

Comparative Genomic Analyses of New and Old World Viscerotropic Leishmanine Parasites: Further Insights into the Origins of Visceral Leishmaniasis Agents
 , , , , , ,
, , , , , , 
Abstract
:1. Introduction
2. Materials and methods
2.1. Parasites
- -
- L. (L.) infantum chagasi: MCER/BR/1981/M6445/Salvaterra/Pará State/Brazil, isolated from the viscera (liver and spleen) of its wild reservoir, the crab-eating fox Cerdocyon thous (which appeared to be in good health) [19];
- -
- L. (L.) infantum chagasi: MHOM/HD/2017/M32502/Isla del Tigre/Amapala District/Honduras, isolated from a human case of atypical dermal leishmaniasis (ADL) or nonulcerated cutaneous leishmaniasis (NUCL) [20].
2.2. Parasite Cultivation and DNA Viability
2.3. Parasite Sequencing Conditions
2.4. Genome Analysis
2.5. Ethical Approval
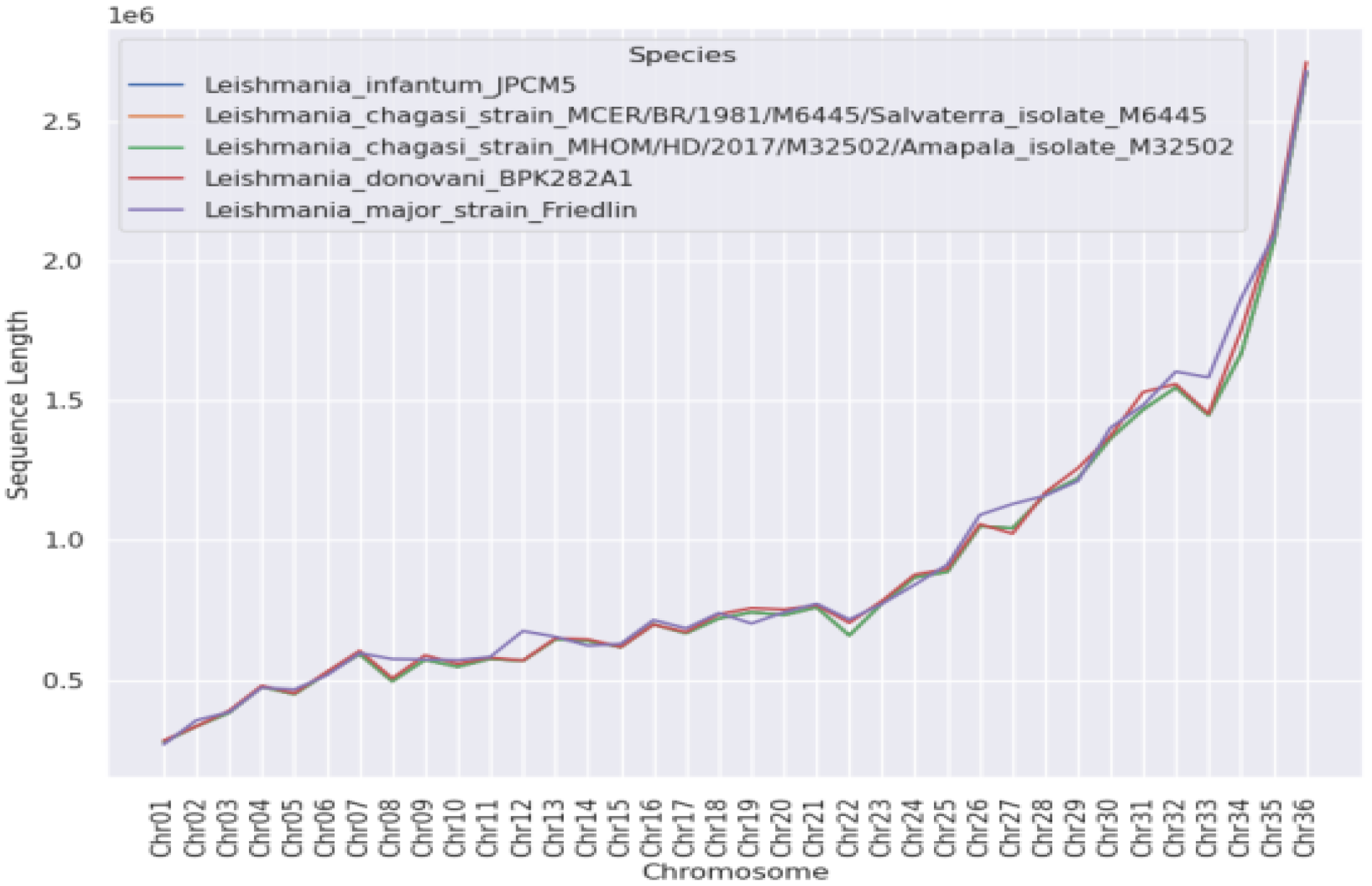
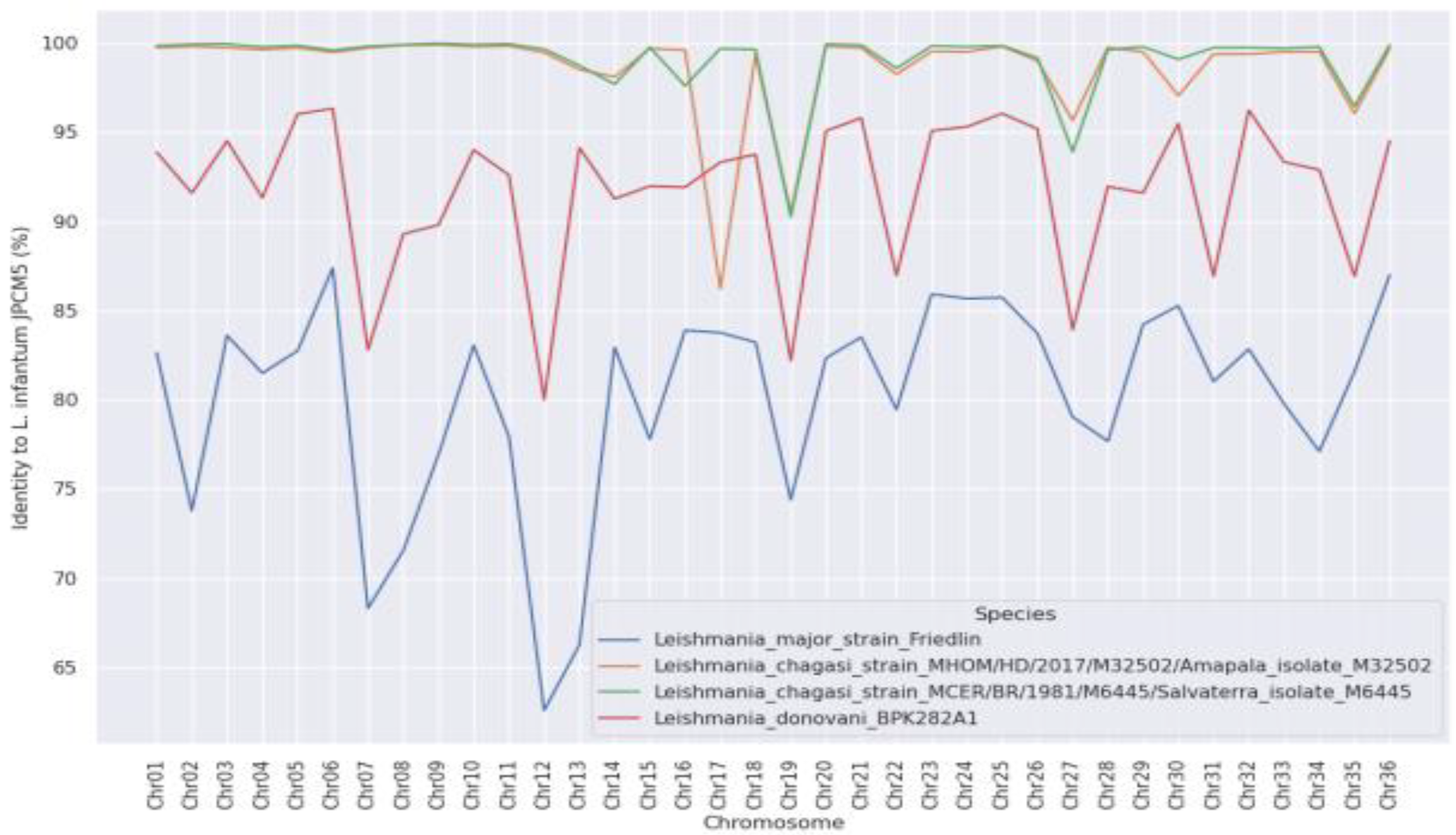
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- WHO. Leishmaniasis: World Health Organization. 2020. Available online: https://www.who.int/news-room/fact-sheets/detail/leishmaniasis (accessed on 12 January 2022).
- Alvar, J.; Vélez, I.D.; Bern, C.; Herrero, M.; Desjeux, P.; Cano, J.; Jannin, J.; den Boer, M.; WHO Leishmaniasis Control Team. Leishmaniasis worldwide and global estimates of its incidence. PLoS ONE 2012, 7, e35671. [Google Scholar] [CrossRef] [PubMed]
- Lainson, R.; Shaw, J.J. New world leishmaniasis. In Topley & Wilson’s Microbiology and Microbial Infections, 10th ed.; Collier, L., Balows, A., Sussman, M., Eds.; Arnold: London, UK, 2010; Volume 5, pp. 313–349. [Google Scholar]
- Silveira, F.T.; Lima, L.V.R.; Vasconcelos dos Santos, T.; Ramos, P.K.S.; Campos, M.C. Reviewing the trajectory of American visceral leishmaniasis in the Amazon, Brazil: From Evandro Chagas to the present days. Rev. Pan Amaz. Saúde 2016, 7, 15–22. [Google Scholar] [CrossRef]
- Lainson, R.; Shaw, J.J. Evaluation, classification and geographical distribution. In The Leishmaniases in Biology and Medicine; Peters, W., Kendrick, K., Eds.; Academic Press: London, UK, 1987; pp. 1–120. [Google Scholar]
- Cunha, A.M.; Chagas, E. Nova espécie de protozoário do gênero Leishmania patogênico para o homem. Leishmania chagasi n.sp. Nota prévia. Hospital 1937, 11, 3–9. [Google Scholar]
- Momen, H.; Grimaldi Junior, G.; Deane, L.M. Leishmania infantum, the aetiological agent of American visceral leishmaniasis (AVL)? Mem. Inst. Oswaldo Cruz. 1987, 82, 447–448. [Google Scholar] [CrossRef] [PubMed]
- Grimaldi, G., Jr.; David, J.R.; McMahon-Pratt, D. Identification and distribution of New World Leishmania species characterized by serodeme analysis using monoclonal antibodies. Am. J. Trop. Med. Hyg. 1987, 36, 270–287. [Google Scholar] [CrossRef] [PubMed]
- Kreutzer, R.D.; Souraty, N.; Semko, M.E. Biochemical identities and differences among Leishmania species and subspecies. Am. J. Trop. Med. Hyg. 1987, 36, 22–32. [Google Scholar] [CrossRef]
- Momen, H.; Pacheco, R.S.; Cupolillo, E.; Grimaldi, G., Jr. Molecular evidence for the importation of Old World Leishmania into the Americas. Biol. Res. 1993, 26, 249–255. [Google Scholar]
- Mauricio, I.L.; Howard, M.K.; Stothard, J.R.; Miles, M.A. Genomic diversity in the Leishmania donovani complex. Parasitology 1999, 119, 237–246. [Google Scholar] [CrossRef] [Green Version]
- Mauricio, I.L.; Stothard, J.R.; Miles, M.A. The strange case of Leishmania chagasi. Parasitol. Today 2000, 16, 188–189. [Google Scholar] [CrossRef]
- Lukeš, J.; Mauricio, I.L.; Schönian, G.; Dujardin, J.-C.; Soteriadou, K.; Dedet, J.-P.; Kuhls, K.; Tintaya, K.W.Q.; Jirků, M.; Chocholová, E.; et al. Evolutionary and geographical history of the Leishmania donovani complex with a revision of current taxonomy. Proc. Natl. Acad. Sci. USA 2007, 104, 9375–9380. [Google Scholar] [CrossRef] [Green Version]
- Leblois, R.; Kuhls, K.; Francois, O.; Schonian, G.; Wirth, T. Guns, germs and dogs: On the origin of Leishmania chagasi. Infect. Genet. Evol. 2011, 11, 1091–1095. [Google Scholar] [CrossRef] [PubMed]
- Kuhls, K.; Alam, M.Z.; Cupolillo, E.; Ferreira, G.E.M.; Mauricio, I.L.; Oddone, R.; Feliciangeli, M.D.; Wirth, T.; Miles, M.A.; Schönian, G. Comparative microsatellite typing of New World Leishmania infantum reveals low heterogeneity among populations and its recent Old World origin. PLoS Negl. Trop. Dis. 2011, 5, e1155. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, G.E.M.; dos Santos, B.N.; Dorval, M.E.C.; Ramos, T.P.B.; Porrozzi, R.; Peixoto, A.A.; Cupolillo, E. The genetic structure of Leishmania infantum populations in Brazil and its possible association with the transmission cycle of visceral leishmaniasis. PLoS ONE 2012, 7, e36242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Motoie, G.; Ferreira, G.E.; Cupolillo, E.; Canavez, F.; Pereira-Chioccola, V.L. Spatial distribution and population genetics of Leishmania infantum genotypes in São Paulo State, Brazil, employing multilocus microsatellite typing directly in dog infected tissues. Infect. Genet. Evol. 2013, 18, 48–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silveira, F.T.; Junior, E.C.S.; Silvestre, R.V.; Costa-Martins, A.G.; Pinheiro, K.D.C.; Ochoa, W.S.; dos Santos, T.V.; Ramos, P.K.; Casseb, S.; da Silva, S.P.; et al. Whole-genome sequencing of Leishmania infantum chagasi isolates from Honduras and Brazil. Microbiol. Res. Ann. 2021, 10, e00471-21. [Google Scholar] [CrossRef] [PubMed]
- Silveira, F.T.; Lainson, R.; Shaw, J.J.; Póvoa, M.M. Leishmaniasis in Brazil: XVIII. Further evidence incriminating the fox Cerdocyon thous (L.) as a reservoir of Amazonian visceral leishmaniasis. Trans. R. Soc. Trop. Med. Hyg. 1982, 76, 830–832. [Google Scholar] [CrossRef] [Green Version]
- Sosa-Ochoa, W.; Zúniga, C.; Chaves, L.F.; Flores, G.V.A.; Pacheco, C.M.S.; da Matta, V.L.R.; Corbett, C.E.P.; Silveira, F.T.; Laurenti, M.D. Clinical and immunological features of human Leishmania (L.) infantum-infection, novel insights Honduras, Central America. Pathogens 2020, 9, 554. [Google Scholar] [CrossRef]
- Bolger, A.M.; Marc, L.; Bjoern, U. Trimmomatic: A flexible trimmer for llumina sequence data. Bioinformatics 2014, 15, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Divorkin, M.; Kulicov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comp. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [Green Version]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [Green Version]
- Steinbiss, S.; Silva-Franco, F.; Brunk, B.; Foth, B.; Hertz-Fowler, C.; Berriman, M.; Otto, T.D. Companion: A web server for annotation and analysis of parasite genomes. Nucleic Acids Res. 2016, 44, W29–W34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waskom, M.L. Seaborn: Statistical data visualization. J. Open Source Softw. 2021, 6, 3021. [Google Scholar] [CrossRef]
- Luton, K.; Johnson, A.M. Cloning and sequence analysis of the DNA polymerase alpha gene of Leishmania donovani: Comparison with the human homologue. Biochem. Biophys. Res. Commun. 1997, 234, 95–100. [Google Scholar] [CrossRef] [PubMed]
- Akhoundi, M.; Kuhls, K.; Cannet, A.; Votýpka, J.; Marty, P.; Delaunay, P.; Sereno, D. A historical overview of the classification, evolution, and dispersion of Leishmania parasites and sandflies. PLoS Negl. Trop. Dis. 2016, 10, e0004349. [Google Scholar] [CrossRef]
- Valdivia, H.O.; Reis-Cunha, J.L.; Rodrigues-Luiz, G.F.; Baptista, R.P.; Baldeviano, G.C.; Gerbasi, R.V.; Dobson, D.E.; Pratlong, F.; Bastien, P.; Lescano, A.G.; et al. Comparative genomic analysis of Leishmania (Viannia) peruviana and Leishmania (Viannia) braziliensis. BMC Genom. 2015, 16, 715. [Google Scholar] [CrossRef] [Green Version]
- Valdivia, H.O.; Almeida, L.V.; Roatt, B.M.; Reis-Cunha, J.L.; Pereira, A.A.S.; Gontijo, C.; Fujiwara, R.T.; Reis, A.B.; Sanders, M.J.; Cotton, J.Á.; et al. Comparative genomics of canine-isolated Leishmania (Leishmania) amazonensis from an endemic focus of visceral leishmaniasis in Governador Valadares, southeastern Brazil. Sci. Rep. 2017, 7, 40804. [Google Scholar] [CrossRef] [Green Version]
- Herrera, G.; Hernández, C.; Ayala, M.S.; Flórez, C.; Teherán, A.A.; Ramírez, J.D. Evaluation of a Multilocus Sequence Typing (MLST) scheme for Leishmania (Viannia) braziliensis and Leishmania (Viannia) panamensis in Colombia. Parasites Vectors 2017, 10, 236. [Google Scholar] [CrossRef]
- Patino, L.H.; Muskus, C.; Muñoz, M.; Ramírez, J.D. Genomic analyses reveal moderate levels of ploidy, high heterozygosity and structural variations in a Colombian isolate of Leishmania (Leishmania) amazonensis. Acta Trop. 2019, 203, 105296. [Google Scholar] [CrossRef]
- Lainson, R.; Rangel, E.F. Lutzomyia longipalpis and the eco-epidemiology of American visceral leishmaniasis, with particular reference to Brazil—A review. Mem. Inst. Oswaldo Cruz 2005, 100, 811–827. [Google Scholar] [CrossRef]
- Vasconcelos dos Santos, T.; Ramos, P.K.S.; da Silva, F.M.M.; Alves, A.C.; Lima, L.V.R.; Campos, M.B.; Furtado, R.R.; Silveira, F.T. Presence of Lutzomyia longipalpis (Diptera: Psychodidae) and natural Leishmania (L.) infantum chagasi-infection in the wild rodent, Proechimys sp. (Rodentia: Echimydae), in the “Serra dos Carajás”, southern of Pará State, Brazil. In Proceedings of the 6th World Congress of Leishmaniasis, Poster Section, Toledo, Spain, 20 May 2016. [Google Scholar]
- Moreno, E.S.; Sabioni, L.A.; de Seixas, M.M.M.; de Souza Filho, J.A.; Marcelino, A.P.; Shimabukuro, P.H.F. Evidence of a sylvatic enzootic cycle of Leishmania infantum in the State of Amapá, Brazil. Rev. Soc. Bras. Med. Trop. 2020, 53, e20190169. [Google Scholar] [CrossRef]
- Sherlock, I.A.; Miranda, J.C.; Sadigursky, M.; Grimaldi, G. Natural infection of the opossum Didelphis albiventris (Marsupialia, Didelphidae) with Leishmania donovani in Brazil. Mem. Inst. Oswaldo Cruz 1984, 79, 511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corrector, A.; Gallego, J.F.; Tesh, R.B.; Peláez, D.; Díaz, A.; Montilla, M.; Paláu, M.T. Didelphis marsupialis, an apparent wild reservoir of Leishmania donovani chagasi in Colombia, South America. Am. J. Trop. Med. Hyg. 1989, 40, 480–486. [Google Scholar] [CrossRef] [PubMed]
- Lainson, R. The Neotropical Leishmania species: A brief historical review of their discovery, ecology and taxonomy. Rev. Pan Amaz. Saude 2010, 1, 13–38. [Google Scholar] [CrossRef]
- Figueiredo, F.B.; Gremião, I.D.; Pereira, S.A.; Fedulo, L.P.; Menezes, R.C.; Balthazar, D.A.; Schubach, T.M.P.; Madeira, M.F. First report of natural infection of a bush dog (Speothos venaticus) with Leishmania (Leishmania) chagasi in Brazil. Trans. R. Soc. Trop. Med. Hyg. 2008, 102, 200–201. [Google Scholar] [CrossRef] [PubMed]
- Dahroug, M.A.; Almeida, A.B.; Sousa, V.R.; Dutra, V.; Turbino, N.C.; Nakazato, L.; De Souza, R.L. Leishmania (Leishmania) chagasi in captive wild felids in Brazil. Trans. R. Soc. Trop. Med. Hyg. 2010, 104, 73–74. [Google Scholar] [CrossRef]
- Gómez-Hernández, C.; Bento, E.C.; Rezende-Oliveira, K.; Nascentes, G.A.N.; Barbosa, C.G.; Batista, L.R.; Tiburcio, M.G.S.; Pedrosa, A.L.; Lages-Silva, E.; Ramirez, J.D.; et al. Leishmania infection in bats from a non-endemic region of leishmaniasis in Brazil. Parasitology 2017, 144, 1980–1986. [Google Scholar] [CrossRef]
- Espinosa, O.A.; Serrano, M.G.; Camargo, E.P.; Teixeira, M.M.G.; Shaw, J.J. An appraisal of the taxonomy and nomenclature of trypanosomatids presently classified as Leishmania and Endotrypanum. Parasitology 2016, 15, 1–13. [Google Scholar] [CrossRef]
- Belli, A.A.; Miles, M.A.; Kelly, J.M. A putative Leishmania panamensis/Leishmania braziliensis hybrid is a causative agent of human cutaneous leishmaniasis in Nicaragua. Parasitology 1994, 109, 435–442. [Google Scholar] [CrossRef]
- Bañuls, A.L.; Guerrini, F.; Le Pont, F.; Barrera, C.; Espinel, I.; Guderian, R.; Echeverria, R.; Tibayrenc, M. Evidence for hybridization by multilocus enzyme electrophoresis and random amplified polymorphic DNA between Leishmania braziliensis and Leishmania panamensis/guyanensis in Ecuador. J. Eukaryot. Microbiol. 1997, 44, 408–411. [Google Scholar] [CrossRef]
- Nolder, D.; Roncal, N.; Davies, C.R.; Llanos-Ceuntas, A.; Miles, M.A. Multiple hybrid genotypes of Leishmania (Viannia) in a focus of mucocutaneous leishmaniasis. Am. J. Trop. Med. Hyg. 2007, 76, 573–578. [Google Scholar] [CrossRef]
- Jennings, Y.L.; de Souza, A.A.; Ishikawa, E.A.; Shaw, J.; Lainson, R.; Silveira, F.T. Phenotypic characterization of Leishmania spp. causing cutaneous leishmaniasis in the lower Amazon region, western Pará state, Brazil, reveals a putative hybrid parasite, Leishmania (Viannia) guyanensis x Leishmania (Viannia) shawi shawi. Parasite 2014, 21, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Silveira, F.T. What makes mucosal and anergic diffuse cutaneous leishmaniases so clinically and immunopathogically different? A review in Brazil. Trans. R. Soc. Trop. Med. Hyg. 2019, 113, 505–516. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, A.L.; Williams, P.; Melo, M.N.; Mayrink, W. Susceptibility of laboratory-reared female Lutzomyia longipalpis (Lutz & Neiva, 1912) to infection by different species and strains of Leishmania Ross, 1903. Mem. Inst. Oswaldo Cruz 1990, 85, 453–458. [Google Scholar] [PubMed] [Green Version]
- Gontijo, C.M.; Falcão, A.R.; Falcão, A.L.; Coelho, M.V. The development of species of Leishmania Ross, 1903 in Lutzomyia longipalpis (Lutz & Neiva, 1912). Mem. Inst. Oswaldo Cruz 1995, 90, 367–373. [Google Scholar] [PubMed]
- Volf, P.; Peckova, J. Sand flies and Leishmania: Specific versus permissive vectors. Trends Parasitol. 2007, 23, 91–92. [Google Scholar] [CrossRef] [Green Version]
- Savani, E.S.; Nunes, V.L.; Galati, E.A.; Castilho, T.M.; Zampieri, R.A.; Floeter-Winter, L.M. The finding of Lutzomyia almerioi and Lutzomyia longipalpis naturally infected by Leishmania spp. in a cutaneous and canine visceral leishmaniases focus in Serra da Bodoquena, Brazil. Vet. Parasitol. 2009, 160, 18–24. [Google Scholar] [CrossRef]
- Paiva, B.R.; Oliveira, A.G.; Dorval, M.E.; Galati, E.A.; Malafronte, R.S. Species-specific identification of Leishmania in naturally infected sand flies captured in Mato Grosso do Sul State, Brazil. Acta Trop. 2010, 115, 126–130. [Google Scholar] [CrossRef]
- Rêgo, F.D.; Rugani, J.M.; Shimabukuro, P.H.; Tonelli, G.B.; Quaresma, P.F.; Gontijo, C.M. Molecular detection of Leishmania in phlebotomine sand flies (Diptera: Psychodidae) from a cutaneous leishmaniasis focus at Xakriaba Indigenous Reserve, Brazil. PLoS ONE 2015, 10, e0122038. [Google Scholar] [CrossRef]
- Saraiva, L.; Leite, C.G.; Lima, A.C.V.M.D.R.; De Carvalho, L.O.A.; Pereira, A.A.S.; Rugani, J.M.N.; Rêgo, F.D.; Gontijo, C.M.F.; Andrade, J.D. Seasonality of sand flies (Diptera: Psychodidae) and Leishmania DNA detection in vector species in an area with endemic visceral leishmaniasis. Mem. Inst. Oswaldo Cruz 2017, 112, 309–318. [Google Scholar] [CrossRef] [Green Version]
- Montoya-Lerma, J.; Palacios, R.; Osorio, L.; Jaramillo, C.; Cadena, H. Further evidence of humans as source of Leishmania Viannia for sandflies. Mem. Inst. Oswaldo Cruz 1998, 93, 735–736. [Google Scholar] [CrossRef] [Green Version]
- Marcili, A.; Sperança, M.A.; da Costa, A.P.; Madeira, M.F.; Soares, H.S.; Sanches, C.O.C.C.; Acostam, I.C.L.; Girotto, A.; Minervino, A.H.H.; Horta, M.C.; et al. Phylogenetic relationships of Leishmania species based on trypanosomatid barcode (SSU rDNA) and gGAPDH genes: Taxonomic revision of L. (L.) infantum chagasi in South America. Infect. Genet. Evol. 2014, 25, 44–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silveira, F.T.; Corbett, C.E.P. Leishmania chagasi Cunha & Chagas, 1937: Indigenous or introduced? A brief review. Rev. Pan Amaz. Saúde 2010, 1, 143–147. [Google Scholar] [CrossRef]
- Silveira, F.T.; Lainson, R.; de Souza, A.A.A.; Crescente, J.A.B.; Corbett, C.E.P. Leishmaniose visceral americana. In RNG Leão, Medicina Tropical e Infectologia na Amazônia, v. 2, 1st ed.; Samauma Editorial, Instituto Evandro Chagas: Belém, Brazil, 2013; pp. 1245–1274. [Google Scholar]
- Teixeira, D.; Monteiro, G.; Martins, D.; Fernandes, M.; Macedo-Silva, V.; Ansaldi, M.; Nascimento, P.; Kurtz, M.; Streit, J.; Ximenes, M.; et al. Comparative analyses of whole genome sequences of Leishmania infantum isolates from humans and dogs in northeastern Brazil. Int. J. Parasitol. 2017, 47, 655–665. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peacock, C.S.; Seeger, K.; Harris, D.; Murphy, L.; Ruiz, J.C.; Quail, M.A.; Peters, N.; Adlem, E.; Tivey, A.; Aslett, M.; et al. Comparative genomic analysis of three Leishmania species that cause diverse human disease. Nat. Genet. 2007, 39, 839–847. [Google Scholar] [CrossRef] [Green Version]
- Carrillo, J.; Chinchilla, M.; Valverde, B.; Porras, O.; Mora, L. Visceral leishmaniasis in Costa Rica: First case report. Clin. Infect. Dis. 1999, 29, 678–679. [Google Scholar] [PubMed] [Green Version]
- Zhang, W.-W.; Matlashewski, G. Screening Leishmania donovani specific genes required for visceral infection. Mol. Microbiol. 2010, 77, 505–517. [Google Scholar] [CrossRef]
- Zhang, W.; Ramasamy, G.; McCall, L.-I.; Haydock, A.; Ranasinghe, S.; Abeygunasekara, P.; Sirimanna, G.; Wickremasinghe, R.; Myler, P.; Matlashewski, G. Genetic analysis of Leishmania donovani tropism using a naturally attenuated cutaneous strain. PLoS Pathog. 2014, 10, e1004244. [Google Scholar] [CrossRef] [Green Version]
- Ponce, C.; Ponce, E.; Cruz, A.; Kreutzer, R.; McMahon Pratt, D.; Neva, F. Leishmania donovani chagasi: New clinical variant of cutaneous leishmaniasis in Honduras. Lancet 1991, 337, 67–70. [Google Scholar] [CrossRef]
- Campos-Ponce, M.; Ponce, C.; Ponce, E.; Maingon, R.D.C. Leishmania chagasi/infantum: Further investigations on Leishmania tropisms in atypical cutaneous and visceral leishmaniasis foci in Central America. Exp. Parasitol. 2005, 109, 209–219. [Google Scholar] [CrossRef]
- Kwok, P.-Y.; Chen, X. Detection of single nucleotide polymorphisms. Curr. Issues Mol. Biol. 2003, 5, 43–60. [Google Scholar]
- Noyes, H.; Chance, M.; Ponce, C.; Ponce, E.; Maingon, R. Leishmania chagasi: Genotypically similar parasites from Honduras cause both visceral and cutaneous leishmaniasis in humans. Exp. Parasitol. 1997, 85, 264–273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franssen, S.U.; Durrant, C.; Stark, O.; Moser, B.; Downing, T.; Imamura, H.; Dujardin, J.-C.; Sanders, M.J.; Mauricio, I.; A Miles, M.; et al. Global genome diversity of the Leishmania donovani complex. Genet. Genom. Microbiol. Infect. Dis. 2020, 9, e51243. [Google Scholar] [CrossRef] [PubMed]
- Harkins, K.M.; Schwartz, R.S.; Cartwright, R.A.; Stone, A.C. Phylogenomic reconstruction supports supercontinent origins for Leishmania. Infect. Genet. Evol. 2016, 38, 101–109. [Google Scholar] [CrossRef] [PubMed]
- Croan, D.G.; Morrison, D.A.; Ellis, J.T. Evolution of the genus Leishmania revealed by comparison of DNA and RNA polymerase gene sequences. Mol. Biochem. Parasitol. 1997, 89, 149–159. [Google Scholar] [CrossRef]
- Butenko, A.; Kostygov, A.Y.; Sádlová, J.; Kleschenko, Y.; Becvar, T.; Podešvová, L.; Macedo, D.H.; Žihala, D.; Lukeš, J.; Bates, P.A.; et al. Comparative genomics of Leishmania (Mundinia). BMC Genom. 2019, 20, 726. [Google Scholar] [CrossRef] [Green Version]
- Arendt, M.; Cairns, K.M.; Ballard, J.W.O.; Savolainen, P.; Axelsson, E. Diet adaptation in dog reflects spread of prehistoric agriculture. Heredity 2016, 117, 301–306. [Google Scholar] [CrossRef] [Green Version]
- Kato, H.; Gomez, E.A.; Seki, C.; Furumoto, H.; Martini-Robles, L.; Muzzio, J.; Calvopiña, M.; Velez, L.; Kubo, M.; Tabbabi, A.; et al. PCR-RFLP analyses of Leishmania species causing cutaneous and mucocutaneous leishmaniasis revealed distribution of genetically complex strains with hybrid and mito-nuclear discordance in Ecuador. PLoS Negl. Trop. Dis. 2019, 13, e0007403. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
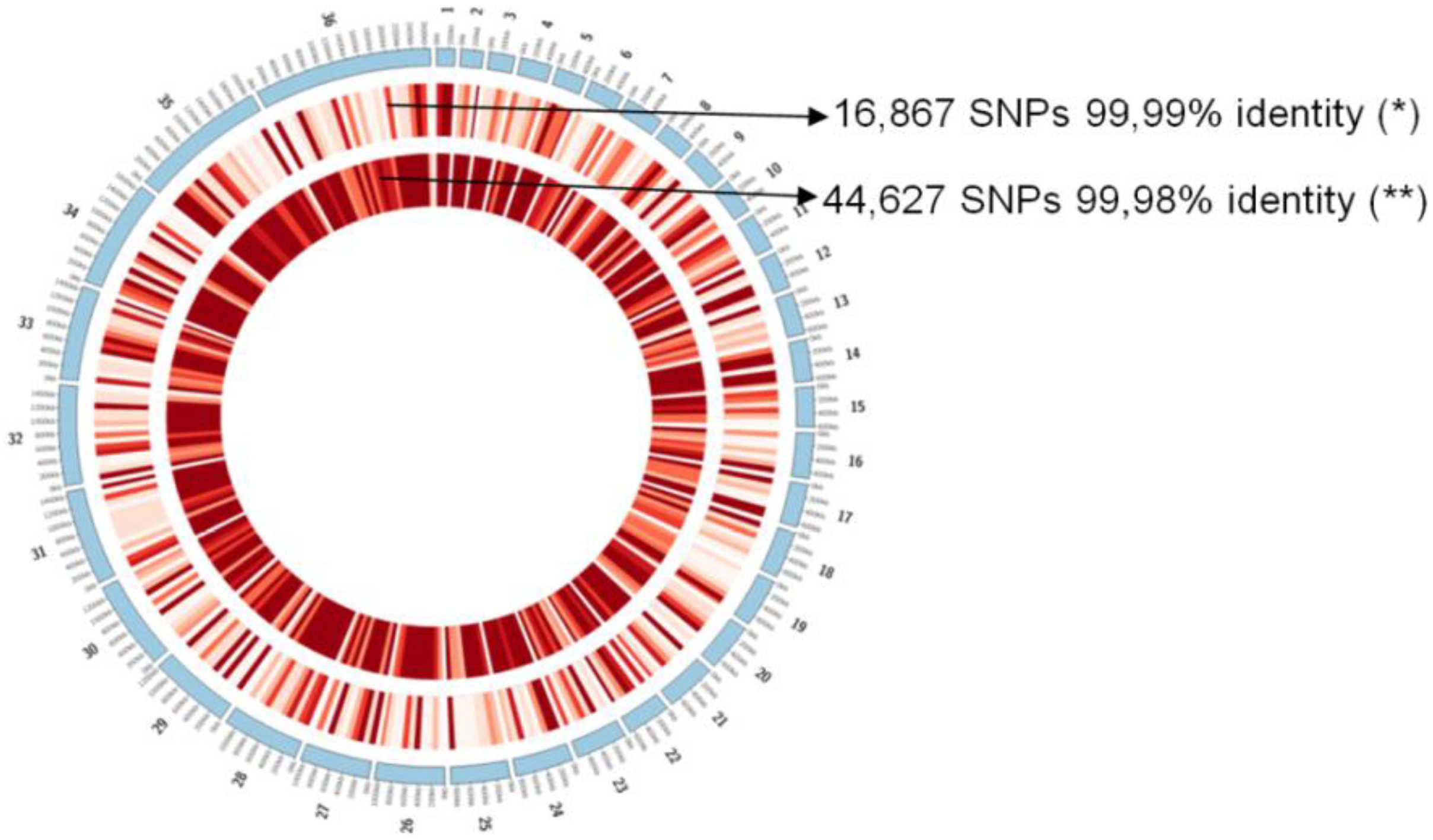
| Sample Code | No. Reads #/ Contigs † | Min. Size † (bp) | Max. Size † (bp) | N50 † (bp) | Coverage † | GC Content † (%) | Identity (%) † |
|---|---|---|---|---|---|---|---|
| MCER/BR/1981/M6445 * | 68,908,876 15,315 | 200 | 60,666 | 8354 | 130× | 59.6 | 99.99% |
| MHOM/HD/2017/M32502 ** | 25,730,242 7193 | 200 | 57,312 | 8724 | 53.47× | 59.3 | 99.98% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Silveira, F.T.; Sousa Junior, E.C.; Silvestre, R.V.D.; Vasconcelos dos Santos, T.; Sosa-Ochoa, W.; Valeriano, C.Z.; Ramos, P.K.S.; Casseb, S.M.M.; Lima, L.V.d.R.; Campos, M.B.; et al. Comparative Genomic Analyses of New and Old World Viscerotropic Leishmanine Parasites: Further Insights into the Origins of Visceral Leishmaniasis Agents. Microorganisms 2023, 11, 25. https://doi.org/10.3390/microorganisms11010025
Silveira FT, Sousa Junior EC, Silvestre RVD, Vasconcelos dos Santos T, Sosa-Ochoa W, Valeriano CZ, Ramos PKS, Casseb SMM, Lima LVdR, Campos MB, et al. Comparative Genomic Analyses of New and Old World Viscerotropic Leishmanine Parasites: Further Insights into the Origins of Visceral Leishmaniasis Agents. Microorganisms. 2023; 11(1):25. https://doi.org/10.3390/microorganisms11010025
Chicago/Turabian StyleSilveira, Fernando Tobias, Edivaldo Costa Sousa Junior, Rodrigo Vellasco Duarte Silvestre, Thiago Vasconcelos dos Santos, Wilfredo Sosa-Ochoa, Concepción Zúniga Valeriano, Patrícia Karla Santos Ramos, Samir Mansour Moraes Casseb, Luciana Vieira do Rêgo Lima, Marliane Batista Campos, and et al. 2023. "Comparative Genomic Analyses of New and Old World Viscerotropic Leishmanine Parasites: Further Insights into the Origins of Visceral Leishmaniasis Agents" Microorganisms 11, no. 1: 25. https://doi.org/10.3390/microorganisms11010025