol. 2014, 58, 541–548. [Google Scholar] [CrossRef]
- Kohatsu, H.; Kamo, S.; Furuta, M.; Tomoshige, S.; Kuramochi, K. Synthesis and Cytotoxic Evaluation of N-Alkyl-2-halophenazin-1-ones. ACS Omega 2020, 5, 27667–27674. [Google Scholar] [CrossRef]
- Shen, H.-B.; Yong, X.-Y.; Chen, Y.-L.; Liao, Z.-H.; Si, R.-W.; Zhou, J.; Wang, S.-Y.; Yong, Y.-C.; OuYang, P.-K.; Zheng, T. Enhanced bioelectricity generation by improving pyocyanin production and membrane permeability through sophorolipid addition in Pseudomonas aeruginosa-inoculated microbial fuel cells. Bioresour. Technol. 2014, 167, 490–494. [Google Scholar] [CrossRef]
- Kumar, R.; Singh, L.; Zularisam, A.W. Exoelectrogens: Recent advances in molecular drivers involved in extracellular electron transfer and strategies used to improve it for microbial fuel cell applications. Renew. Sustain. Energy Rev. 2016, 56, 1322–1336. [Google Scholar] [CrossRef] [Green Version]
- Islam, M.A.; Ethiraj, B.; Cheng, C.K.; Yousuf, A.; Khan, M.M.R. An Insight of Synergy between Pseudomonas aeruginosa and Klebsiella variicola in a Microbial Fuel Cell. ACS Sustain. Chem. Eng. 2018, 6, 4130–4137. [Google Scholar] [CrossRef]
- Chukwubuikem, A.; Berger, C.; Mady, A.; Rosenbaum, M.A. Role of phenazine-enzyme physiology for current generation in a bioelectrochemical system. Microb. Biotechnol. 2021, 14, 1613–1626. [Google Scholar] [CrossRef]
- Jancheva, M.; Böttcher, T. A Metabolite of Pseudomonas Triggers Prophage-Selective Lysogenic to Lytic Conversion in Staphylococcus aureus. J. Am. Chem. Soc. 2021, 143, 8344–8351. [Google Scholar] [CrossRef] [PubMed]
- Saleem, H.; Mazhar, S.; Syed, Q.; Javed, M.Q.; Adnan, A. Bio-characterization of food grade pyocyanin bio-pigment extracted from chromogenic Pseudomonas species found in Pakistani native flora. Arab. J. Chem. 2021, 14, 103005. [Google Scholar] [CrossRef]
- DeBritto, S.; Gajbar, T.D.; Satapute, P.; Sundaram, L.; Lakshmikantha, R.Y.; Jogaiah, S.; Ito, S. Isolation and characterization of nutrient dependent pyocyanin from Pseudomonas aeruginosa and its dye and agrochemical properties. Sci. Rep. 2020, 10, 1542. [Google Scholar] [CrossRef] [Green Version]
- Chong, H.; Li, Q. Microbial production of rhamnolipids: Opportunities, challenges and strategies. Microb. Cell Fact. 2017, 16, 137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varjani, S.; Rakholiya, P.; Yong Ng, H.; Taherzadeh, M.J.; Hao Ngo, H.; Chang, J.-S.; Wong, J.W.C.; You, S.; Teixeira, J.A.; Bui, X.-T. Bio-based rhamnolipids production and recovery from waste streams: Status and perspectives. Bioresour. Technol. 2021, 319, 124213. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Mawgoud, A.M.; Lépine, F.; Déziel, E. Rhamnolipids: Diversity of structures, microbial origins and roles. Appl. Microbiol. Biotechnol. 2010, 86, 1323–1336. [Google Scholar] [CrossRef] [Green Version]
- George, E.; Kumar, S.N.; Jacob, J.; Bommasani, B.; Lankalapalli, R.S.; Morang, P.; Kumar, B.S.D. Characterization of the Bioactive Metabolites from a Plant Growth-Promoting Rhizobacteria and Their Exploitation as Antimicrobial and Plant Growth-Promoting Agents. Appl. Biochem. Biotechnol. 2015, 176, 529–546. [Google Scholar] [CrossRef]
- Goswami, D.; Vaghela, H.; Parmar, S.; Dhandhukia, P.; Thakker, J.N. Plant growth promoting potentials of Pseudomonas spp. strain OG isolated from marine water. J. Plant Interact. 2013, 8, 281–290. [Google Scholar] [CrossRef] [Green Version]
- Mishra, S.; Lin, Z.; Pang, S.; Zhang, Y.; Bhatt, P.; Chen, S. Biosurfactant is a powerful tool for the bioremediation of heavy metals from contaminated soils. J. Hazard. Mater. 2021, 418, 126253. [Google Scholar] [CrossRef]
- Lan, G.; Fan, Q.; Liu, Y.; Liu, Y.; Liu, Y.; Yin, X.; Luo, M. Effects of the addition of waste cooking oil on heavy crude oil biodegradation and microbial enhanced oil recovery using Pseudomonas sp. SWP-4. Biochem. Eng. J. 2015, 103, 219–226. [Google Scholar] [CrossRef]
- Gaur, V.K.; Tripathi, V.; Gupta, P.; Dhiman, N.; Regar, R.K.; Gautam, K.; Srivastava, J.K.; Patnaik, S.; Patel, D.K.; Manickam, N. Rhamnolipids from Planococcus spp. and their mechanism of action against pathogenic bacteria. Bioresour. Technol. 2020, 307, 123206. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Dai, L.; Wang, D.; Mao, L.; Gao, Y. Stabilization and Rheology of Concentrated Emulsions Using the Natural Emulsifiers Quillaja Saponins and Rhamnolipids. J. Agric. Food Chem. 2018, 66, 3922–3929. [Google Scholar] [CrossRef] [PubMed]
- Singh, V.; Haque, S.; Khare, S.; Tiwari, A.K.; Katiyar, D.; Banerjee, B.; Kumari, K.; Tripathi, C.K.M. Isolation and purification of antibacterial compound from Streptomyces levis collected from soil sample of north India. PLoS ONE 2018, 13, e0200500. [Google Scholar] [CrossRef] [PubMed]
- Rahimi, K.; Lotfabad, T.B.; Jabeen, F.; Mohammad Ganji, S. Cytotoxic effects of mono- and di-rhamnolipids from Pseudomonas aeruginosa MR01 on MCF-7 human breast cancer cells. Colloids Surfaces B Biointerfaces 2019, 181, 943–952. [Google Scholar] [CrossRef] [PubMed]
- Bezerra, K.G.O.; Rufino, R.D.; Luna, J.M.; Sarubbo, L.A. Saponins and microbial biosurfactants: Potential raw materials for the formulation of cosmetics. Biotechnol. Prog. 2018, 34, 1482–1493. [Google Scholar] [CrossRef]
- Diab, A.M.; Ibrahim, S.; Abdulla, H.M. Safe Application and Preservation Efficacy of Low-toxic Rhamnolipids Produced from Ps. aeruginosa for Cosmetics and Personal Care Formulation. Egypt. J. Microbiol. 2020, 55, 57–70. [Google Scholar] [CrossRef]
- Konopacki, M.; Grygorcewicz, B.; Kordas, M.; Dołęgowska, B.; Rakoczy, R. Methods of Bacteriophages Production with Application of Alternate Magnetic Field. In Practical Aspects of Chemical Engineering; Ochowiak, M., Woziwodzki, S., Mitkowski, P.T., Doligalski, M., Eds.; Springer International Publishing: Cham, Germany, 2020; pp. 171–182. ISBN 978-3-030-39867-5. [Google Scholar]
- Hutchins, C.F.; Moore, G.; Thompson, K.-A.; Webb, J.; Walker, J.T. Contamination of hospital tap water: The survival and persistence of Pseudomonas aeruginosa on conventional and ‘antimicrobial’ outlet fittings. J. Hosp. Infect. 2017, 97, 156–161. [Google Scholar] [CrossRef] [Green Version]
- Palleroni, N.J. Pseudomonas. In Bergey’s Manual of Systematics of Archaea and Bacteria; Major Reference Works; Wiley: Weinheim, Germany, 2015; p. 1. ISBN 9781118960608. [Google Scholar]
- Essar, D.W.; Eberly, L.; Hadero, A.; Crawford, I.P. Identification and characterization of genes for a second anthranilate synthase in Pseudomonas aeruginosa: Interchangeability of the two anthranilate synthase and evolutionary implications. J. Bacteriol. 1990, 172, 884–900. [Google Scholar] [CrossRef] [Green Version]
- Pinzon, N.M.; Ju, L.K. Analysis of rhamnolipid biosurfactants by methylene blue complexation. Appl. Microbiol. Biotechnol. 2009, 82, 975–981. [Google Scholar] [CrossRef]
- Zlokarnik, M. Homogenization. In Stirring; Wiley-VCH Verlag GmbH: Weinheim, Germany, 2001; pp. 97–125. ISBN 9783527612703. [Google Scholar]
- Jaszczur, M.; Młynarczykowska, A. A general review of the current development of mechanically agitated vessels. Processes 2020, 8, 982. [Google Scholar] [CrossRef]
- Cudak, M. Hydrodynamic characteristics of mechanically agitated air—Aqueous sucrose solutions. Chem. Process. Eng. 2014, 35, 97–107. [Google Scholar] [CrossRef] [Green Version]
- Lange, H.; Taillandier, P.; Riba, J.-P. Effect of high shear stress on microbial viability. J. Chem. Technol. Biotechnol. 2001, 76, 501–505. [Google Scholar] [CrossRef]
- Konopacki, M.; Augustyniak, A.; Grygorcewicz, B.; Dołęgowska, B.; Kordas, M.; Rakoczy, R. Single Mathematical Parameter for Evaluation of the Microorganisms’ Growth as the Objective Function in the Optimization by the DOE Techniques. Microorganisms 2020, 8, 1706. [Google Scholar] [CrossRef]
- Arrua, L.A.; McCoy, B.J.; Smith, J.M. Gas–liquid mass transfer in stirred tanks. AIChE J. 1990, 36, 1768–1772. [Google Scholar] [CrossRef]
- Karimi, A.; Golbabaei, F.; Mehrnia, M.R.; Neghab, M.; Mohammad, K.; Nikpey, A.; Pourmand, M.R. Oxygen mass transfer in a stirred tank bioreactor using different impeller configurations for environmental purposes. Iran. J. Environ. Health Sci. Eng. 2013, 10, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Augustyniak, A.; Dubrowska, K.; Jabłońska, J.; Cendrowski, K.; Wróbel, R.J.; Piz, M.; Filipek, E.; Rakoczy, R. Basic physiology of Pseudomonas aeruginosa contacted with carbon nanocomposites. Appl. Nanosci. 2022, 12, 1917–1927. [Google Scholar] [CrossRef]
- Jabłońska, J.; Dubrowska, K.; Augustyniak, A.; Wróbel, R.J.; Piz, M.; Cendrowski, K.; Rakoczy, R. The influence of nanomaterials on pyocyanin production by Pseudomonas aeruginosa. Appl. Nanosci. 2022, 12, 1929–1940. [Google Scholar] [CrossRef]
- Jabłońska, J.; Dubrowska, K.; Augustyniak, A.; Kordas, M.; Rakoczy, R. Application of Magnetically Assisted Reactors for Modulation of Growth and Pyocyanin Production by Pseudomonas aeruginosa. Front. Bioeng. Biotechnol. 2022, 10, 795871. [Google Scholar] [CrossRef]
- Ozdal, M.; Gurkok, S.; Ozdal, O.G.; Kurbanoglu, E.B. Enhancement of pyocyanin production by Pseudomonas aeruginosa via the addition of n-hexane as an oxygen vector. Biocatal. Agric. Biotechnol. 2019, 22, 101365. [Google Scholar] [CrossRef]
- Ozdal, M. A new strategy for the efficient production of pyocyanin, a versatile pigment, in Pseudomonas aeruginosa OG1 via toluene addition. 3 Biotech 2019, 9, 374. [Google Scholar] [CrossRef]
- Chincholkar, S.; Thomashow, L. Microbial Phenazines; Chincholkar, S., Thomashow, L., Eds.; Springer: Berlin/Heidelberg, Germany, 2013; ISBN 978-3-642-40572-3. [Google Scholar]
- El-Fouly, M.Z.; Sharaf, A.M.; Shahin, A.A.M.; El-Bialy, H.A.; Omara, A.M.A. Biosynthesis of pyocyanin pigment by Pseudomonas aeruginosa. J. Radiat. Res. Appl. Sci. 2015, 8, 36–48. [Google Scholar] [CrossRef] [Green Version]
- Elbargisy, R.M. Optimization of nutritional and environmental conditions for pyocyanin production by urine isolates of Pseudomonas aeruginosa. Saudi J. Biol. Sci. 2021, 28, 993–1000. [Google Scholar] [CrossRef] [PubMed]
- Das, T.; Kutty, S.K.; Kumar, N.; Manefield, M. Pyocyanin Facilitates Extracellular DNA Binding to Pseudomonas aeruginosa Influencing Cell Surface Properties and Aggregation. PLoS ONE 2013, 8, e58299. [Google Scholar] [CrossRef] [PubMed]
- Zhao, F.; Shi, R.; Ma, F.; Han, S.; Zhang, Y. Oxygen effects on rhamnolipids production by Pseudomonas aeruginosa. Microb. Cell Fact. 2018, 17, 39. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Konopacki, M.; Jabłońska, J.; Dubrowska, K.; Augustyniak, A.; Grygorcewicz, B.; Gliźniewicz, M.; Wróblewski, E.; Kordas, M.; Dołęgowska, B.; Rakoczy, R. The Influence of Hydrodynamic Conditions in a Laboratory-Scale Bioreactor on Pseudomonas aeruginosa Metabolite Production. Microorganisms 2023, 11, 88. https://doi.org/10.3390/microorganisms11010088
Konopacki M, Jabłońska J, Dubrowska K, Augustyniak A, Grygorcewicz B, Gliźniewicz M, Wróblewski E, Kordas M, Dołęgowska B, Rakoczy R. The Influence of Hydrodynamic Conditions in a Laboratory-Scale Bioreactor on Pseudomonas aeruginosa Metabolite Production. Microorganisms. 2023; 11(1):88. https://doi.org/10.3390/microorganisms11010088
Chicago/Turabian StyleKonopacki, Maciej, Joanna Jabłońska, Kamila Dubrowska, Adrian Augustyniak, Bartłomiej Grygorcewicz, Marta Gliźniewicz, Emil Wróblewski, Marian Kordas, Barbara Dołęgowska, and Rafał Rakoczy. 2023. "The Influence of Hydrodynamic Conditions in a Laboratory-Scale Bioreactor on Pseudomonas aeruginosa Metabolite Production" Microorganisms 11, no. 1: 88. https://doi.org/10.3390/microorganisms11010088