

Mechanisms of BPA Degradation and Toxicity Resistance in Rhodococcus equi
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains and Chemicals
2.2. R-001 Degrades BPA
2.3. Detection Methods
2.4. Transcriptome Sequencing
2.5. qPCR Analysis of the BPA-Degrading Genes
2.6. Bioinformatics Analysis
3. Results and Discussion
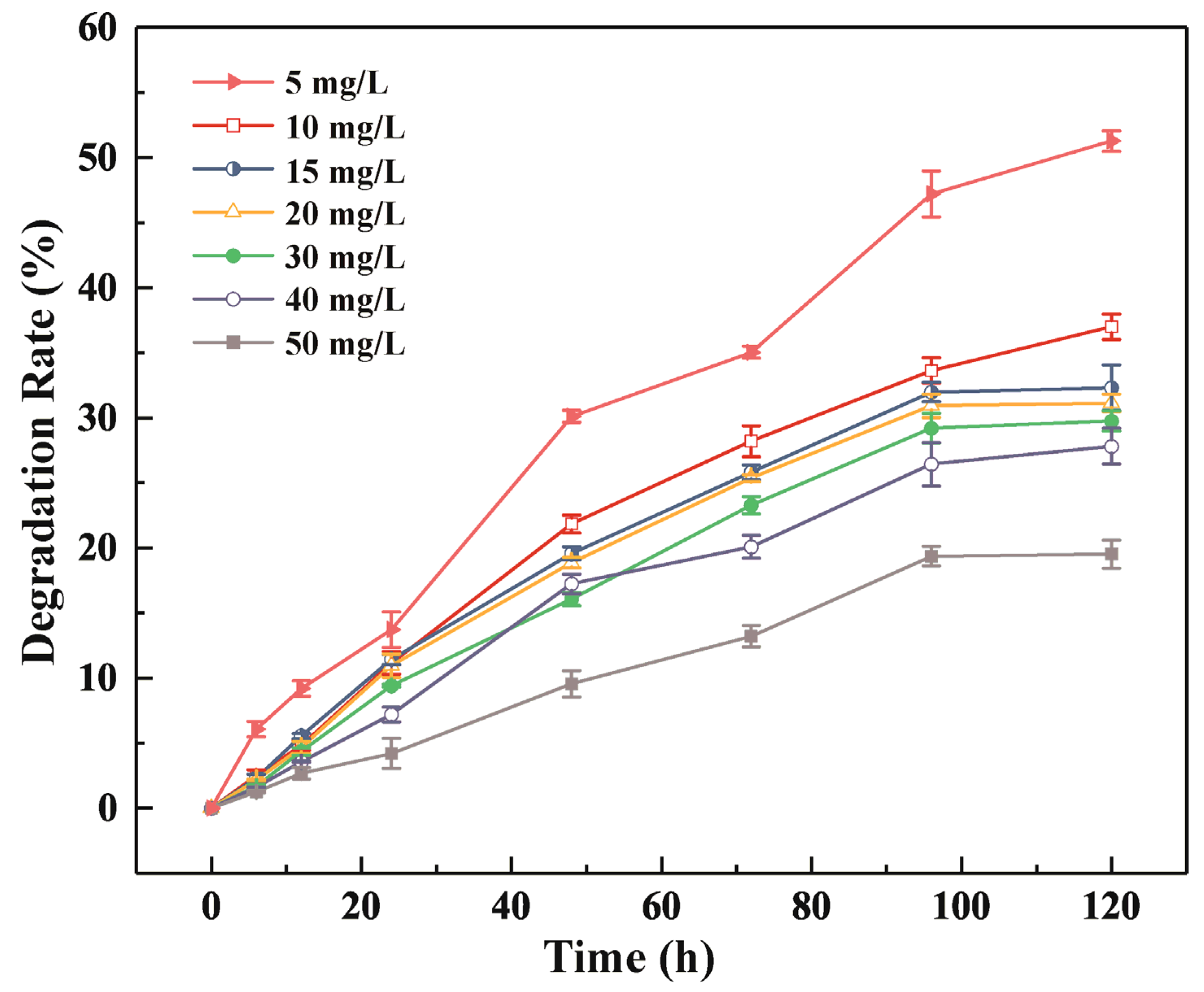
3.1. BPA Degradation by R-001
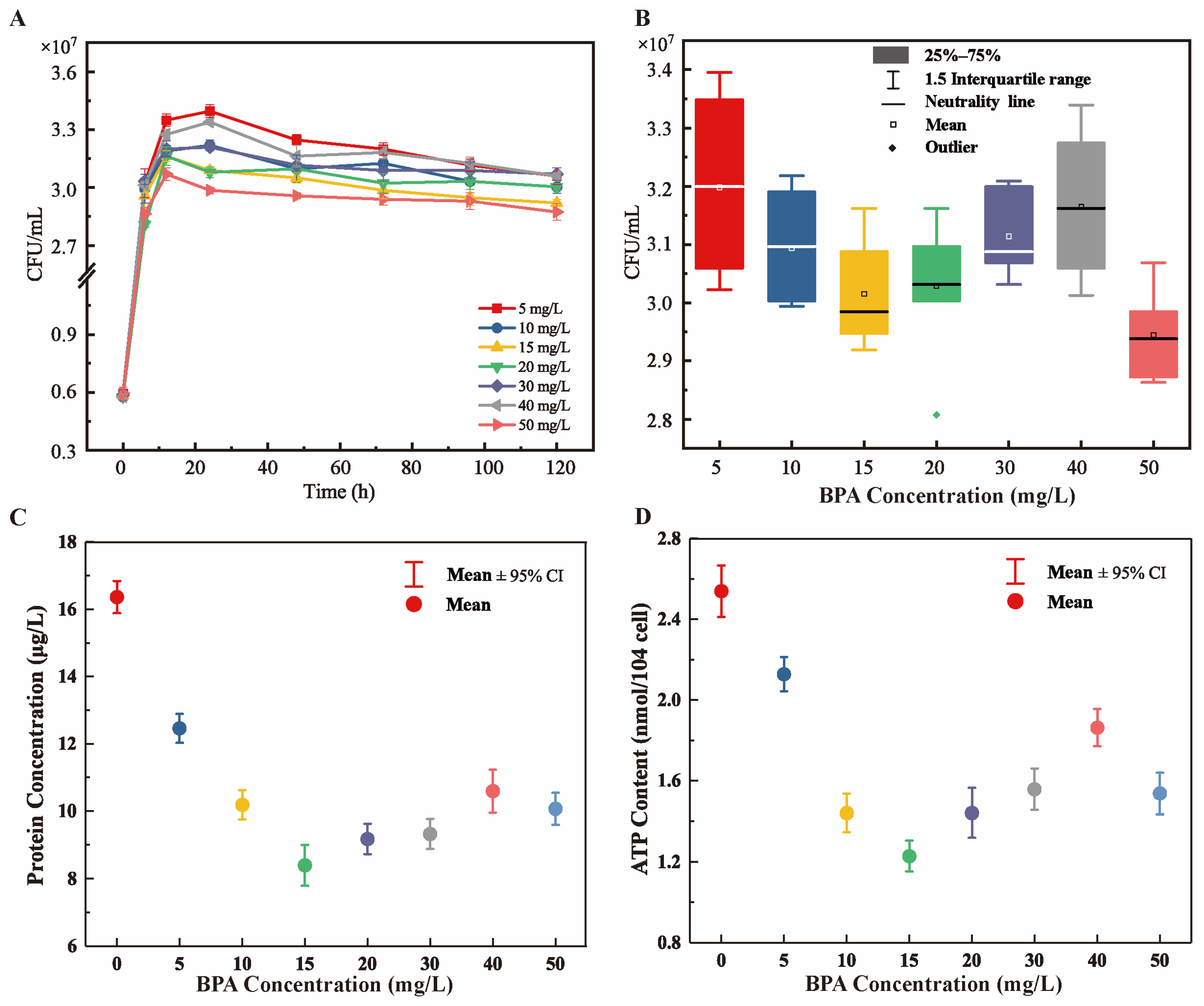
3.2. Toxic Effects of BPA on the Growth and Metabolism of R-001
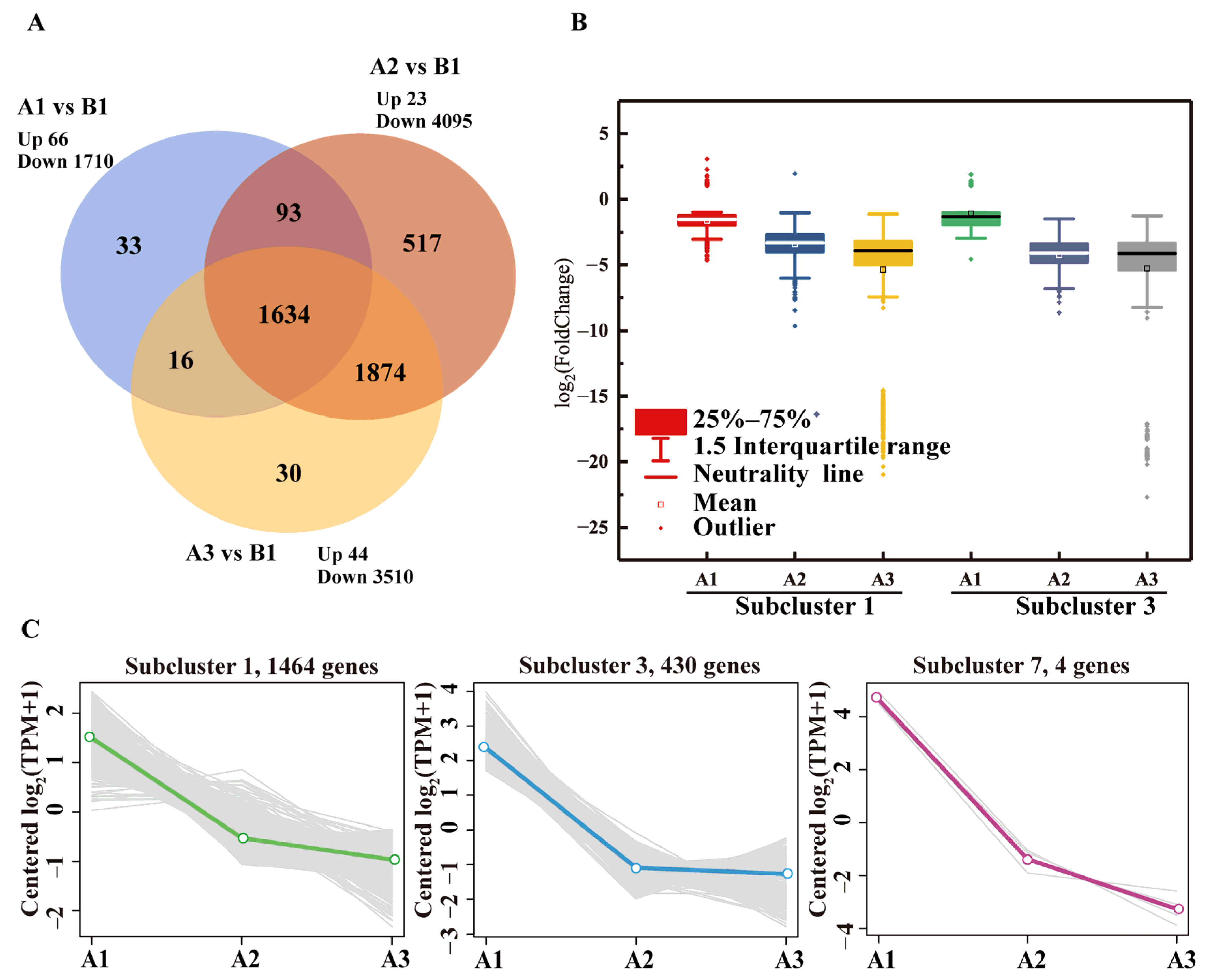
3.3. Inhibitory Effects of BPA on the Expression Profiles of Metabolic Genes
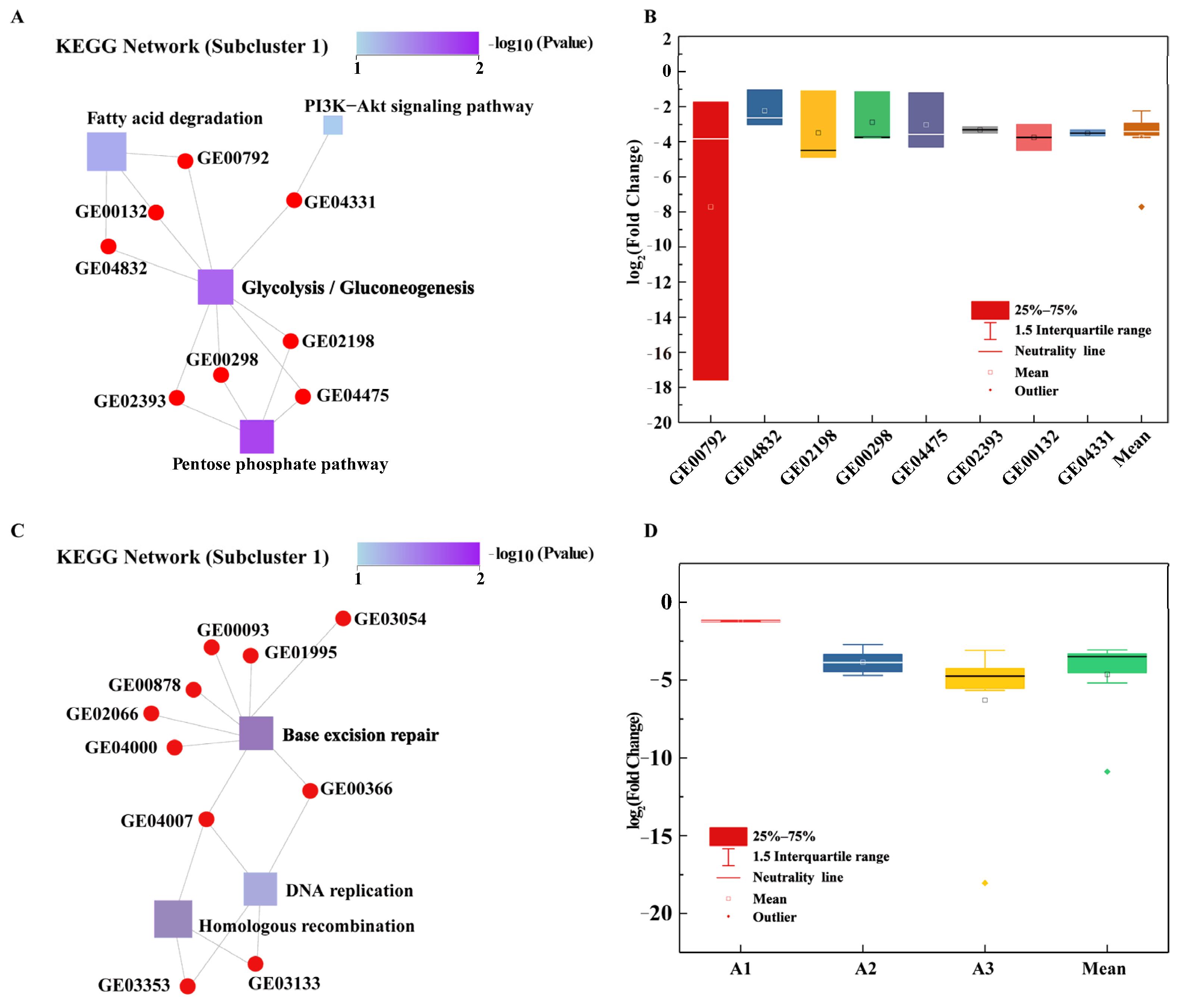
3.3.1. Glycolysis/Gluconeogenesis Pathway
3.3.2. Glyoxylate and Dicarboxylate Metabolism
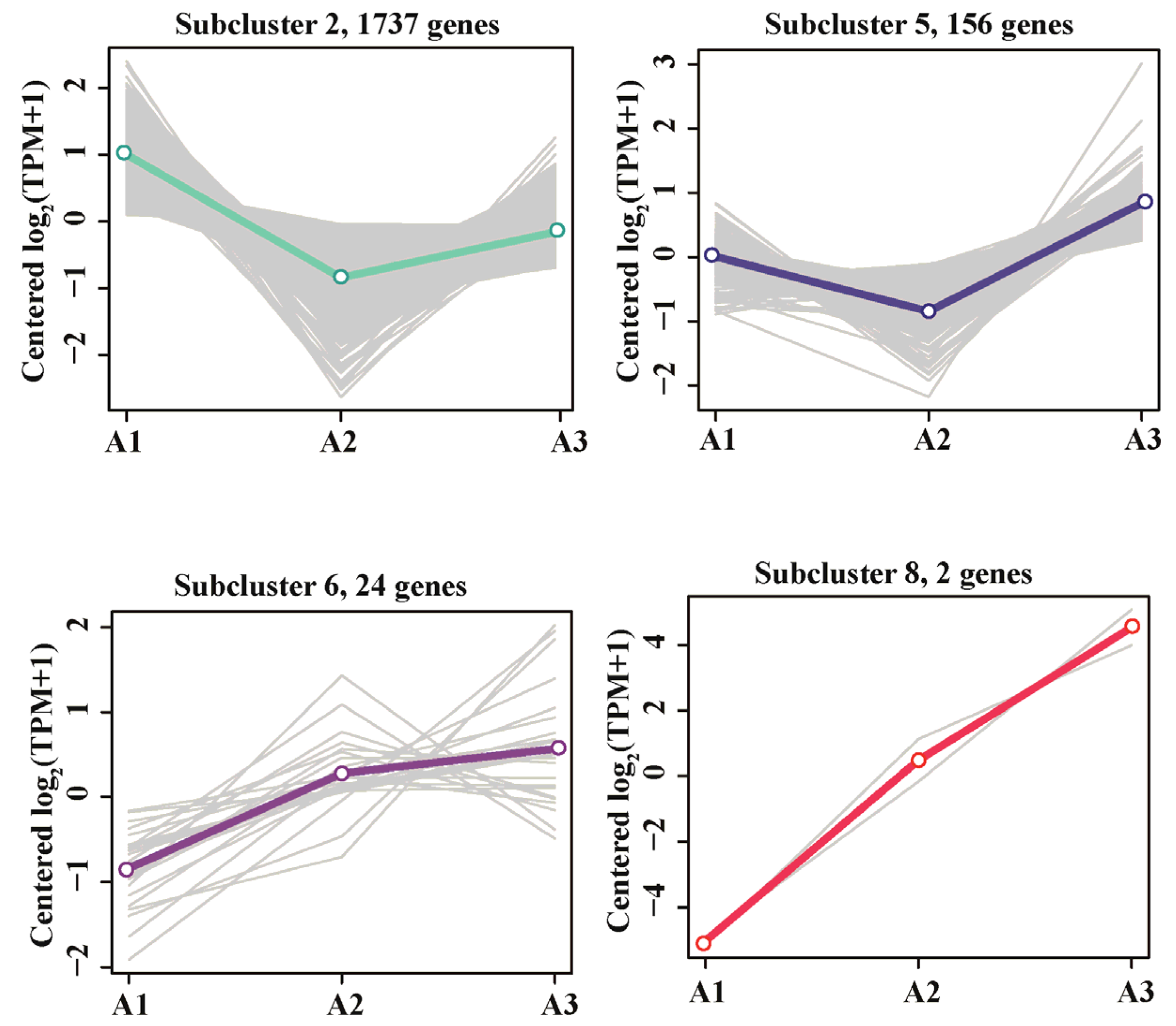
3.4. Resistance of R-001 to BPA Toxicity
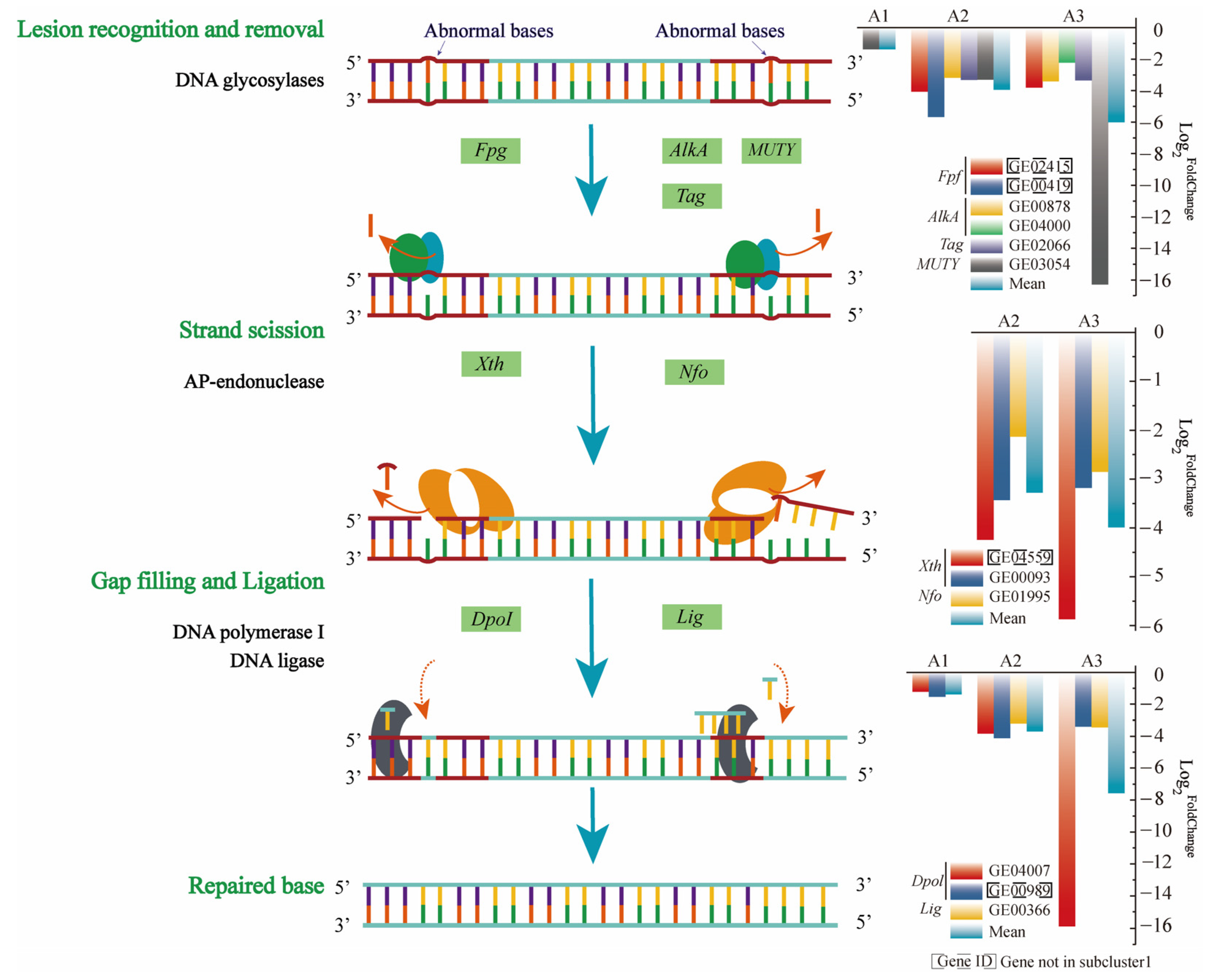
3.4.1. Downregulation of BER-Related Genes to Increase Strain Mutation
3.4.2. Upregulation of Metabolic Genes to Maintain Energy Supply
3.4.3. Upregulation of Transport-System Genes to Maintain Cell Homeostasis
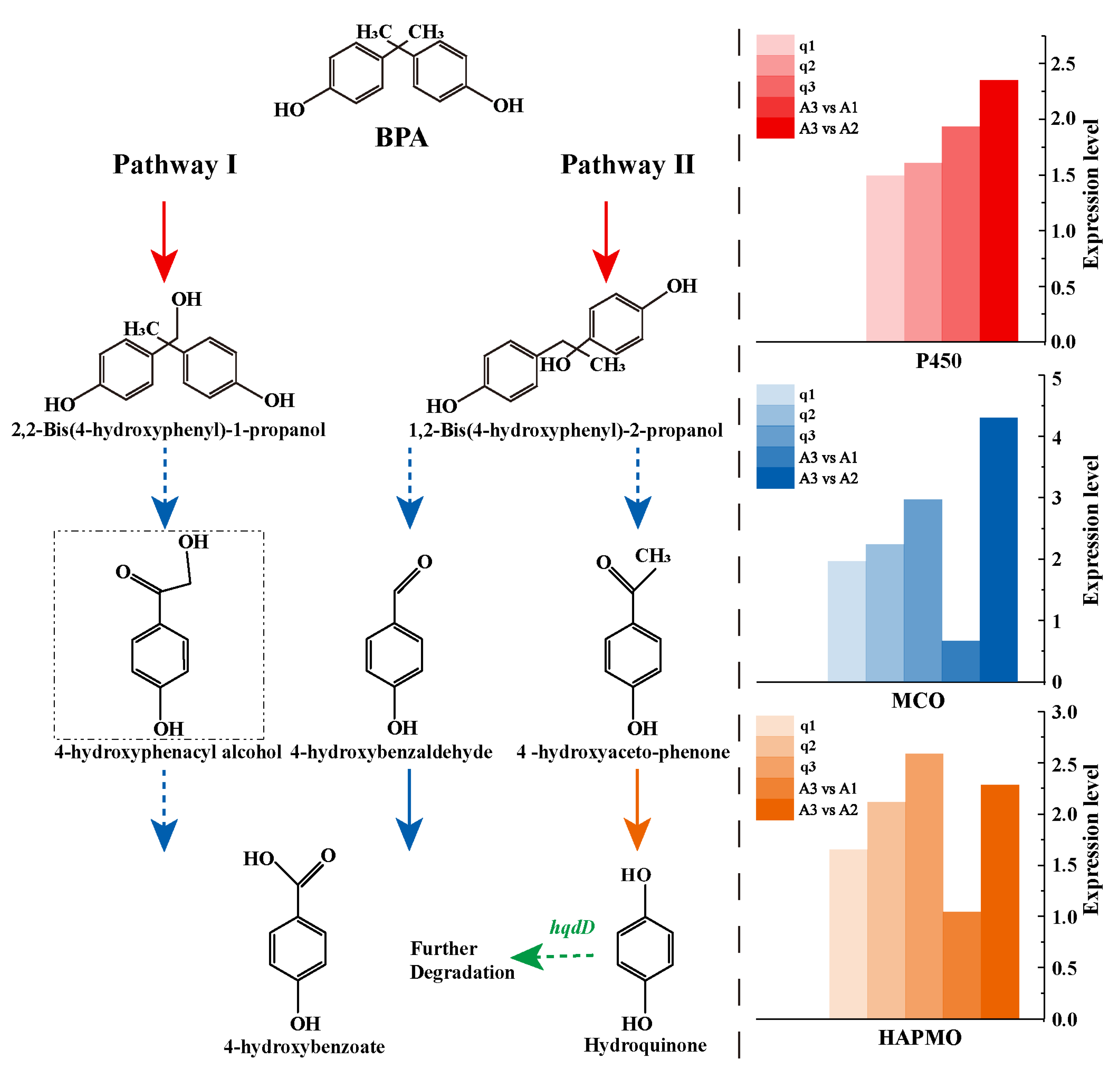
3.5. Genes and Pathways Associated with BPA Degradation in Strain R-001
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Viñas, R.; Jeng, Y.-J.; Watson, C.S. Non-Genomic Effects of Xenoestrogen Mixtures. Int. J. Environ. Res. Public Health 2012, 9, 2694–2714. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Xu, L.-C.; Chen, J.-F.; Song, L.; Wang, X.R. Effect of bisphenol A, tetrachlorobisphenol A and pentachlorophenol on the transcriptional activities of androgen receptor-mediated reporter gene. Food Chem. Toxicol. 2006, 44, 1916–1921. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Wang, Y.; Chen, Z.; Ren, J.; Rehman, A.; Ahmad, D.W.; Long, D.; Hou, J.; Zhou, Y.; Yang, L.; et al. Single-cell transcriptome analysis of Bisphenol A exposure reveals the key roles of the testicular microenvironment in male reproduction. Biomed. Pharmacother. 2022, 145, 112449. [Google Scholar] [CrossRef] [PubMed]
- Mustieles, V.; D’Cruz, S.C.; Couderq, S.; Rodríguez-Carrillo, A.; Fini, J.B.; Hofer, T.; Steffensen, I.L.; Dirven, H.; Barouki, R.; Olea, N.; et al. Bisphenol A and its analogues: A comprehensive review to identify and prioritize effect biomarkers for human biomonitoring. Environ. Int. 2020, 144, 105811. [Google Scholar] [CrossRef]
- Seok, J.W.; Park, J.Y.; Park, H.K.; Lee, H. Endrin potentiates early-stage adipogenesis in 3T3-L1 cells by activating the mammalian target of rapamycin. Life Sci. 2022, 288, 120151. [Google Scholar] [CrossRef]
- Tarapore, P.; Ying, J.; Ouyang, B.; Burke, B.; Bracken, B.; Ho, S.M. Exposure to Bisphenol a Correlates with Early-Onset Prostate Cancer and Promotes Centrosome Amplification and Anchorage Independent Growth In Vitro. Endocr. Rev. 2014, 9, e90332. [Google Scholar] [CrossRef] [Green Version]
- Acevedo, N.; Davis, B.; Schaeberle, C.M.; Sonnenschein, C.; Soto, A.M. Perinatally Administered Bisphenol A as a Potential Mammary Gland Carcinogen in Rats. Environ. Health Perspect. 2013, 121, 1040–1046. [Google Scholar] [CrossRef] [Green Version]
- Park, M.; Choi, K.J. Effects of 4-nonylphenol and bisphenol A on stimulation of cell growth via disruption of the transforming growth factor-β signaling pathway in ovarian cancer models. Chem. Res. Toxicol. 2014, 27, 119–128. [Google Scholar] [CrossRef]
- Liu, S.; Pan, C.; Tang, Y.; Chen, F.; Yang, M.; Wang, K.J. Identification of novel long non-coding RNAs involved in bisphenol A induced immunotoxicity in fish primary macrophages. Fish Shellfish Immunol. 2020, 100, 152–160. [Google Scholar] [CrossRef]
- Burgos-Aceves, M.A.; Abo-Al-Ela, H.G.; Faggio, C. Impact of phthalates and bisphenols plasticizers on haemocyte immune function of aquatic invertebrates: A review on physiological, biochemical, and genomic aspects. J. Hazard. Mater. 2021, 419, 126426. [Google Scholar] [CrossRef]
- McDonough, C.M.; Xu, H.S.; Guo, T.L. Toxicity of bisphenol analogues on the reproductive, nervous, and immune systems, and their relationships to gut microbiome and metabolism: Insights from a multi-species comparison. Crit. Rev. Toxicol. 2021, 51, 283–300. [Google Scholar] [CrossRef] [PubMed]
- Alexander, H.C.; Dill, D.C.; Smith, L.W.; Guiney, P.D.; Dorn, P. Bisphenol A: Acute aquatic toxicity. Environ. Toxicol. Chem. 1988, 7, 19–26. [Google Scholar] [CrossRef]
- Zhang, W.; Xiong, B.; Sun, W.-F.; An, S.; Lin, K.F.; Guo, M.J.; Cui, X.H. Acute and chronic toxic effects of bisphenol a on Chlorella pyrenoidosa and Scenedesmus obliquus. Environ. Toxicol. 2014, 29, 714–722. [Google Scholar] [CrossRef] [PubMed]
- Mathieu-Denoncourt, J.; Wallace, S.J.; de Solla, S.R.; Langlois, V.S. Influence of Lipophilicity on the Toxicity of Bisphenol A and Phthalates to Aquatic Organisms. Bull. Environ. Contam. Toxicol. 2016, 97, 4–10. [Google Scholar] [CrossRef] [PubMed]
- Huang, R.P.; Liu, Z.H.; Yin, H.; Dang, Z.; Wu, P.X.; Zhu, N.W.; Lin, Z. Bisphenol A concentrations in human urine, human intakes across six continents, and annual trends of average intakes in adult and child populations worldwide: A thorough literature review. Sci. Total Environ. 2018, 626, 971–981. [Google Scholar] [CrossRef]
- Zhang, W.; Yin, K.; Chen, L. Bacteria-mediated bisphenol A degradation. Appl. Microbiol. Biotechnol. 2013, 97, 5681–5689. [Google Scholar] [CrossRef]
- Zuehlke, M.-K.; Schlueter, R.; Mikolasch, A.; Zühlke, D.; Giersberg, M.; Schindler, H.; Henning, A.K.; Frenzel, H.; Hammer, E.; Lalk, M.; et al. Biotransformation and reduction of estrogenicity of bisphenol A by the biphenyl-degrading Cupriavidus basilensis. Appl. Microbiol. Biotechnol. 2017, 101, 3743–3758. [Google Scholar] [CrossRef]
- López-Moreno, A.; Torres-Sánchez, A.; Acuña, I.; Suárez, A.; Aguilera, M. Representative Bacillus sp. AM1 from Gut Microbiota Harbor Versatile Molecular Pathways for Bisphenol A Biodegradation. Int. J. Mol. Sci. 2021, 22, 4952. [Google Scholar] [CrossRef]
- Suyamud, B.; Inthorn, D.; Panyapinyopol, B.; Thiravetyan, P. Biodegradation of Bisphenol A by a Newly Isolated Bacillus megaterium Strain ISO-2 from a Polycarbonate Industrial Wastewater. Water Air Soil Pollut. 2018, 229, 348. [Google Scholar] [CrossRef]
- Matsumura, Y.; Akahira-Moriya, A.; Sasaki-Mori, M. Bioremediation of Bisphenol-A Polluted Soil by Sphingomonas bisphenolicum AO1 and the Microbial Community Existing in the Soil. Biocontrol Sci. 2015, 20, 35–42. [Google Scholar] [CrossRef]
- Chen, X.; Zhao, J.; Bao, L.; Wang, L.; Zhang, Y. The investigation of different pollutants and operation processes on sludge toxicity in sequencing batch bioreactors. Environ. Technol. 2016, 37, 2048–2057. [Google Scholar] [CrossRef] [PubMed]
- Shao, J.; Tian, K.; Meng, F.; Li, S.; Li, H.; Yu, Y.; Qiu, Q.; Chang, M.; Huo, H. Effects of Bisphenol A Stress on Activated Sludge in Sequential Batch Reactors and Functional Recovery. Appl. Sci. 2022, 12, 8026. [Google Scholar] [CrossRef]
- Zaborowska, M.; Wyszkowska, J.; Borowik, A.; Kucharski, J. Bisphenol A—A Dangerous Pollutant Distorting the Biological Properties of Soil. Int. J. Mol. Sci. 2021, 22, 12753. [Google Scholar] [CrossRef] [PubMed]
- Cydzik-Kwiatkowska, A.; Grzyb, M.; Jachimowicz, P. Metatranscriptome Analysis of Bisphenol A-Exposed Aerobic Granular Sludge. Energies 2021, 14, 3263. [Google Scholar] [CrossRef]
- Tian, K.; Meng, F.; Meng, Q.; Gao, Y.; Zhang, L.; Wang, L.; Wang, Y.; Li, X.; Huo, H. The Analysis of Estrogen-Degrading and Functional Metabolism Genes in Rhodococcus equi DSSKP-R-001. Int. J. Genom. 2020, 2020, 9369182. [Google Scholar] [CrossRef]
- Yamanaka, H.; Moriyoshi, K.; Ohmoto, T.; Ohe, T.; Sakai, K. Degradation of bisphenol A by Bacillus pumilus isolated from kimchi, a traditionally fermented food. Appl. Biochem. Biotechnol. 2007, 136, 39–51. [Google Scholar] [CrossRef]
- Kyrila, G.; Katsoulas, A.; Schoretsaniti, V.; Rigopoulos, A.; Rizou, E.; Doulgeridou, S.; Sarli, V.; Samanidou, V.; Touraki, M. Bisphenol A removal and degradation pathways in microorganisms with probiotic properties. J. Hazard. Mater. 2021, 413, 125363. [Google Scholar] [CrossRef]
- Fischer, J.; Kappelmeyer, U.; Kastner, M.; Schauer, F.; Heipieper, H.J. The degradation of bisphenol A by the newly isolated bacterium Cupriavidus basilensis JF1 can be enhanced by biostimulation with phenol. Int. Biodeterior. Biodegrad. 2010, 64, 324–330. [Google Scholar] [CrossRef]
- Zhang, X.-M.; Li, Y.; Gu, Y.-X.; Zhang, C.N.; Lai, H.C.; Shi, J.Y. Ta-Coated Titanium Surface with Superior Bacteriostasis And Osseointegration. Int. J. Nanomed. 2019, 14, 8693–8706. [Google Scholar] [CrossRef] [Green Version]
- Shobnam, N.; Sun, Y.; Mahmood, M.; Löffler, F.E.; Im, J. Biologically mediated abiotic degradation (BMAD) of bisphenol A by manganese-oxidizing bacteria. J. Hazard. Mater. 2021, 417, 125987. [Google Scholar] [CrossRef]
- Malea, P.; Kokkinidi, D.; Kevrekidou, A.; Adamakis, I.D.S. The Enzymatic and Non-Enzymatic Antioxidant System Response of the Seagrass Cymodocea nodosa to Bisphenol-A Toxicity. Int. J. Mol. Sci. 2022, 23, 1348. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Ou, C.; Zhu, X.; Tan, C.; Xiang, X.; He, Y. Potential role of CFTR in bisphenol A-induced malignant transformation of prostate cells via mitochondrial apoptosis. Toxicol. Ind. Health 2020, 36, 531–539. [Google Scholar] [CrossRef] [PubMed]
- Adamakis, I.-D.S.; Malea, P.; Sperdouli, I.; Panteris, E.; Kokkinidi, D.; Moustakas, M. Evaluation of the spatiotemporal effects of bisphenol A on the leaves of the seagrass Cymodocea nodosa. J. Hazard. Mater. 2021, 404, 124001. [Google Scholar] [CrossRef] [PubMed]
- Fukuda, W.; Fukui, T.; Atomi, H.; Imanaka, T. First Characterization of an Archaeal GTP-Dependent Phosphoenolpyruvate Carboxykinase from the Hyperthermophilic Archaeon Thermococcus kodakaraensis KOD1. J. Bacteriol. 2004, 186, 4620–4627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, V.; Siedenburg, G.; Birke, J.; Mobeen, F.; Jendrossek, D.; Prakash, T. Metabolic and taxonomic insights into the Gram-negative natural rubber degrading bacterium Steroidobacter cummioxidans sp. nov., strain 35Y. PLoS ONE 2018, 13, e0197448. [Google Scholar]
- Quinonez, C.G.; Lee, J.J.; Lim, J.; Odell, M.; Lawson, C.P.; Anyogu, A.; Raheem, S.; Eoh, H. Fatty acid metabolism of Mycobacterium tuberculosis: A double-edged sword. Microb Cell 2022, 9, 123–125. [Google Scholar] [CrossRef]
- Hansen, T.; Wendorff, D.; Schönheit, P. Bifunctional Phosphoglucose/Phosphomannose Isomerases from the Archaea Aeropyrum pernix and Thermoplasma acidophilum Constitute a Novel Enzyme Family within the Phosphoglucose Isomerase Superfamily. J. Biol. Chem. 2004, 279, 2262–2272. [Google Scholar] [CrossRef] [Green Version]
- Wei, G.; Zou, L.F.; Li, Y.R.; Cui, Y.P.; Ji, Z.Y.; Cai, L.L.; Zou, H.S.; Hutchins, W.C.; Yang, C.H.; Chen, G.Y. Fructose-Bisphophate Aldolase Exhibits Functional Roles between Carbon Metabolism and the hrp System in Rice Pathogen Xanthomonas oryzae pv. oryzicola. PLoS ONE 2012, 7, e31855. [Google Scholar]
- Guo, W.; Zou, L.; Ji, Z.; Cai, L.; Chen, G. Glucose 6-phosphate isomerase (Pgi) is required for extracellular polysaccharide biosynthesis, DSF signals production and full virulence of Xanthomonas oryzae pv. oryzicola in rice. Physiol. Mol. Plant Pathol. 2017, 100, 209–219. [Google Scholar] [CrossRef]
- Degeest, B.; De Vuyst, L. Correlation of Activities of the Enzymes α-Phosphoglucomutase, UDP-Galactose 4-Epimerase, and UDP-Glucose Pyrophosphorylase with Exopolysaccharide Biosynthesis by Streptococcus thermophilus LY03. Appl. Environ. Microbiol. 2000, 66, 3519–3527. [Google Scholar] [CrossRef] [Green Version]
- Gail, G.H.; Melissa, J.C.; Yother, J. Capsule Biosynthesis and Basic Metabolism in Streptococcus pneumoniae Are Linked through the Cellular Phosphoglucomutase. J. Bacteriol. 2000, 182, 1854–1863. [Google Scholar]
- Rittmann, D.; Schaffer, S.; Wendisch, V.F.; Sahm, H. Fructose-1,6-bisphosphatase from Corynebacterium glutamicum: Expression and deletion of the fbp gene and biochemical characterization of the enzyme. Arch. Microbiol. 2003, 180, 285–292. [Google Scholar] [CrossRef] [PubMed]
- Zgiby, S.M.; Thomson, G.J.; Qamar, S.; Berry, A. Exploring substrate binding and discrimination in fructose1,6-bisphosphate and tagatose 1,6-bisphosphate aldolases. Eur. J. Biochem. 2000, 267, 1858–1868. [Google Scholar] [CrossRef] [PubMed]
- Qi, X.; Yin, H.; Zhu, M.; Shao, P.; Dang, Z. Understanding the role of biochar in affecting BDE-47 biodegradation by Pseudomonas plecoglossicida: An integrated analysis using chemical, biological, and metabolomic approaches. Water Res. 2022, 220, 118679. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Lin, C.-Y.; Chen, S.-H.; Lo, K.J.; Liu, C.T.; Chou, T.H.; Shih, Y.H. Using high-throughput transcriptome sequencing to investigate the biotransformation mechanism of hexabromocyclododecane with Rhodopseudomonas palustris in water. Sci. Total Environ. 2019, 692, 249–258. [Google Scholar] [CrossRef]
- Wang, W.; Yu, H.; Qin, H.; Long, Y.; Ye, J.; Qu, Y. Bisphenol A degradation pathway and associated metabolic networks in Escherichia coli harboring the gene encoding CYP450. J. Hazard. Mater. 2020, 388, 121737. [Google Scholar] [CrossRef]
- Kurthkoti, K.; Varshney, U. Base excision and nucleotide excision repair pathways in mycobacteria. Tuberculosis 2011, 91, 533–543. [Google Scholar] [CrossRef]
- Fromme, J.C.; Banerjee, A.; Verdine, G.L. DNA glycosylase recognition and catalysis. Curr. Opin. Struct. Biol. 2004, 14, 43–49. [Google Scholar] [CrossRef]
- Boiteux, S.; Guillet, M. Abasic sites in DNA: Repair and biological consequences in Saccharomyces cerevisiae. DNA Repair 2004, 3, 1–12. [Google Scholar] [CrossRef]
- Schroeder, J.W.; Randall, J.R.; Hirst, W.G.; O’Donnell, M.E.; Simmons, L.A. Mutagenic cost of ribonucleotides in bacterial DNA. Proc. Natl. Acad. Sci. USA 2017, 114, 11733–11738. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Sun, Z. Successful application of municipal domestic wastewater as a co-substrate in 2,4,6-trichlorophenol degradation. Chemosphere 2021, 280, 130707. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Wang, Z.; You, Y.; Xu, W.; Lv, Z.; Liu, Z.; Chen, W.; Shi, Y. Response of Arthrobacter QD 15-4 to dimethyl phthalate by regulating energy metabolism and ABC transporters. Ecotoxicol. Environ. Saf. 2019, 174, 146–152. [Google Scholar] [CrossRef] [PubMed]
- Chittori, S.; Savithri, H.S.; Murthy, M.R.N. Preliminary X-ray crystallographic studies on acetate kinase (AckA) from Salmonella typhimurium in two crystal forms. Acta Crystallogr. Sect. F Struct. Biol. Commun. 2011, 67, 1658–1661. [Google Scholar] [CrossRef] [PubMed]
- Erb, T.J.; Retey, J.; Fuchs, G.; Alber, B.E. Ethylmalonyl-CoA Mutase from Rhodobacter sphaeroides Defines a New Subclade of Coenzyme B-12-dependent Acyl-CoA Mutases. J. Biol. Chem. 2008, 283, 32283–32293. [Google Scholar] [CrossRef] [Green Version]
- Jitrapakdee, S.; St Maurice, M.; Rayment, I.; Cleland, W.W.; Wallace, J.C.; Attwood, P.V. Structure, mechanism and regulation of pyruvate carboxylase. Biochem. J. 2008, 413, 369–387. [Google Scholar] [CrossRef] [Green Version]
- Teichmann, L.; Chen, C.; Hoffmann, T.; Smits, S.H.; Schmitt, L.; Bremer, E. From substrate specificity to promiscuity: Hybrid ABC transporters for osmoprotectants. Mol. Microbiol. 2017, 104, 761–780. [Google Scholar] [CrossRef] [Green Version]
- Du, Y.; Shi, W.-W.; He, Y.-X.; Yang, Y.H.; Zhou, C.Z.; Chen, Y. Structures of the substrate-binding protein provide insights into the multiple compatible solute binding specificities of the Bacillus subtilis ABC transporter OpuC. Biochem. J. 2011, 436, 283–289. [Google Scholar] [CrossRef] [Green Version]
- Carter, D.M.; Miousse, I.R.; Gagnon, J.-N.; Martinez, E.; Clements, A.; Lee, J.; Hancock, M.A.; Gagnon, H.; Pawelek, P.D.; Coulton, J.W. Interactions between TonB from Escherichia coli and the Periplasmic Protein FhuD. J. Biol. Chem. 2006, 281, 35413–35424. [Google Scholar] [CrossRef] [Green Version]
- Sasaki, M.; Akahira, A.; Oshiman, K.-I.; Tsuchido, T.; Matsumura, Y. Purification of Cytochrome P450 and Ferredoxin, Involved in Bisphenol A Degradation, from Sphingomonas sp. Strain AO1. Appl. Environ. Microbiol. 2005, 71, 8024–8030. [Google Scholar] [CrossRef] [Green Version]
- Sasaki, M.; Maki, J.-I.; Oshiman, K.-I.; Matsumura, Y.; Tsuchido, T. Biodegradation of bisphenol A by cells and cell lysate from Sphingomonas sp. strain AO1. Biodegradation 2005, 16, 449–459. [Google Scholar] [CrossRef]
- Das, R.; Liang, Z.; Li, G.; Mai, B.; An, T. Genome sequence of a spore-laccase forming, BPA-degrading Bacillus sp. GZB isolated from an electronic-waste recycling site reveals insights into BPA degradation pathways. Arch. Microbiol. 2019, 201, 623–638. [Google Scholar] [CrossRef] [PubMed]
- Kolvenbach, B.; Schlaich, N.; Raoui, Z.; Prell, J.; Zuhlke, S.; Schaffer, A.; Guengerich, F.P.; Corvini, P.F.X. Degradation Pathway of Bisphenol A: Does ipso Substitution Apply to Phenols Containing a Quaternary α-Carbon Structure in the para Position? Appl. Environ. Microbiol. 2007, 73, 4776–4784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Held, C.; Kandelbauer, A.; Schroeder, M.; Cavaco-Paulo, A.; Gübitz, G.M. Biotransformation of phenolics with laccase containing bacterial spores. Environ. Chem. Lett. 2005, 3, 74–77. [Google Scholar] [CrossRef]
- Das, R.; Li, G.; Mai, B.; An, T. Spore cells from BPA degrading bacteria Bacillus sp. GZB displaying high laccase activity and stability for BPA degradation. Sci. Total Environ. 2018, 640–641, 798–806. [Google Scholar] [CrossRef]
- Kolvenbach, B.A.; Dobrowinski, H.; Fousek, J.; Vlcek, C.; Schäffer, A.; Gabriel, F.L.; Kohler, H.P.E.; Corvini, P.F. An unexpected gene cluster for downstream degradation of alkylphenols in Sphingomonas sp. strain TTNP3. Appl. Microbiol. Biotechnol. 2012, 93, 1315–1324. [Google Scholar] [CrossRef] [Green Version]
- Kolvenbach, B.A.; Lenz, M.; Benndorf, D.; Rapp, E.; Fousek, J.; Vlcek, C.; Schäffer, A.; Gabriel, F.L.; Kohler, H.P.E.; Corvini, P.F. Purification and characterization of hydroquinone dioxygenase from Sphingomonas sp. strain TTNP3. AMB Express 2011, 1, 8. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene ID | Gene Description | Synonym | qPCR | Log2FC | |||
|---|---|---|---|---|---|---|---|
| q1 | q2 | q3 | A3 vs. A1 | A3 vs. A2 | |||
| GE00504 | Cytochrome P450 130 | P450 | 1.48 | 1.60 | 1.92 | 2.337 | |
| GE00283 | Multicopper oxidase | MCO | 1.93 | 2.21 | 2.95 | 0.64 | 4.28 |
| GE03924 | 4-hydroxyacetophenone monooxygenase | HAPMO | 1.64 | 2.10 | 2.57 | 1.027 | 2.273 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tian, K.; Yu, Y.; Qiu, Q.; Sun, X.; Meng, F.; Bi, Y.; Gu, J.; Wang, Y.; Zhang, F.; Huo, H. Mechanisms of BPA Degradation and Toxicity Resistance in Rhodococcus equi. Microorganisms 2023, 11, 67. https://doi.org/10.3390/microorganisms11010067
Tian K, Yu Y, Qiu Q, Sun X, Meng F, Bi Y, Gu J, Wang Y, Zhang F, Huo H. Mechanisms of BPA Degradation and Toxicity Resistance in Rhodococcus equi. Microorganisms. 2023; 11(1):67. https://doi.org/10.3390/microorganisms11010067
Chicago/Turabian StyleTian, Kejian, Yue Yu, Qing Qiu, Xuejian Sun, Fanxing Meng, Yuanping Bi, Jinming Gu, Yibing Wang, Fenglin Zhang, and Hongliang Huo. 2023. "Mechanisms of BPA Degradation and Toxicity Resistance in Rhodococcus equi" Microorganisms 11, no. 1: 67. https://doi.org/10.3390/microorganisms11010067