
Spatio-Temporal Patterns of Ticks and Molecular Survey of Anaplasma marginale, with Notes on Their Phylogeny


 , ,
, ,  ,
,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethical Approval
2.2. Study Area
2.3. Tick Collection and Preservation
2.4. Morphological Identification of Ticks
2.5. DNA Extraction and PCR
2.6. DNA Sequencing and Phylogenetic Analysis
3. Results
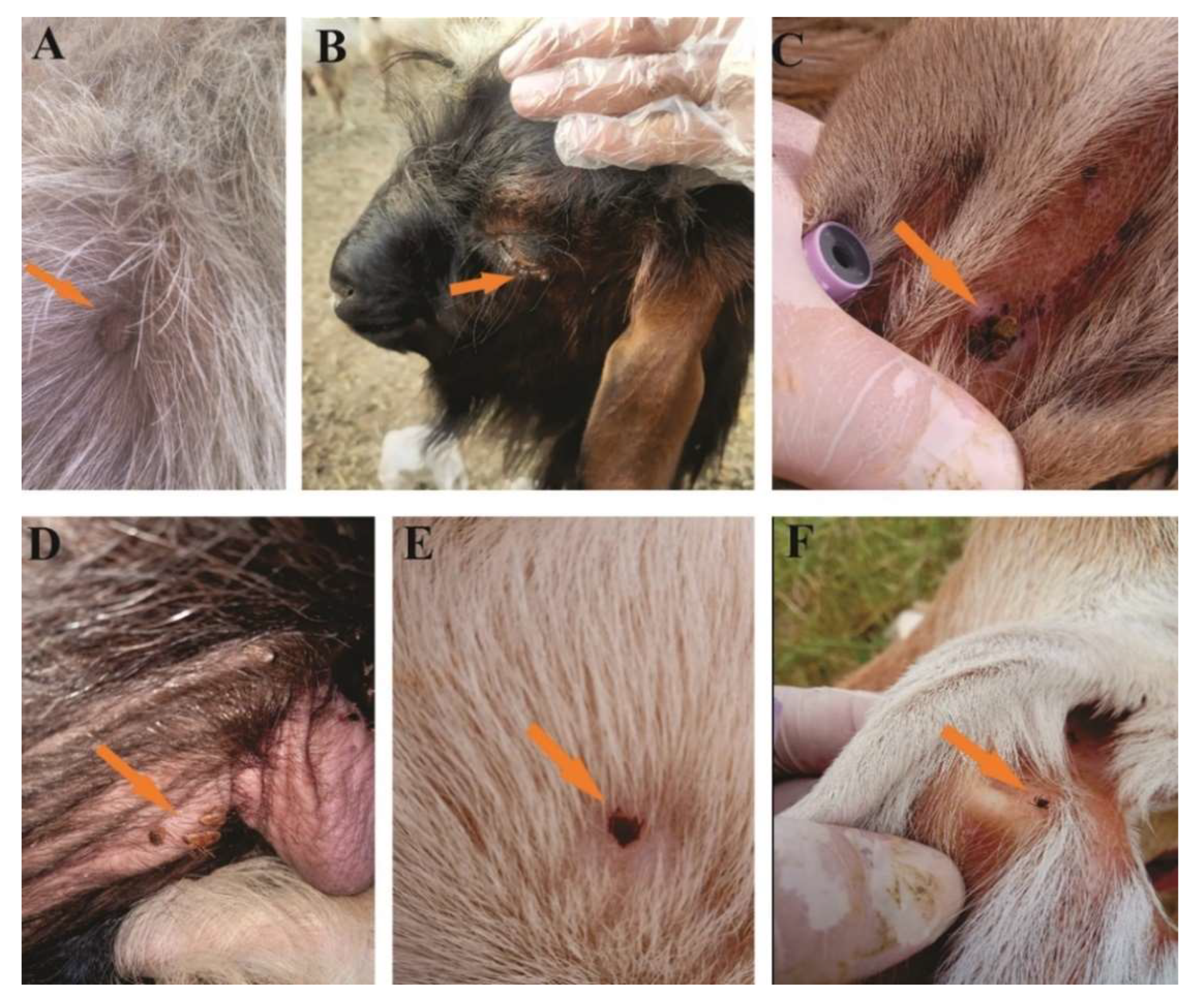
3.1. Morphologically Identified Ticks
3.2. Prevalence of Ticks
3.3. Spatial Pattern of Ticks
3.4. Seasonal Abundance of Ticks
3.5. Detection of Anaplasma spp. in Ticks
3.6. Sequencing Analysis
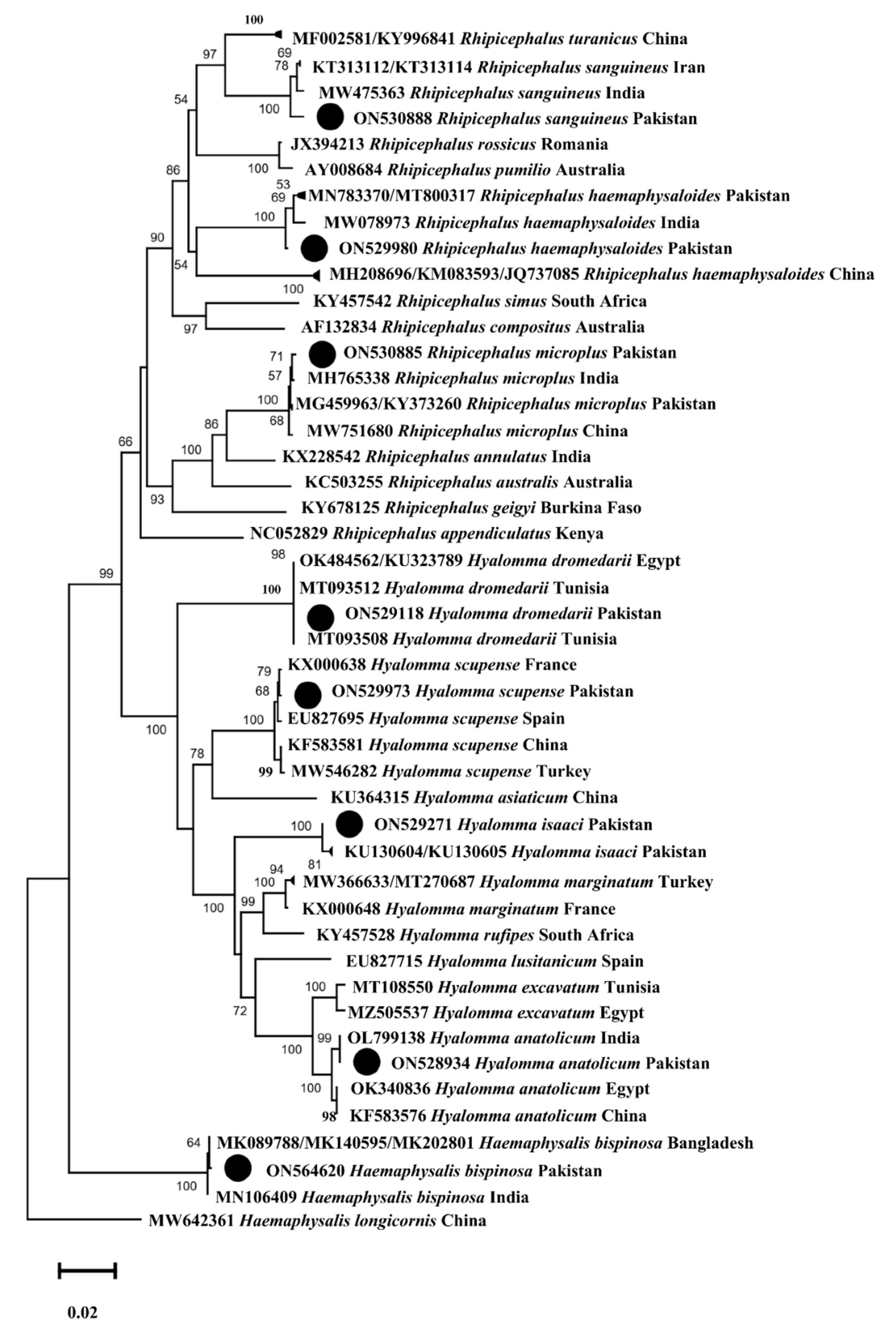
3.7. Phylogenetic Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sonenshine, D.E.; Roe, R.M. Biology of Ticks; Oxford University Press: Oxford, UK, 2013; Volume 2. [Google Scholar]
- Karim, S.; Budachetri, K.; Mukherjee, N.; Williams, J.; Kausar, A.; Hassan, M.J.; Adamson, S.; Dowd, S.E.; Apanskevich, D.; Arijo, A. A study of ticks and tick-borne livestock pathogens in Pakistan. PLoS Negl. Trop. Dis. 2017, 11, e0005681. [Google Scholar] [CrossRef] [PubMed]
- Ali, A.; Khan, M.A.; Zahid, H.; Yaseen, P.M.; Qayash Khan, M.; Nawab, J.; Ur Rehman, Z.; Ateeq, M.; Khan, S.; Ibrahim, M. Seasonal dynamics, record of ticks infesting humans, wild and domestic animals, and molecular phylogeny of Rhipicephalus microplus in Khyber Pakhtunkhwa Pakistan. Front. Physiol. 2019, 10, 793. [Google Scholar] [CrossRef] [PubMed]
- Boulanger, N.; Boyer, P.; Talagrand-Reboul, E.; Hansmann, Y. Ticks and tick-borne diseases. Med. Mal. Infect. 2019, 49, 87–97. [Google Scholar] [CrossRef]
- Eckstein, D.; Künzel, V.; Schäfer, L. Global Climate Risk Index 2021. Who Suffers Most from Extreme Weather Events; Germanwatch: Bonn, Germany, 2021; pp. 2000–2019. [Google Scholar]
- Hoogstraal, H.; Varma, M.G.R. Haemaphysalis cornupunctata sp. n. and H. kashmirensis sp. n. from Kashmir, with Notes on H. sundrai Sharif and H. sewelli Sharif of India and Pakistan (Ixodoidea, Ixodidae). J. Parasitol. 1962, 48, 185–194. [Google Scholar] [CrossRef]
- Ali, A.; Mulenga, A.; Vaz, I.S., Jr. Tick and tick-borne pathogens: Molecular and immune targets for control strategies. Front. Physiol. 2020, 11, 744. [Google Scholar] [CrossRef]
- Rehman, A.; Nijhof, A.M.; Sauter-Louis, C.; Schauer, B.; Staubach, C.; Conraths, F.J. Distribution of ticks infesting ruminants and risk factors associated with high tick prevalence in livestock farms in the semi-arid and arid agro-ecological zones of Pakistan. Parasites Vectors 2017, 10, 190. [Google Scholar] [CrossRef] [PubMed]
- Ali, A.; Zahid, H.; Zeb, I.; Tufail, M.; Khan, S.; Haroon, M.; Bilal, M.; Hussain, M.; Alouffi, A.S.; Muñoz-Leal, S.; et al. Risk factors associated with tick infestations on equids in Khyber Pakhtunkhwa, Pakistan, with notes on Rickettsia massiliae detection. Parasites Vectors 2021, 14, 363. [Google Scholar] [CrossRef] [PubMed]
- Aiman, O.; Ullah, S.; Chitimia-Dobler, L.; Nijhof, A.M.; Ali, A. First report of Nosomma monstrosum ticks infesting Asian water buffaloes (Bubalus bubalis) in Pakistan. Ticks Tick-Borne Dis. 2022, 13, 101899. [Google Scholar] [CrossRef]
- Ali, A.; Shehla, S.; Zahid, H.; Ullah, F.; Zeb, I.; Ahmed, H.; da Silva Vaz, I.; Tanaka, T. Molecular survey and spatial distribution of Rickettsia spp. in ticks infesting free-ranging wild animals in Pakistan (2017–2021). Pathogens 2022, 11, 162. [Google Scholar] [CrossRef]
- Ali, A.; Numan, M.; Khan, M.; Aiman, O.; Muñoz-Leal, S.; Chitimia-Dobler, L.; Labruna, M.B.; Nijhof, A.M. Ornithodoros (Pavlovskyella) ticks associated with a Rickettsia sp. in Pakistan. Parasites Vectors 2022, 15, 138. [Google Scholar] [CrossRef]
- Begum, F.; Wisseman, C.L., Jr.; Casals, J. Tick-borne viruses of West Pakistan: II. Hazara virus, a new agent isolated from Ixodes redikorzevi ticks from the Kaghan valley, W. Pakistan. Am. J. Epidemiol. 1970, 92, 192–194. [Google Scholar] [CrossRef] [PubMed]
- Clifford, C.M.; Hoogstraal, H.; Kohls, G.M. Ixodes hyatti, n. sp.; and I. shahi, n. sp. (Acarina: Ixodidae), parasites of Pikas (Lagomorpha: Ochotonidae) in the Himalayas of Nepal and West Pakistan. J. Med. Entomol. 1971, 8, 430–438. [Google Scholar] [CrossRef] [PubMed]
- Auffenberg, W.; Auffenberg, T. The reptile tick Aponomma gervaisi: (Acarina Ixodidae) as a parasite of monitor lizards in Pakistan and India. Bull. Fla. Mus. Nat. Hist. Biol. Sci. 1990, 35, 1–34. [Google Scholar]
- McCarthy, V.C. Ixodid Ticks (Acarina, Ixodidae) of West Pakistan. Doctoral dissertation, University of Maryland, College Park, MD, USA, 1967; p. 533. [Google Scholar]
- Jabbar, A.; Abbas, T.; Sandhu, Z.U.D.; Saddiqi, H.A.; Qamar, M.F.; Gasser, R.B. Tick-borne diseases of bovines in Pakistan: Major scope for future research and improved control. Parasites Vectors 2015, 8, 283. [Google Scholar] [CrossRef]
- Rehman, A.; Conraths, F.J.; Sauter-Louis, C.; Krücken, J.; Nijhof, A.M. Epidemiology of tick-borne pathogens in the semi-arid and the arid agro-ecological zones of Punjab province, Pakistan. Transbound. Emerg. Dis. 2019, 66, 526–536. [Google Scholar] [CrossRef] [PubMed]
- Aktas, M.; Altay, K.; Dumanli, N. Molecular detection and identification of Anaplasma and Ehrlichia species in cattle from Turkey. Ticks Tick-Borne Dis. 2011, 2, 62–65. [Google Scholar] [CrossRef] [PubMed]
- Raoult, D.; Parola, P. (Eds.) Rickettsial Diseases; Informa Healthcare: New York, NY, USA, 2007; p. 379. [Google Scholar]
- Ybañez, A.P.; Ybanez, R.H.D.; Claveria, F.G.; Cruz-Flores, M.J.; Xuenan, X.; Yokoyama, N.; Inokuma, H. High genetic diversity of Anaplasma marginale detected from Philippine cattle. J. Vet. Med. Sci. 2014, 76, 1009–1014. [Google Scholar] [CrossRef]
- Begum, F.; Wisseman, C.L., Jr.; Traub, R. Tick-borne viruses of West Pakistan: I. Isolation and general characteristics. Am. J. Epidemiol. 1970, 92, 180–191. [Google Scholar] [CrossRef]
- Ghafar, A.; Gasser, R.B.; Rashid, I.; Ghafoor, A.; Jabbar, A. Exploring the prevalence and diversity of bovine ticks in five agro-ecological zones of Pakistan using phenetic and genetic tools. Ticks Tick-Borne Dis. 2020, 11, 101472. [Google Scholar] [CrossRef]
- Khan, Z.; Shehla, S.; Alouffi, A.; Kashif Obaid, M.; Zeb Khan, A.; Almutairi, M.M.; Numan, M.; Aiman, O.; Alam, S.; Ullah, S.; et al. Molecular survey and genetic characterization of Anaplasma marginale in ticks collected from livestock hosts in Pakistan. Animals 2022, 12, 1708. [Google Scholar] [CrossRef]
- Hoogstraal, H.; Kaiser, M.N. Observations on Egyptian Hyalomma ticks (Ixodoidea, Ixodidae). Biological notes and differences in identity of H. anatolicum and its subspecies anatolicum Koch and excavatum Koch among Russian and other workers. Identity of H. lusitanicum Koch. Ann. Entomol. Soc. Am. 1959, 52, 243–261. [Google Scholar] [CrossRef]
- Hoogstraal, H.; Trapido, H.; Kohls, G.M. Studies on southeast Asian Haemaphysalis ticks (Ixodoidea, Ixodidae). Speciation in the H. (Kaiseriana) obesa group: H. semermis Neumann, H. obesa Larrousse, H. roubaudi Toumanoff, H. montgomeryi Nuttall, and H. hirsuta sp. n. J. Parasitol. 1966, 52, 169–191. [Google Scholar] [CrossRef] [PubMed]
- Hoogstraal, H.; Trapido, H. Redescription of the type materials of Haemaphysalis (Kaiseriana) bispinosa Neumann (India), H. (K.) neumanni Dönitz (Japan), H. (K.) lagrangei Larrousse (Vietnam), and H. (K.) yeni Toumanoff (Vietnam) (Ixodoidea, Ixodidae). J. Parasitol. 1966, 52, 1188–1198. [Google Scholar] [CrossRef]
- Apanaskevich, D.A. Differentiation of closely related species Hyalomma anatolicum and H. excavatum (Acari: Ixodidae) based on a study of all life cycle stages, throughout entire geographical range. Parazitologiia 2003, 37, 259–280. [Google Scholar] [PubMed]
- Walker, A.R. Ticks of Domestic Animals in Africa: A Guide to Identification of Species; Bioscience Reports: Edinburgh, UK, 2003; pp. 3–210. [Google Scholar]
- Walker, J.B.; Keirans, J.E.; Horak, I.G. The Genus Rhipicephalus (Acari, Ixodidae): A Guide to the Brown Ticks of the World; Cambridge University Press: Cambridge, UK, 2005. [Google Scholar]
- Apanaskevich, D.A.; Horak, I.G. The genus Hyalomma Koch, 1844: V. Re-evaluation of the taxonomic rank of taxa comprising the H. (Euhyalomma) marginatum Koch complex of species (Acari: Ixodidae) with redescription of all parasitic stages and notes on biology. Int. J. Acarol. 2008, 34, 13–42. [Google Scholar] [CrossRef]
- Apanaskevich, D.A.; Schuster, A.L.; Horak, I.G. The genus Hyalomma: VII. Redescription of all parasitic stages of H. (Euhyalomma) dromedarii and H. (E.) schulzei (Acari: Ixodidae). J. Med. Entomol. 2008, 45, 817–831. [Google Scholar] [CrossRef]
- Apanaskevich, D.A.; Filippova, N.A.; Horak, I.G. The genus Hyalomma Koch, 1844. X. Redescription of all parasitic stages of H. (Euhyalomma) scupense Schulze, 1919 (= H. detritum Schulze) (Acari: Ixodidae) and notes on its biology. Folia Parasitol. 2010, 57, 69–78. [Google Scholar] [CrossRef]
- Sambrook, J.; Fritsch, E.F.; Maniatis, T. Molecular Cloning: A Laboratory Manual, 2nd ed.; Cold Spring Harbor Laboratory Press: New York, NY, USA, 1989. [Google Scholar]
- Chitimia, L.; Lin, R.Q.; Cosoroaba, I.; Wu, X.Y.; Song, H.Q.; Yuan, Z.G.; Zhu, X.Q. Genetic characterization of ticks from southwestern Romania by sequences of mitochondrial cox1 and nad5 genes. Exp. App. Acarol. 2010, 52, 305–311. [Google Scholar] [CrossRef]
- Mangold, A.J.; Bargues, M.D.; Mas-Coma, S. Mitochondrial 16S rDNA sequences and phylogenetic relationships of species of Rhipicephalus and other tick genera among Metastriata (Acari: Ixodidae). Parasitol. Res. 1998, 84, 478–484. [Google Scholar] [CrossRef]
- Hailemariam, Z.; Ahmed, J.S.; Clausen, P.H.; Nijhof, A.M. A comparison of DNA extraction protocols from blood spotted on FTA cards for the detection of tick-borne pathogens by Reverse Line Blot hybridization. Ticks Tick-Borne Dis. 2017, 8, 185–189. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic Local Alignment Search Tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Hall, T.; Biosciences, I.; Carlsbad, C. BioEdit: An important software for molecular biology. GERF Bull. Biosci. 2011, 2, 60–61. [Google Scholar]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA-X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Pakistan Economic Survey. Government of Pakistan. Ministry of Finance, Islamabad. Available online: https://www.finance.gov.pk/survey (accessed on 2 October 2021).
- MacDonald, A.J. Abiotic and habitat drivers of tick vector abundance, diversity, phenology and human encounter risk in southern California. PLoS ONE 2018, 13, e0201665. [Google Scholar] [CrossRef] [PubMed]
- Uspensky, I. Preliminary observations on specific adaptations of exophilic ixodid ticks to forests or open country habitats. Exp. App. Acarol. 2002, 28, 147–154. [Google Scholar] [CrossRef] [PubMed]
- Kamran, K.; Ali, A.; Villagra, C.A.; Bazai, Z.A.; Iqbal, A.; Sajid, M.S. Hyalomma anatolicum resistance against ivermectin and fipronil is associated with indiscriminate use of acaricides in southwestern Balochistan, Pakistan. Parasitol. Res. 2021, 120, 15–25. [Google Scholar] [CrossRef]
- Palmer, G.H.; Rurangirwa, F.R.; McElwain, T.F. Strain composition of the Ehrlichia Anaplasma marginale within persistently infected cattle, a mammalian reservoir for tick transmission. J. Clin. Microbiol. 2001, 39, 631–635. [Google Scholar] [CrossRef]
- Hegab, A.A.; Omar, H.M.; Abuowarda, M.; Ghattas, S.G.; Mahmoud, N.E.; Fahmy, M.M. Screening and phylogenetic characterization of tick-borne pathogens in a population of dogs and associated ticks in Egypt. Parasites Vectors 2022, 15, 1–15. [Google Scholar] [CrossRef]
- Parker, R.J. The Australian brown dog tick Rhipicephalus sanguineus as an experimental parasite of cattle and vector of Anaplasma marginale. Aust. Vet. J. 1982, 58, 47–50. [Google Scholar] [CrossRef]
- Shkap, V.; Kocan, K.; Molad, T.; Mazuz, M.; Leibovich, B.; Krigel, Y.; Michoytchenko, A.; Blouin, E.; De la Fuente, J.; Samish, M.; et al. Experimental transmission of field Anaplasma marginale and the A. centrale vaccine strain by Hyalomma excavatum, Rhipicephalus sanguineus and Rhipicephalus (Boophilus) annulatus ticks. Vet. Microbiol. 2009, 134, 254–260. [Google Scholar] [CrossRef]
- Quiroz-Castañeda, R.E.; Amaro-Estrada, I.; Rodríguez-Camarillo, S.D. Anaplasma marginale: Diversity, virulence, and vaccine landscape through a genomics approach. BioMed Res. Int. 2016, 2016, 9032085. [Google Scholar] [CrossRef]
- Krasnov, B.R.; Mouillot, D.; Shenbrot, G.I.; Khokhlova, I.S.; Vinarski, M.V.; Korallo-Vinarskaya, N.P.; Poulin, R. Similarity in ectoparasite faunas of Palearctic rodents as a function of host phylogenetic, geographic, or environmental distances: Which matters the most? Int. J. Parasitol. 2010, 40, 807–817. [Google Scholar] [CrossRef] [PubMed]
- Vatansever, Z. Hyalomma anatolicum Koch, 1844 (Figs. 158–160). In Ticks of Europe and North Africa; Springer: Cham, Switzerland, 2017; pp. 391–395. [Google Scholar]
- Lv, J.; Wu, S.; Zhang, Y.; Chen, Y.; Feng, C.; Yuan, X.; Jia, G.; Deng, J.; Wang, C.; Wang, Q.; et al. Assessment of four DNA fragments (COI, 16S rDNA, ITS2, 12S rDNA) for species identification of the Ixodida (Acari: Ixodida). Parasites Vectors 2014, 7, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Beati, L.; Klompen, H. Phylogeography of ticks (Acari: Ixodida). Ann. Rev. Entomol. 2019, 64, 379–397. [Google Scholar] [CrossRef] [PubMed]
- Patel, J.B. 16S rRNA gene sequencing for bacterial pathogen identification in the clinical laboratory. Mol Diagn. 2001, 6, 313–321. [Google Scholar] [CrossRef]
- Janda, J.M.; Abbott, S.L. 16S rRNA gene sequencing for bacterial identification in the diagnostic laboratory: Pluses, perils, and pitfalls. J. Clin. Microbiol. 2007, 45, 2761–2764. [Google Scholar] [CrossRef]
- Geevarghese, G.; Mishra, A.C. Haemaphysalis Ticks of India; Elsevier: Amsterdam, The Netherlands, 2011. [Google Scholar]
- Sands, A.F.; Apanaskevich, D.A.; Matthee, S.; Horak, I.G.; Harrison, A.; Karim, S.; Mohammad, M.K.; Mumcuoglu, K.Y.; Rajakaruna, R.S.; Santos-Silva, M.M.; et al. Effects of tectonics and large-scale climatic changes on the evolutionary history of Hyalomma ticks. Mol. Phylogenet. Evol. 2017, 114, 153–165. [Google Scholar] [CrossRef]
- Pomerantzev, B.I. Ticks (fam. Ixodidae) of the USSR and neighboring countries. Opred. Faune SSSR Zool. Inst. Akad. Nauk SSSR 1946, 26, 26–28. (in Russian). [Google Scholar]
- Hoogstraal, H.; Kaiser, M.N. Observations on ticks (Ixodoidea) of Libya. Ann. Entomol. Soc. Am. 1960, 53, 445–457. [Google Scholar] [CrossRef]
- Kaiser, M.N.; Hoogstraal, H. The Hyalomma ticks (Ixodoidea, Ixodidae) of Afghanistan. J. Parasitol. 1963, 49, 130–139. [Google Scholar] [CrossRef]
- Bakkes, D.K.; Ropiquet, A.; Chitimia-Dobler, L.; Matloa, D.E.; Apanaskevich, D.A.; Horak, I.G.; Mans, B.J.; Matthee, C.A. Adaptive radiation and speciation in Rhipicephalus ticks: A medley of novel hosts, nested predator-prey food webs, off-host periods, and dispersal along temperature variation gradients. Mol. Phylogenet. Evol. 2021, 162, 107178. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Zheng, Y.C.; Ma, L.; Jia, N.; Jiang, B.G.; Jiang, R.R.; Huo, Q.B.; Wang, Y.W.; Liu, H.B.; Chu, Y.L.; et al. Human infection with a novel tick-borne Anaplasma species in China: A surveillance study. Lancet Infect. Dis. 2015, 15, 663–670. [Google Scholar] [CrossRef]
- Low, V.L.; Tay, S.T.; Kho, K.L.; Koh, F.X.; Tan, T.K.; Lim, Y.A.L.; Ong, B.L.; Panchadcharam, C.; Norma-Rashid, Y.; Sofian-Azirun, M. Molecular characterization of the tick Rhipicephalus microplus in Malaysia: New insights into the cryptic diversity and distinct genetic assemblages throughout the world. Parasites Vectors 2015, 8, 341. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Organism/Gene | Sequence (5′-3′) | Amplicon Size | PCR Condition | Ref. |
|---|---|---|---|---|
| Ticks/cox1 | cox1 F, GGAACAATATATTTAATTTTTGG cox1 R, ATCTATCCCTACTGTAAATATATG | 801 bp | 95 °C 5 min, 40× (95 °C 30 s, 55 °C 60 s, 72 °C 1 min), 72 °C 5 min | [35] |
| Ticks/16S rRNA | 16S+1, CCGGTCTGAACTCAGATCAAGT 16S−1, GCTCAATGATTTTTTAAATTGCTG | 460 bp | 95 °C 3 min, 40× (95 °C 30 s, 55 °C 60 s, 72 °C 1 min), 72 °C 7 min | [36] |
| Anaplasma spp./16S rRNA | Ehr-F2, AGAGTTTGATCCTGGCTCAG Ehr-R, AGTTTGCCGGGACTTYTTCT | 1100 bp | 95 °C 3 min, 35× (95 °C 30 s, 50 °C 30 s, 72 °C 1 min), 72 °C 7 min | [37] |
| Tick Species | Tick Life Stages | Total Ticks (%) | Ticks Subjected to PCR | Infested Hosts | Anaplasma marginale | |||
|---|---|---|---|---|---|---|---|---|
| Female • (%) | Male (%) | Nymph (%) | Positive Ticks | Infection Rate % | ||||
| Hy. dromedarii | 187 (47.9) | 170 (43.6) | 33 (8.5) | 390 (20.8) | 45 | Camels, Sheep, Cattle | 14 | 31.1 |
| Hy. anatolicum | 140 (47.6) | 128 (43.5) | 26 (8.9) | 294 (15.7) | 42 | Cattle, Sheep, Goats, Dogs, Asian water buffaloes, Horses, Camels | 10 | 23.8 |
| Hy. scupense | 103 (49.7) | 86 (41.6) | 18 (8.7) | 207 (11.0) | 33 | Cattle, Asian water buffaloes, Horses | 9 | 27.3 |
| Hy. isaaci | 33 (56.9) | 19 (32.8) | 6 (10.3) | 58 (3.1) | 15 | Sheep, Cattle, Goats | 0 | 0 |
| Ha. cornupunctata | 51 (47.7) | 42 (39.3) | 14 (13) | 107 (5.7) | 8 | Sheep, Goats | 0 | 0 |
| Ha. montgomeryi | 42 (48.3) | 36 (41.4) | 9 (10.3) | 87 (4.6) | 6 | Goats, Sheep | 0 | 0 |
| Ha. bispinosa | 26 (48.2) | 18 (33.3) | 10 (18.5) | 54 (2.9) | 8 | Goats, Sheep | 0 | 0 |
| Ha. sulcata | 21 (44.7) | 17 (36.2) | 9 (19.1) | 47 (2.5) | 7 | Sheep, Goats | 0 | 0 |
| R. microplus | 126 (48.1) | 78 (29.8) | 58 (22.1) | 262 (14) | 40 | Cattle, Asian water buffaloes, Sheep, Goats, Dogs | 12 | 30 |
| R. turanicus | 61 (50.4) | 51 (42.2) | 9 (7.4) | 121 (6.5) | 6 | Sheep, Goats, Dogs, Horses | 0 | 0 |
| R. sanguineus | 71 (52.2) | 52 (38.2) | 13 (9.6) | 136 (7.3) | 30 | Dogs, Sheep, Goats | 6 | 20 |
| R. haemaphysaloides | 53 (48.2) | 45 (40.9) | 12 (10.9) | 110 (5.9) | 30 | Dogs, Sheep, Goats | 6 | 20 |
| Total | 914 (48.8) | 742 (39.6) | 217 (11.6) | 1873 | 314 | 54 | 17.2 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alam, S.; Khan, M.; Alouffi, A.; Almutairi, M.M.; Ullah, S.; Numan, M.; Islam, N.; Khan, Z.; Aiman, O.; Zaman Safi, S.; et al. Spatio-Temporal Patterns of Ticks and Molecular Survey of Anaplasma marginale, with Notes on Their Phylogeny. Microorganisms 2022, 10, 1663. https://doi.org/10.3390/microorganisms10081663
Alam S, Khan M, Alouffi A, Almutairi MM, Ullah S, Numan M, Islam N, Khan Z, Aiman O, Zaman Safi S, et al. Spatio-Temporal Patterns of Ticks and Molecular Survey of Anaplasma marginale, with Notes on Their Phylogeny. Microorganisms. 2022; 10(8):1663. https://doi.org/10.3390/microorganisms10081663
Chicago/Turabian StyleAlam, Shumaila, Mehran Khan, Abdulaziz Alouffi, Mashal M. Almutairi, Shafi Ullah, Muhammad Numan, Nabila Islam, Zaibullah Khan, Ome Aiman, Sher Zaman Safi, and et al. 2022. "Spatio-Temporal Patterns of Ticks and Molecular Survey of Anaplasma marginale, with Notes on Their Phylogeny" Microorganisms 10, no. 8: 1663. https://doi.org/10.3390/microorganisms10081663