
Serological and Molecular Survey of Leishmania infantum in a Population of Iberian Lynxes (Lynx pardinus)
 ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site and Sampled Population
2.2. Plasma and DNA Samples
2.3. Serologic Assays
2.3.1. Enzyme-Linked Immunosorbent Assay (ELISA)
2.3.2. Indirect Fluorescence Antibody Test (IFAT)
2.4. Polymerase Chain Reaction (PCR)
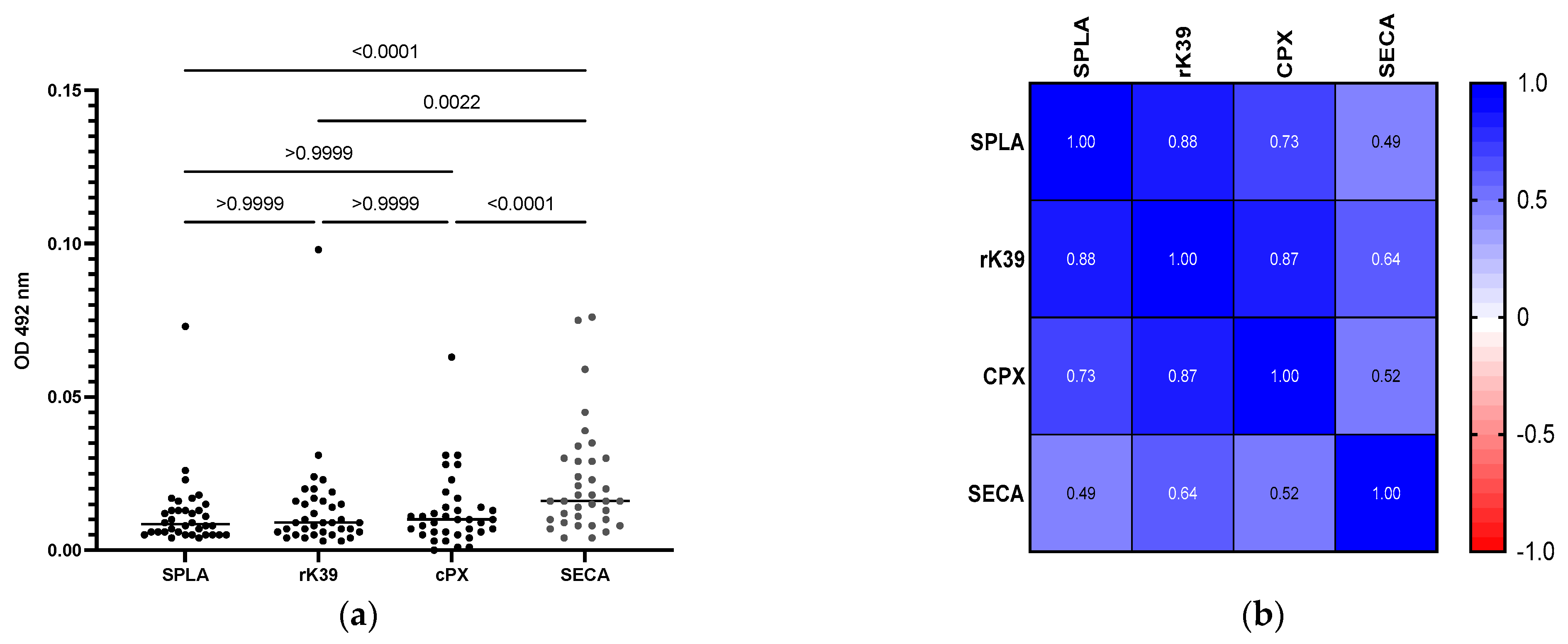
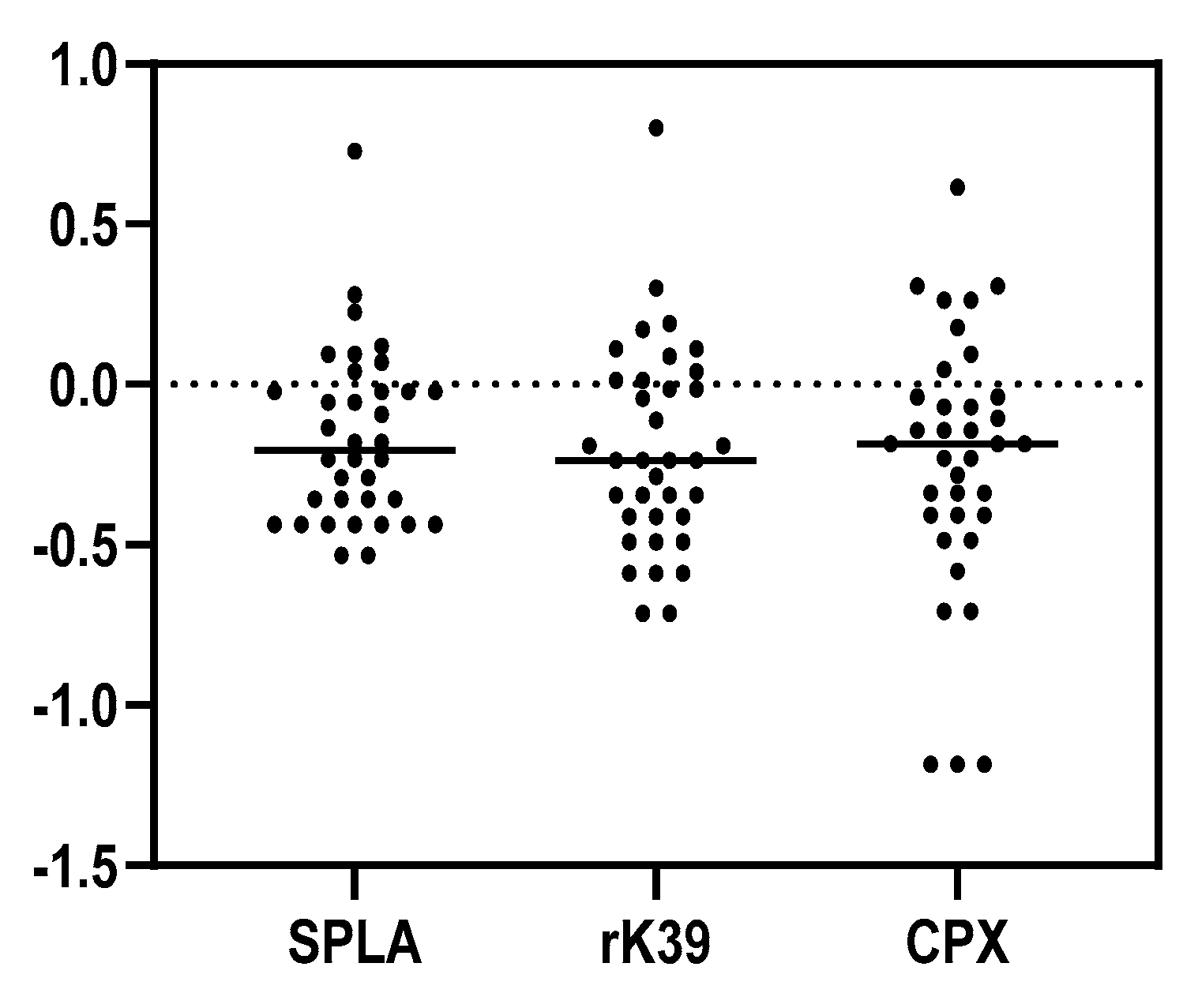
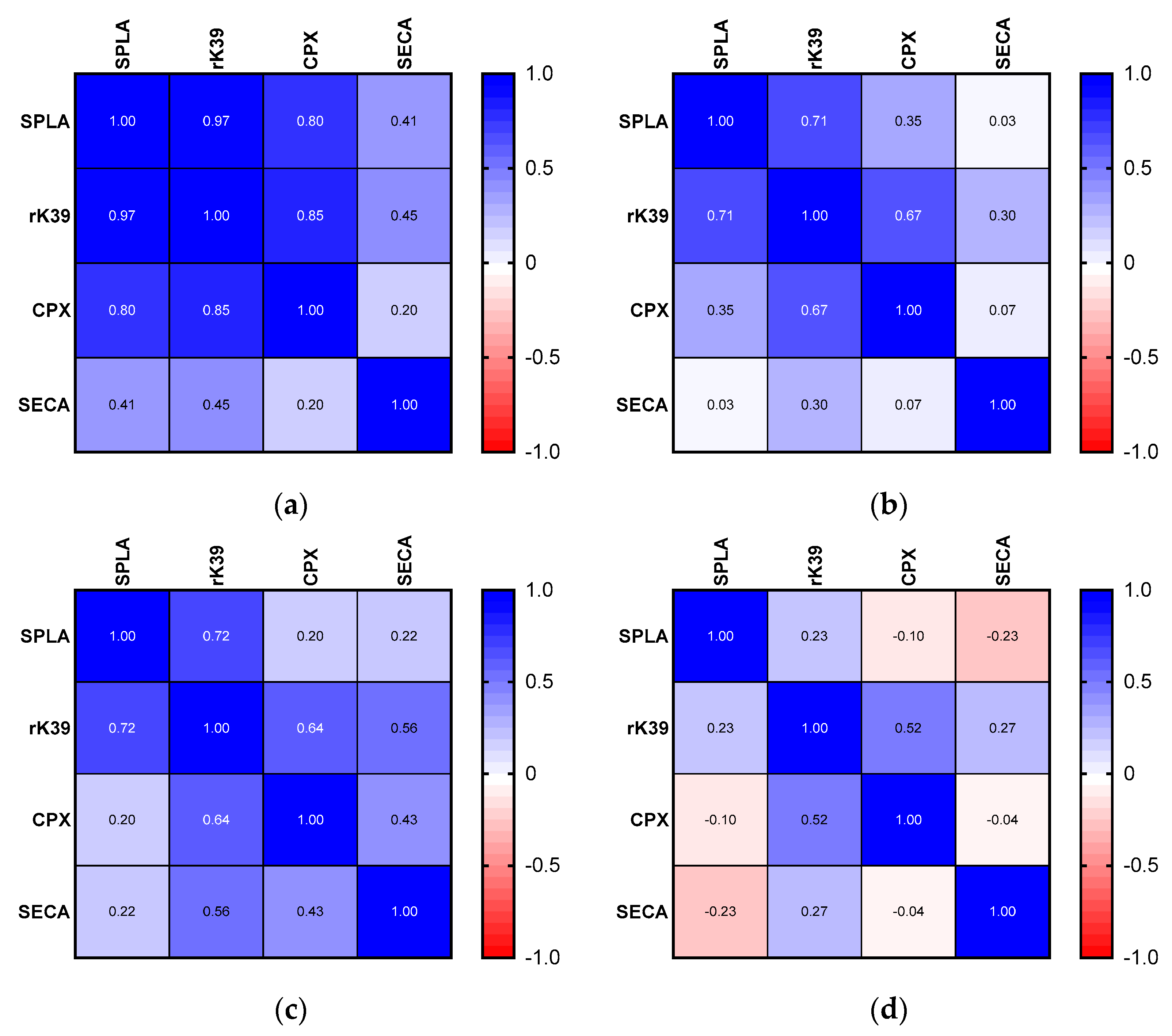
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Akuffo, H.; Costa, C.; van Griensven, J.; Burza, S.; Moreno, J.; Herrero, M. New insights into leishmaniasis in the immunosuppressed. PLoS Negl. Trop. Dis. 2018, 12, e0006375. [Google Scholar] [CrossRef] [PubMed]
- Quinell, R.J.; Courtenay, O. Transmission, reservoir hosts and control of zoonotic visceral leishmaniasis. Parasitology 2009, 136, 1915–1934. [Google Scholar] [CrossRef] [PubMed]
- Moreno, J.; Alvar, J. Canine leishmaniasis: Epidemiological risk and the experimental model. Trends Parasitol. 2002, 18, 399–405. [Google Scholar] [CrossRef]
- Miró, G.; Wright, I.; Michael, H.; Burton, W.; Hegarty, E.; Rodón, J.; Buch, J.; Pantchev, N.; von Samson-Himmelstjerna, G. Seropositivity of main vector-borne pathogens in dogs across Europe. Parasit. Vectors 2022, 15, 189. [Google Scholar] [CrossRef] [PubMed]
- Maroli, M.; Feliciangeli, M.D.; Bichaud, L.; Charrel, R.N.; Gradoni, L. Phlebotomine sandflies and the spreading of leishmaniases and other diseases of public health concern. Med. Vet. Entomol. 2013, 27, 123–147. [Google Scholar] [CrossRef]
- Maia, C.; Campino, L. Biomarkers associated with Leishmania infantum exposure, infection, and disease in dogs. Front. Cell Infect. Microbiol. 2018, 8, 302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lima, C.S.; Esteves, S.; Costa, I.; Brancal, H.; Lima, C.; Amorim, C.; Cardoso, L.; Santarém, N.; Cordeiro-da-Silva, A. Use of antigen combinations to address complex Leishmania-seropositivity patterns in Dogs living in canine leishmaniosis endemic eegions of Portugal. Microorganisms 2022, 10, 2018. [Google Scholar] [CrossRef]
- Blavier, A.; Keroack, S.; Denerolle, P.; Goy-Thollot, I.; Chabanne, L.; Cadoré, L.; Bourdoiseau, G. Atypical forms of canine leishmaniosis. Vet. J. 2001, 162, 108–120. [Google Scholar] [CrossRef]
- Santarém, N.; Sousa, S.; Amorim, C.G.; de Carvalho, N.L.; de Carvalho, H.L.; Felgueiras, O.; Brito, M.; da Silva, A.C. Challenges in the serological evaluation of dogs clinically suspect for canine leishmaniasis. Sci. Rep. 2020, 10, 3099. [Google Scholar] [CrossRef] [Green Version]
- Pereira, M.A.; Santos, R.; Nóbrega, C.; Mega, C.; Cruz, R.; Esteves, F.; Santos, C.; Coelho, C.; Mesquita, J.R.; Vala, H.; et al. A Questionnaire-based survey on the long-term management of canine leishmaniosis by veterinary practitioners. Animals 2022, 12, 731. [Google Scholar] [CrossRef]
- Velez, R.; Gállego, M. Commercially approved vaccines for canine leishmaniosis: A review of available data on their safety and efficacy. Trop. Med. Int. Health 2020, 25, 540–557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Millán, J.; Ferroglio, E.; Solano-Gallego, L. Role of wildlife in the epidemiology of Leishmania infantum infection in Europe. Parasitol. Res. 2014, 113, 2005–2014. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, L.; Schallig, H.; Persichetti, M.F.; Pennisi, M.G. New epidemiological aspects of animal leishmaniosis in Europe: The role of vertebrate hosts other than dogs. Pathogens 2021, 10, 307. [Google Scholar] [CrossRef] [PubMed]
- Vioti, G.; da Silva, M.D.; Galvis-Ovallos, F.; Alves, M.L.; da Silva, D.T.; Leonel, J.A.F.; Pereira, N.W.B.; Benassi, J.C.; Spada, J.C.P.; Maia, C.; et al. Xenodiagnosis in four domestic cats naturally infected by Leishmania infantum. Transbound. Emerg. Dis. 2021, 69, 2182–2190. [Google Scholar] [CrossRef] [PubMed]
- Batista, J.F.; Magalhães Neto, F.D.C.R.; Lopes, K.S.P.D.P.; Sato, M.O.; Costa, C.H.N.; Mendonça, I.L. Transmission of Leishmania infantum from cats to dogs. Rev. Bras. Parasitol. Vet. 2020, 29, e017820. [Google Scholar] [CrossRef]
- Rodríguez, A.; Calzada, J. Lynx pardinus (errata version published in 2020). The IUCN Red List of Threatened Species 2015. e.T12520A50655794. Available online: http://dx.doi.org/10.2305/IUCN.UK.2015-2.RLTS.T12520A50655794.en (accessed on 1 August 2022). [CrossRef]
- Instituto da Conservação da Natureza e das Florestas. Censo de lince-ibérico 2021. ICNF20JUNHO2022 2022. Available online: https://www.icnf.pt/imprensa/censodelinceiberico2021 (accessed on 1 August 2022).
- Fordham, D.A.; Akçakaya, H.R.; Brook, B.W.; Rodríguez, A.; Alves, P.C.; Civantos, E.; Triviño, M.; Watts, M.J.; Araújo, M. Adapted conservation measures are required to save the Iberian lynx in a changing climate. Nat. Clim. Chang. 2013, 3, 899–903. [Google Scholar] [CrossRef]
- Peña, L.; Garcia, P.; Jiménez, M.A.; Benito, A.; Pérez Alenza, M.; Sánchez, B. Histopathological and immunohistochemical findings in lymphoid tissues of the endangered Iberian lynx (Lynx pardinus). Comp. Immunol. Microbiol. Infect. Dis. 2006, 29, 114–126. [Google Scholar] [CrossRef]
- Rodríguez, A.; Delibes, M. Patterns and causes of non-natural mortality in the Iberian lynx during a 40-year period of range contraction. Biol. Conserv. 2004, 118, 151–161. [Google Scholar] [CrossRef]
- Millán, J.; Candela, M.G.; Palomares, F.; Cubero, M.J.; Rodríguez, A.; Barral, M.; de la Fuente, J.; Almería, S.; León-Vizcaíno, L. Disease threats to the endangered Iberian lynx (Lynx pardinus). Vet. J. 2009, 182, 114–124. [Google Scholar] [CrossRef]
- Nájera, F.; Grande-Gómez, R.; Peña, J.; Vázquez, A.; Palacios, M.J.; Rueda, C.; Corona-Bravo, A.I.; Zorrilla, I.; Revuelta, L.; Gil-Molino, M.; et al. Disease surveillance during the reintroduction of the Iberian Lynx (Lynx pardinus) in southwestern Spain. Animals 2021, 11, 547. [Google Scholar] [CrossRef]
- Pérez, J.; Calzada, J.; León-Vizcaíno, L.; Cubero, M.J.; Velarde, J.; Mozos, E. Tuberculosis in an Iberian lynx (Lynx pardina). Vet. Record 2001, 148, 414–415. [Google Scholar] [CrossRef] [PubMed]
- Briones, V. Bovine tuberculosis and the endangered Iberian lynx. Emerg. Infect. Dis. 2000, 6, 189–191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pennisi, M.G.; Persichetti, M.F. Feline leishmaniosis: Is the cat a small dog? Vet. Parasitol. 2018, 251, 131–137. [Google Scholar] [CrossRef] [PubMed]
- Meinecke, C.K.; Schottelius, J.; Oskam, L.; Fleischer, B. Congenital transmission of visceral leishmaniasis (Kala Azar) from an asymptomatic mother to her child. Pediatrics 1999, 104, e65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naucke, T.J.; Lorentz, S. First report of venereal and vertical transmission of canine leishmaniosis from naturally infected dogs in Germany. Parasit. Vectors 2012, 5, 107. [Google Scholar] [CrossRef] [Green Version]
- Reimann, M.M.; Torres-Santos, E.C.; Souza, C.S.F.; Andrade-Neto, V.V.; Jansen, A.M.; Brazil, R.P.; Roque, A.L.R. Oral and intragastric: New routes of infection by Leishmania braziliensis and Leishmania infantum? Pathogens 2022, 11, 688. [Google Scholar] [CrossRef] [PubMed]
- Tsakmakidis, Ι.; Pavlou, C.; Tamvakis, A.; Papadopoulos, T.; Christodoulou, V.; Angelopoulou, K.; Dovas, C.I.; Antoniou, M.; Anastasakis, C.; Diakou, A. Leishmania infection in lagomorphs and minks in Greece. Vet. Parasitol. Reg. Stud. Rep. 2019, 16, 100279. [Google Scholar] [CrossRef]
- Ruiz-Fons, F.; Ferroglio, E.; Gortázar, C. Leishmania infantum in free-ranging hares, Spain, 2004–2010. Eurosurveillance 2013, 18, 20541. [Google Scholar] [CrossRef] [Green Version]
- Calzolari, M.; Romeo, G.; Bergamini, F.; Dottori, M.; Rugna, G.; Carra, E. Host preference and Leishmania infantum natural infection of the sand fly Phlebotomus perfiliewi in northern Italy. Acta Trop. 2022, 226, 106246. [Google Scholar] [CrossRef]
- Priolo, V.; Masucci, M.; Donato, G.; Solano-Gallego, L.; Martínez-Orellana, P.; Persichetti, M.F.; Raya-Bermúdez, A.; Pennisi, M.G. Association between feline immunodeficiency virus and Leishmania infantum infections in cats: A retrospective matched case-control study. Parasit. Vectors 2022, 15, 107. [Google Scholar] [CrossRef]
- Grevot, A.; Jaussaud Hugues, P.; Marty, P.; Pratlong, F.; Ozon, C.; Haas, P.; Breton, C.; Bourdoiseau, G. Leishmaniosis due to Leishmania infantum in a FIV and FelV positive cat with a squamous cell carcinoma diagnosed with histological, serological and isoenzymatic methods. Parasite 2005, 12, 271–275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pimenta, P.; Alves-Pimenta, S.; Barros, J.; Barbosa, P.; Rodrigues, A.; Pereira, M.J.; Maltez, L.; Gama, A.; Cristóvão, J.M.; Campino, L.; et al. Feline leishmaniosis in Portugal: 3 cases (year 2014). Vet. Parasitol. Reg. Stud. Rep. 2015, 1–2, 65–69. [Google Scholar] [CrossRef] [PubMed]
- Casas-Marce, M.; Soriano, L.; López-Bao, J.V.; Godoy, J.A. Genetics at the verge of extinction: Insights from the Iberian lynx. Mol. Ecol. 2013, 22, 5503–5515. [Google Scholar] [CrossRef] [PubMed]
- Santarém, N.; Silvestre, R.; Cardoso, L.; Schallig, H.; Reed, S.G.; Cordeiro-da-Silva, A. Application of an improved enzyme-linked immunosorbent assay method for serological diagnosis of canine leishmaniasis. J. Clin. Microbiol. 2010, 48, 1866–1874. [Google Scholar] [CrossRef] [Green Version]
- Santarém, N.; Tomás, A.; Ouaissi, A.; Tavares, J.; Ferreira, N.; Manso, A.; Campino, L.; Correia, J.M.; Cordeiro-da-Silva, A. Antibodies against a Leishmania infantum peroxiredoxin as a possible marker for diagnosis of visceral leishmaniasis and for monitoring the efficacy of treatment. Immunol. Lett. 2005, 101, 18–23. [Google Scholar] [CrossRef]
- Lima, C.; Mesquita, J.R.; Brancal, H.; Vahlenkamp, T.; Teixeira, A.R.; Cardoso, L.; Amorim, C.; Santarém, N.; da Silva, A.C. The use of Escherichia coli total antigens as a complementary approach to address seropositivity to Leishmania antigens in canine leishmaniosis. Parasitology 2017, 144, 1384–1393. [Google Scholar] [CrossRef] [Green Version]
- Iatta, R.; Trerotoli, P.; Lucchese, L.; Natale, A.; Buonavoglia, C.; Nachum-Biala, Y.; Baneth, G.; Otranto, D. Validation of a new immunofluorescence antibody test for the detection of Leishmania infantum infection in cats. Parasitol. Res. 2020, 119, 1381–1386. [Google Scholar] [CrossRef]
- Lachaud, L.; Marchergui-Hammami, S.; Chabbert, E.; Dereure, J.; Dedet, J.P.; Bastien, P. Comparison of six PCR methods using peripheral blood for detection of canine visceral leishmaniasis. J. Clin. Microbiol. 2002, 40, 210–215. [Google Scholar] [CrossRef] [Green Version]
- Sievers, F.; Wilm, A.; Dineen, D.; Gibson, T.J.; Karplus, K.; Li, W.; Lopez, R.; McWilliam, H.; Remmert, M.; Söding, J.; et al. Fast, scalable generation of high-quality protein multiple sequence alignments using Clustal Omega. Mol. Syst. Biol. 2011, 7, 539. [Google Scholar] [CrossRef]
- Sobrino, R.; Ferroglio, E.; Oleaga, A.; Romano, A.; Millan, J.; Revilla, M.; Arnal, M.C.; Trisciuoglio, A.; Gortázar, C. Characterization of widespread canine leishmaniasis among wild carnivores from Spain. Vet. Parasitol. 2008, 155, 198–203. [Google Scholar] [CrossRef]
- Chitimia, L.; Muñoz-García, C.I.; Sánchez-Velasco, D.; Lizana, V.; Del Río, L.; Murcia, L.; Fisa, R.; Riera, C.; Giménez-Font, P.; Jiménez-Montalbán, P.; et al. Cryptic leishmaniosis by Leishmania infantum, a feature of canines only? A study of natural infection in wild rabbits, humans and dogs in southeastern Spain. Vet. Parasitol. 2011, 181, 12–16. [Google Scholar] [CrossRef] [PubMed]
- Kuhls, K.; Mauricio, I.; Pratlong, F.; Presber, H.; Schönian, G. Analysis of ribosomal DNA internal transcribed spacer sequences of the Leishmania donovani complex. Microbes Infect. 2005, 7, 1224–1234. [Google Scholar] [CrossRef]
- Solano-Gallego, L.; Rodríguez-Cortés, A.; Iniesta, L.; Quintana, J.; Pastor, J.; Espada, Y.; Portús, M.; Alberola, J. Cross-sectional serosurvey of feline leishmaniasis in ecoregions around the northwestern Mediterranean. Am. J. Trop. Med. Hyg. 2007, 76, 676–680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Da Silveira Neto, S.; Sobrinho, L.S.; Martins, C.O.; Machado, R.Z.; Marcondes, M.; de Lima, V.M. Use of crude, FML and rK39 antigens in ELISA to detect anti-Leishmania spp. antibodies in Felis catus. Vet. Parasitol. 2011, 177, 374–377. [Google Scholar] [CrossRef]
- Alves-Martin, M.F.; Paixão, M.S.; da Silva, D.T.; Tenório, M.S.; Alves, M.L.; Starke-Buzetti, W.A.; Pereira, V.B.R.; Lucheis, S.B. Detection of Leishmania spp. using parasitological, serological and molecular assays in asymptomatic and sick cats from an endemic area of visceral leishmaniosis in Brazil. Asian Pac. J. Trop. Dis. 2017, 7, 659–664. [Google Scholar] [CrossRef]
- Persichetti, M.F.; Solano-Gallego, L.; Vullo, A.; Masucci, M.; Marty, P.; Delaunay, P.; Vitale, F.; Pennisi, M.G. Diagnostic performance of ELISA, IFAT and Western blot for the detection of anti-Leishmania infantum antibodies in cats using a Bayesian analysis without a gold standard. Parasit. Vectors 2017, 10, 119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alcover, M.M.; Basurco, A.; Fernandez, A.; Riera, C.; Fisa, R.; Gonzalez, A.; Verde, M.; Garrido, A.M.; Ruíz, H.; Yzuel, A.; et al. A cross-sectional study of Leishmania infantum infection in stray cats in the city of Zaragoza (Spain) using serology and PCR. Parasit. Vectors 2021, 14, 178. [Google Scholar] [CrossRef] [PubMed]
- Zanette, M.F.; Lima, V.M.; Laurenti, M.D.; Rossi, C.N.; Vides, J.P.; Vieira, R.F.; Biondo, A.W.; Marcondes, M. Serological cross-reactivity of Trypanosoma cruzi, Ehrlichia canis, Toxoplasm gondii, Neospora caninum and Babesia canis to Leishmania infantum chagasi tests in dogs. Rev. Soc. Bras. Med. Trop. 2014, 47, 105–107. [Google Scholar] [CrossRef]
- Marcelino, A.P.; Filho, J.A.S.; Bastos, C.V.; Ribeiro, S.R.; Medeiros, F.A.C.; Reis, I.A.; Lima, A.C.V.M.D.R.; Barbosa, J.R.; Paz, G.F.; Gontijo, C.M.F. Comparative PCR-based diagnosis for the detection of Leishmania infantum in naturally infected dogs. Acta Trop. 2020, 207, 105495. [Google Scholar] [CrossRef]
- Manna, L.; Vitale, F.; Reale, S.; Caracappa, S.; Pavone, L.M.; Morte, R.D.; Cringoli, G.; Staiano, N.; Gravino, A.E. Comparison of different tissue sampling for PCR-based diagnosis and follow-up of canine visceral leishmaniosis. Vet. Parasitol. 2004, 125, 251–262. [Google Scholar] [CrossRef]
- Guerbouj, S.; Mkada-Driss, I.; Guizani, I. Molecular for Understanding Eco-Epidemiology, Diversity and Pathogenesis of Leishmania Parasites in Leishmaniasis–Trends in Epidemiology, Diagnosis and Treatment; IntechOpen: London, UK, 2014. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Diagnostic Tool | Total Analyzed N | Total Positive n | % Positive |
|---|---|---|---|
| SPLA | 36 | 9 | 25.0 |
| SPLA + rK39 | 36 | 8 | 22.2 |
| SPLA + CPX | 36 | 8 | 22.2 |
| SPLA + rK39 + CPX | 36 | 8 | 22.2 |
| SPLA + IFAT | 36 | 1 | 2.8 |
| SPLA + rK39 + IFAT | 36 | 1 | 2.8 |
| SPLA + rK39 + CPX + IFAT | 36 | 1 | 2.8 |
| SPLA + PCR | 27 | 2 | 7.4 |
| SPLA + rK39 + PCR | 27 | 2 | 7.4 |
| SPLA + rK39 + CPX + PCR | 27 | 2 | 7.4 |
| SPLA + IFAT + PCR | 27 | 1 | 3.7 |
| SPLA + rK39 + IFAT + PCR | 27 | 1 | 3.7 |
| SPLA + rK39 + CPX + IFAT + PCR | 27 | 1 | 3.7 |
| rK39 | 36 | 10 | 27.8 |
| rK39 + CPX | 36 | 8 | 22.2 |
| rK39 + IFAT | 36 | 1 | 2.8 |
| rK39 + CPX + IFAT | 36 | 1 | 2.8 |
| rK39 + PCR | 27 | 2 | 7.4 |
| rK39 + CPX + PCR | 27 | 2 | 7.4 |
| rK39 + IFAT + PCR | 27 | 1 | 3.7 |
| rK39 + CPX + IFAT + PCR | 27 | 1 | 3.7 |
| CPX | 36 | 8 | 22.2 |
| CPX + IFAT | 36 | 1 | 2.8 |
| CPX + PCR | 27 | 2 | 7.4 |
| CPX + IFAT + PCR | 27 | 1 | 3.7 |
| IFAT | 36 | 4 | 11.1 |
| PCR | 35 | 8 | 22.9 |
| IFAT + PCR | 27 | 3 | 11.1 |
| Associations between Diagnostic Tools | Total n | Overall % |
|---|---|---|
| ELISA antigens | ||
| SPLA + rK39 | 18/36 | 50.0 |
| SPLA + CPX | 17/36 | 47.2 |
| rK39 + CPX | 18/36 | 50 |
| SPLA + rK39 + CPX | 16/36 | 44.4 |
| IFAT and ELISA antigens | ||
| IFAT + SPLA | 4/36 | 11.11 |
| IFAT + rK39 | 3/36 | 8.33 |
| IFAT + CPX | 3/36 | 8.33 |
| IFAT + SPLA + rk39 | 3/36 | 8.33 |
| IFAT + SPLA + CPX | 3/36 | 8.33 |
| IFAT + rK39 + CPX | 3/36 | 8.33 |
| IFAT + SPLA + rK39 + CPX | 3/36 | 8.33 |
| PCR and ELISA antigens | ||
| SPLA + PCR | 6/27 | 22.2 |
| rK39 + PCR | 6/27 | 22.2 |
| CPX + PCR | 6/27 | 22.2 |
| SPLA + rK39 + PCR | 6/27 | 22.2 |
| SPLA + CPX + PCR | 6/27 | 22.2 |
| rK39 + CPX + PCR | 6/27 | 22.2 |
| SPLA + rK39 + CPX + PCR | 6/27 | 22.2 |
| PCR and IFAT | ||
| IFAT + PCR | 3/27 | 11.1 |
| All diagnostic tools | ||
| SPLA + rK39 + CPX + IFAT + PCR | 3/27 | 11.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lima, C.M.; Santarém, N.; Neves, N.C.; Sarmento, P.; Carrapato, C.; de Sousa, R.; Cardoso, L.; Cordeiro-da-Silva, A. Serological and Molecular Survey of Leishmania infantum in a Population of Iberian Lynxes (Lynx pardinus). Microorganisms 2022, 10, 2447. https://doi.org/10.3390/microorganisms10122447
Lima CM, Santarém N, Neves NC, Sarmento P, Carrapato C, de Sousa R, Cardoso L, Cordeiro-da-Silva A. Serological and Molecular Survey of Leishmania infantum in a Population of Iberian Lynxes (Lynx pardinus). Microorganisms. 2022; 10(12):2447. https://doi.org/10.3390/microorganisms10122447
Chicago/Turabian StyleLima, Clara M., Nuno Santarém, Nuno Costa Neves, Pedro Sarmento, Carlos Carrapato, Rita de Sousa, Luís Cardoso, and Anabela Cordeiro-da-Silva. 2022. "Serological and Molecular Survey of Leishmania infantum in a Population of Iberian Lynxes (Lynx pardinus)" Microorganisms 10, no. 12: 2447. https://doi.org/10.3390/microorganisms10122447