
Epidemiology and Traits of Mobile Colistin Resistance (mcr) Gene-Bearing Organisms from Horses

 ,
,  ,
,  and
and 
Abstract
:1. Introduction
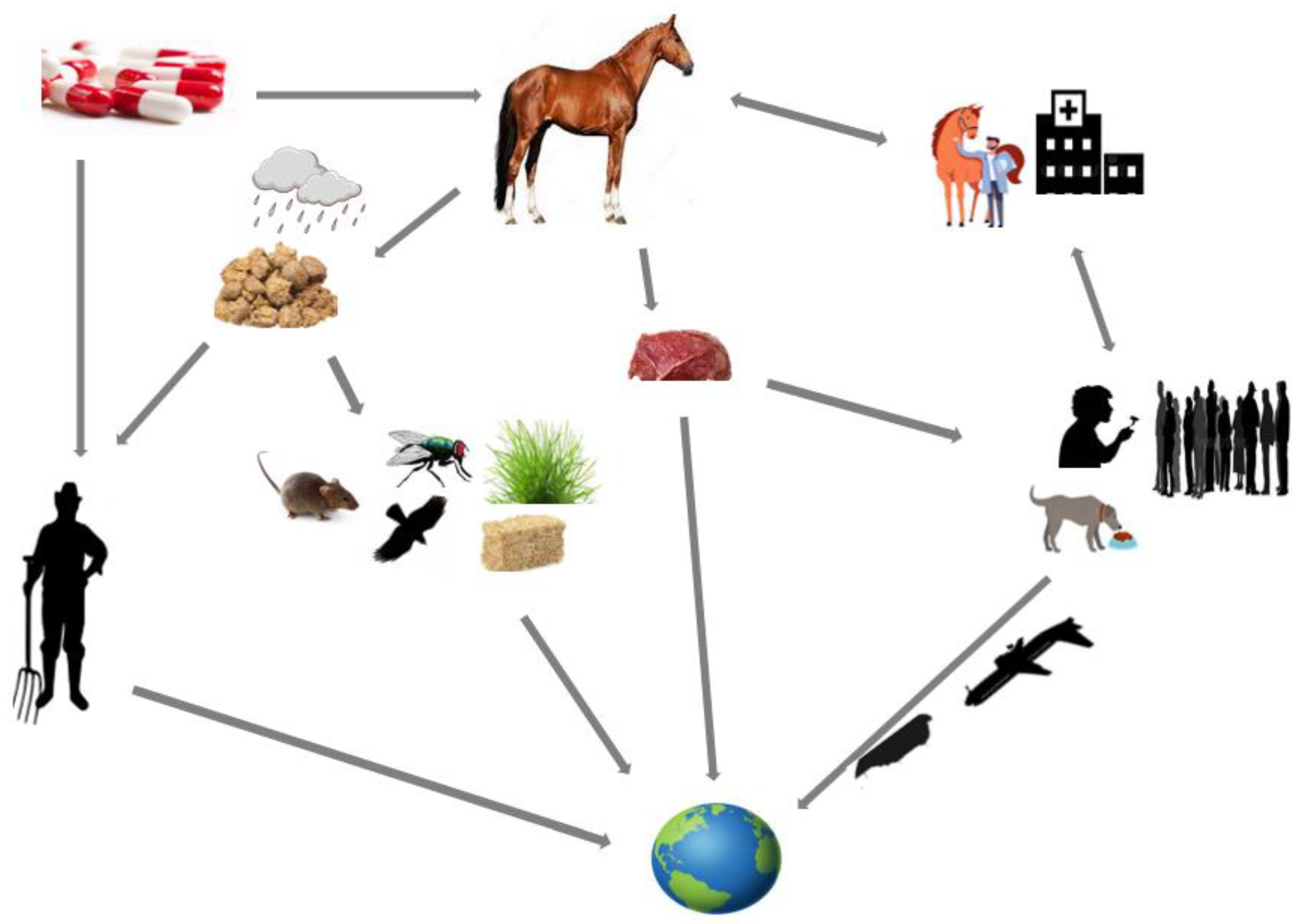
2. Possible Sources, Transmission Routes, and Impacts of mcr-Gene-Bearing Organisms in Horses
3. Epidemiology and Traits of mcr-Gene-Bearing Bacteria Isolated from Horses in Different World Regions
3.1. Africa
3.2. Europe
3.3. South America
3.4. North America
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Murray, C.J.; Ikuta, K.S.; Sharara, F.; Swetschinski, L.; Robles Aguilar, G.; Gray, A.; Han, C.; Bisignano, C.; Rao, P.; Wool, E.; et al. Global Burden of Bacterial Antimicrobial Resistance in 2019: A Systematic Analysis. Lancet 2022, 399, 629–655. [Google Scholar] [CrossRef]
- Kakoullis, L.; Papachristodoulou, E.; Chra, P.; Panos, G. Mechanisms of Antibiotic Resistance in Important Gram-Positive and Gram-Negative Pathogens and Novel Antibiotic Solutions. Antibiotics 2021, 10, 415. [Google Scholar] [CrossRef] [PubMed]
- Barriere, S.L. Expert Opinion on Pharmacotherapy Clinical, Economic and Societal Impact of Antibiotic Resistance. Expert Opin. Pharmacother. 2015, 16, 151–153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhen, X.; Lundborg, C.S.; Sun, X.; Hu, X.; Dong, H. The Clinical and Economic Impact of Antibiotic Resistance in China: A Systematic Review and Meta-Analysis. Antibiotics 2019, 8, 115. [Google Scholar] [CrossRef] [Green Version]
- Founou, R.C.; Founou, L.L.; Essack, S.Y. Clinical and Economic Impact of Antibiotic Resistance in Developing Countries: A Systematic Review and Meta-Analysis. PLoS ONE 2017, 12, e0189621. [Google Scholar]
- Ahmad, M.; Khan, A.U. Global Economic Impact of Antibiotic Resistance: A Review. J. Glob. Antimicrob. Resist. 2019, 19, 313–316. [Google Scholar] [CrossRef]
- Anyanwu, M.U.; Anyogu, D.C.; Chah, K.F.; Shoyinka, V.S. Mobile Colistin Resistance (Mcr-1) Gene-Positive Escherichia coli from Chickens in Nigeria Is Potentially Pathogenic and Transfers Colistin Resistance to Other Organisms. Comp. Clin. Pathol. 2022, 31, 323–332. [Google Scholar] [CrossRef]
- Partridge, S.R.; Kwong, S.M.; Firth, N.; Jensen, S.O. Mobile Genetic Elements Associated with Antimicrobial Resistance. Clin. Microbiol. Rev. 2018, 31, e00088-17. [Google Scholar] [CrossRef] [Green Version]
- Vrancianu, C.O.; Popa, L.I.; Bleotu, C.; Chifiriuc, M.C. Targeting Plasmids to Limit Acquisition and Transmission of Antimicrobial Resistance. Front. Microbiol. 2020, 11, 761. [Google Scholar] [CrossRef]
- Rozwandowicz, M.; Brouwer, M.S.M.; Fischer, J.; Wagenaar, J.A.; Gonzalez-Zorn, B.; Guerra, B.; Mevius, D.J.; Hordijk, J. Plasmids Carrying Antimicrobial Resistance Genes in Enterobacteriaceae. J. Antimicrob. Chemother. 2018, 73, 1121–1137. [Google Scholar] [CrossRef] [Green Version]
- Poirel, L.; Jayol, A.; Nordmanna, P. Polymyxins: Antibacterial Activity, Susceptibility Testing, and Resistance Mechanisms Encoded by Plasmids or Chromosomes. Clin. Microbiol. Rev. 2017, 30, 557–596. [Google Scholar] [CrossRef] [Green Version]
- Binsker, U.; Käsbohrer, A.; Hammerl, J.A. Global Colistin Use: A Review of the Emergence of Resistant Enterobacterales and the Impact on Their Genetic Basis. FEMS Microbiol. Rev. 2022, 46, fuab049. [Google Scholar] [CrossRef] [PubMed]
- Anyanwu, M.U.; Jaja, I.F.; Nwobi, O.C. Occurrence and Characteristics of Mobile Colistin Resistance (Mcr) Gene-Containing Isolates from the Environment: A Review. Int. J. Environ. Res. Public Health 2020, 17, 1028. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khuntayaporn, P.; Thirapanmethee, K.; Chomnawang, M.T. An Update of Mobile Colistin Resistance in Non-Fermentative Gram-Negative Bacilli. Front. Cell. Infect. Microbiol. 2022, 12, 882236. [Google Scholar] [CrossRef]
- El-Sayed Ahmed, M.A.E.G.; Zhong, L.L.; Shen, C.; Yang, Y.; Doi, Y.; Tian, G.B. Colistin and Its Role in the Era of Antibiotic Resistance: An Extended Review (2000–2019). Emerg. Microbes Infect. 2020, 9, 868–885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anyanwu, M.U.; Okpala, C.O.R.; Chah, K.F.; Shoyinka, V.S. Prevalence and Traits of Mobile Colistin Resistance Gene Harbouring Isolates from Different Ecosystems in Africa. BioMed Res. Int. 2021, 2021, 6630379. [Google Scholar] [CrossRef]
- Liu, Y.Y.; Wang, Y.; Walsh, T.R.; Yi, L.X.; Zhang, R.; Spencer, J.; Doi, Y.; Tian, G.; Dong, B.; Huang, X.; et al. Emergence of Plasmid-Mediated Colistin Resistance Mechanism MCR-1 in Animals and Human Beings in China: A Microbiological and Molecular Biological Study. Lancet Infect. Dis. 2016, 16, 161–168. [Google Scholar] [CrossRef]
- Elbediwi, M.; Li, Y.; Paudyal, N.; Pan, H.; Li, X.; Xie, S.; Rajkovic, A.; Feng, Y.; Fang, W.; Rankin, S.C.; et al. Global Burden of Colistin-Resistant Bacteria: Mobilized Colistin Resistance Genes Study (1980–2018). Microorganisms 2019, 7, 461. [Google Scholar] [CrossRef] [Green Version]
- Martiny, H.-M.; Munk, P.; Brinch, C.; Szarvas, J.; Aarestrup, F.M.; Petersen, T.N. Global Distribution of Mcr Gene Variants in 214K Metagenomic Samples. mSystems 2022, 7, e00105-22. [Google Scholar] [CrossRef]
- Anyanwu, M.U.; Jaja, I.F.; Okpala, C.O.R.; Jaja, C.J.I.; Oguttu, J.W.; Chah, K.F.; Shoyinka, V.S. Potential Sources and Characteristic Occurrence of Mobile Colistin Resistance (Mcr) Gene-Harbouring Bacteria Recovered from the Poultry Sector: A Literature Synthesis Specific to High-Income Countries. PeerJ 2021, 9, e11606. [Google Scholar] [CrossRef]
- Leshaba, T.M.S.; Mbelle, N.M.; Osei Sekyere, J. Current and Emerging Polymyxin Resistance Diagnostics: A Systematic Review of Established and Novel Detection Methods. J. Appl. Microbiol. 2022, 132, 8–30. [Google Scholar] [CrossRef] [PubMed]
- Bardet, L.; Rolain, J.M. Development of New Tools to Detect Colistin-Resistance among Enterobacteriaceae Strains. Can. J. Infect. Dis. Med. Microbiol. 2018, 2018, 3095249. [Google Scholar] [CrossRef] [PubMed]
- Ling, Z.; Yin, W.; Shen, Z.; Wang, Y.; Shen, J.; Walsh, T.R. Epidemiology of Mobile Colistin Resistance Genes Mcr-1 to Mcr-9. J. Antimicrob. Chemother. 2020, 75, 3087–3095. [Google Scholar] [CrossRef] [PubMed]
- Shedko, E.D.; Timoshina, O.Y.; Azyzov, I.S. Molecular Epidemiology of Mcr Gene Group. Clin. Microbiol. Antimicrob. Chemother. 2020, 22, 287–300. [Google Scholar] [CrossRef]
- Sun, J.; Zhang, H.; Liu, Y.H.; Feng, Y. Towards Understanding MCR-like Colistin Resistance. Trends Microbiol. 2018, 26, 794–808. [Google Scholar] [CrossRef]
- Anyanwu, M.U.; Jaja, I.F.; Oguttu, J.W.; Jaja, C.J.; Chah, K.F.; Shodeinde Shoyinka, V. Is Africa Ready for Mobile Colistin Resistance Threat? Infect. Ecol. Epidemiol. 2021, 11, 1962781. [Google Scholar] [CrossRef]
- Shi, X.; Li, Y.; Yang, Y.; Shen, Z.; Wu, Y.; Wang, S. Global Impact of Mcr-1-Positive Enterobacteriaceae Bacteria on “One Health”. Crit. Rev. Microbiol. 2020, 46, 565–577. [Google Scholar]
- Valiakos, G.; Kapna, I. Colistin Resistant Mcr Genes Prevalence in Livestock Animals (Swine, Bovine, Poultry) from a Multinational Perspective. a Systematic Review. Vet. Sci. 2021, 8, 265. [Google Scholar] [CrossRef]
- Bastidas-Caldes, C.; de Waard, J.H.; Salgado, M.S.; Villacís, M.J.; Coral-Almeida, M.; Yamamoto, Y.; Calvopiña, M. Worldwide Prevalence of mcr-mediated Colistin-Resistance Escherichia coli in Isolates of Clinical Samples, Healthy Humans, and Livestock—A Systematic Review and Meta-Analysis. Pathogens 2022, 11, 659. [Google Scholar] [CrossRef]
- Sun, J.; Zeng, X.; Li, X.P.; Liao, X.P.; Liu, Y.H.; Lin, J. Plasmid-Mediated Colistin Resistance in Animals: Current Status and Future Directions. Anim. Health Res. Rev. 2017, 18, 136–152. [Google Scholar] [CrossRef]
- Nang, S.C.; Li, J.; Velkov, T. The Rise and Spread of mcr Plasmid-Mediated Polymyxin Resistance. Crit. Rev. Microbiol. 2019, 45, 131–161. [Google Scholar] [CrossRef] [PubMed]
- Agina, O.A.; Ihedioha, J.I. Occurrence of Wounds in Nigerian Horses. J. Appl. Anim. Welf. Sci. 2017, 20, 372–380. [Google Scholar] [CrossRef] [PubMed]
- Belaunzaran, X.; Bessa, R.J.B.; Lavín, P.; Mantecón, A.R.; Kramer, J.K.G.; Aldai, N. Horse-Meat for Human Consumption-Current Research and Future Opportunities. Meat Sci. 2015, 108, 74–81. [Google Scholar] [CrossRef]
- Anyanwu, M.U.; Ugwu, I.C.; Onah, C.U. Occurrence and Antibiogram of Generic Extended-Spectrum Cephalosporin-Resistant and Extended-Spectrum β-Lactamase-Producing Enterobacteria in Horses. Maced. Vet. Rev. 2018, 41, 123–132. [Google Scholar] [CrossRef] [Green Version]
- Cross, P. Global Horse Statistics Internal 02 2019; HiPoint Agro Bedding Corp.: Guelph, ON, Canada, 2019; pp. 1–19. [Google Scholar]
- Chung, Y.S.; Song, J.W.; Kim, D.H.; Shin, S.; Park, Y.K.; Yang, S.J.; Lim, S.K.; Park, K.T.; Park, Y.H. Isolation and Characterization of Antimicrobial-Resistant Escherichia coli from National Horse Racetracks and Private Horse-Riding Courses in Korea. J. Vet. Sci. 2016, 17, 199–206. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.-Y. The Horse Industry in Korea: Its Present Condition and Prospect. Adv Sci Technol Lett. 2015, 99, 256–260. [Google Scholar]
- Hardefeldt, L.Y.; Bailey, K.E.; Slater, J. Overview of the Use of Antimicrobial Drugs for the Treatment of Bacterial Infections in Horses. Equine Vet. Educ. 2021, 33, 602–611. [Google Scholar] [CrossRef]
- Isgren, C. Antimicrobial Resistance in Horses. Vet. Rec. 2018, 183, 316–318. [Google Scholar] [CrossRef] [Green Version]
- Isgren, C.M. Improving Clinical Outcomes via Responsible Antimicrobial Use in Horses. Equine Vet. Educ. 2021. [Google Scholar] [CrossRef]
- Álvarez–Narváez, S.; Berghaus, L.J.; Morris, E.R.A.; Willingham-Lane, J.M.; Slovis, N.M.; Giguere, S.; Cohen, N.D. A Common Practice of Widespread Antimicrobial Use in Horse Production Promotes Multi-Drug Resistance. Sci. Rep. 2020, 10, 911–913. [Google Scholar] [CrossRef]
- Nielsen, S.S.; Bicout, D.J.; Calistri, P.; Canali, E.; Drewe, J.A.; Garin-Bastuji, B.; Gonzales Rojas, J.L.; Gortazar Schmidt, C.; Herskin, M.; Michel, V.; et al. Assessment of Animal Diseases Caused by Bacteria Resistant to Antimicrobials: Horses. EFSA J. 2021, 19, e06955. [Google Scholar] [CrossRef]
- Schnepf, A.; Bienert-Zeit, A.; Ertugrul, H.; Wagels, R.; Werner, N.; Hartmann, M.; Feige, K.; Kreienbrock, L. Antimicrobial Usage in Horses: The Use of Electronic Data, Data Curation, and First Results. Front. Vet. Sci. 2020, 7, 216. [Google Scholar] [CrossRef] [PubMed]
- Sultan, I.; Rahman, S.; Jan, A.T.; Siddiqui, M.T.; Mondal, A.H.; Haq, Q.M.R. Antibiotics, Resistome and Resistance Mechanisms: A Bacterial Perspective. Front. Microbiol. 2018, 9, 2066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamame, A.; Davoust, B.; Cherak, Z.; Rolain, J.-M.; Diene, S.M. Mobile Colistin Resistance (Mcr) Genes in Cats and Dogs and Their Zoonotic Transmission Risks. Pathogens 2022, 11, 698. [Google Scholar] [CrossRef] [PubMed]
- Tyrrell, C.; Burgess, C.M.; Brennan, F.P.; Walsh, F. Antibiotic Resistance in Grass and Soil. Biochem. Soc. Trans. 2019, 47, 477–486. [Google Scholar] [CrossRef]
- Cherak, Z.; Loucif, L.; Moussi, A.; Rolain, J.M. Epidemiology of Mobile Colistin Resistance (Mcr) Genes in Aquatic Environments. J. Glob. Antimicrob. Resist. 2021, 27, 51–62. [Google Scholar] [CrossRef]
- Fletcher, S. Understanding the Contribution of Environmental Factors in the Spread of Antimicrobial Resistance. Environ. Health Prev. Med. 2015, 20, 243–252. [Google Scholar] [CrossRef] [Green Version]
- He, Y.; Yuan, Q.; Mathieu, J.; Stadler, L.; Senehi, N.; Sun, R.; Alvarez, P.J.J. Antibiotic Resistance Genes from Livestock Waste: Occurrence, Dissemination, and Treatment. NPJ Clean Water 2020, 3, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Steinman, A.; Navon-Venezia, S. Antimicrobial Resistance in Horses. Animals 2020, 10, 1161. [Google Scholar] [CrossRef]
- Sykes, B.W.; Furr, M.O. Equine Endotoxaemia-A State-of-the-Art Review of Therapy. Aust. Vet. J. 2005, 83, 45–52. [Google Scholar] [CrossRef]
- Dierikx, C.M.; Meijs, A.P.; Hengeveld, P.D.; van der Klis, F.R.M.; van Vliet, J.; Gijsbers, E.F.; Rozwandowicz, M.; van Hoek, A.H.A.M.; Hendrickx, A.P.A.; Hordijk, J.; et al. Colistin-Resistant Enterobacterales among Veterinary Healthcare Workers and in the Dutch Population. JAC-Antimicrob. Resist. 2022, 4, dlac041. [Google Scholar] [CrossRef] [PubMed]
- Moreira Soares de Souza, M.; Marcos Rocha-de-Souza, C.; Araújo de Melo, D.; Couto da Motta, C.; Loureiro Pimenta, R.; da Silva Coelho, I.; de Mattos de Oliveira Coelho, S. Of Animal and Men: The Importance of Animal Environment to Antimicrobial Resistance: A One Health Approach. In Antimicrobial Resistance-A One Health Perspective; Books on Demand: Norderstedt, Germany, 2021. [Google Scholar]
- Apostolakos, I.; Franz, E.; van Hoek, A.H.A.M.; Florijn, A.; Veenman, C.; Sloet-van Oldruitenborgh-Oosterbaan, M.M.; Dierikx, C.; van Duijkeren, E. Occurrence and Molecular Characteristics of ESBL/AmpC-Producing Escherichia coli in Faecal Samples from Horses in an Equine Clinic. J. Antimicrob. Chemother. 2017, 72, 1915–1921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandes, M.R.; Cerdeira, L.; Silva, M.M.; Sellera, F.P.; Muñoz, M.; Junior, F.G.; Azevedo, S.S.; Power, P.; Gutkind, G.; Lincopan, N. Novel Mcr-5.3 Variant in a CTX-M-8-Producing Escherichia coli ST711 Isolated from an Infected Horse. J. Antimicrob. Chemother. 2018, 73, 3520–3522. [Google Scholar] [CrossRef] [PubMed]
- Börjesson, S.; Greko, C.; Myrenås, M.; Landén, A.; Nilsson, O.; Pedersen, K. A Link between the Newly Described Colistin Resistance Gene Mcr-9 and Clinical Enterobacteriaceae Isolates Carrying BlaSHV-12 from Horses in Sweden. J. Glob. Antimicrob. Resist. 2020, 20, 285–289. [Google Scholar] [CrossRef]
- Touati, M.; Hadjadj, L.; Berrazeg, M.; Baron, S.A.; Rolain, J.M. Emergence of Escherichia coli Harbouring Mcr-1 and Mcr-3 Genes in North West Algerian Farmlands. J. Glob. Antimicrob. Resist. 2020, 21, 132–137. [Google Scholar] [CrossRef]
- Avraam, C.; Lambrou, A.S.; Jiang, W.; Siddiqui, S. Antimicrobial Resistance and Livestock Trade for Low and Middle Income Countries: Regional Analysis of Global Coordination Policies. Front. Sustain. Food Syst. 2021, 5, 650315. [Google Scholar] [CrossRef]
- Furuhata, K.; Ishizaki, N.; Sugi, Y.; Fukuyama, M. Isolation and Identification of Enterobacteriaceae from Raw Horsemeat Intended for Human Consumption (Basashi). Biocontrol Sci. 2014, 19, 181–188. [Google Scholar] [CrossRef]
- Stanciu, S. Horse Meat Consumption—Between Scandal and Reality. Procedia Econ. Financ. 2015, 23, 697–703. [Google Scholar] [CrossRef] [Green Version]
- Wolny-Koładka, K.; Lenart-Boroń, A. Antimicrobial Resistance and the Presence of Extended-Spectrum Beta-Lactamase Genes in Escherichia coli Isolated from the Environment of Horse Riding Centers. Environ. Sci. Pollut. Res. 2018, 25, 21789–21800, Correction on: Environ. Sci. Pollut. Res. 2018, 25, 31790–31791. [Google Scholar] [CrossRef]
- Hassan, J.; Kassem, I.I. Audacious Hitchhikers: The Role of Travel and the International Food Trade in the Global Dissemination of Mobile Colistin-Resistance (Mcr) Genes. Antibiotics 2020, 9, 370. [Google Scholar] [CrossRef]
- Manges, A.R.; Geum, H.M.; Guo, A.; Edens, T.J.; Fibke, C.D.; Pitout, J.D.D. Global Extraintestinal Pathogenic Escherichia coli (Expec) Lineages. Clin. Microbiol. Rev. 2019, 32, e00135-18. [Google Scholar] [CrossRef] [PubMed]
- Bachiri, T.; Lalaoui, R.; Bakour, S.; Allouache, M.; Belkebla, N.; Rolain, J.M.; Touati, A. First Report of the Plasmid-Mediated Colistin Resistance Gene mcr-1 in Escherichia coli ST405 Isolated from Wildlife in Bejaia, Algeria. Microb. Drug Resist. 2018, 24, 890–895. [Google Scholar] [CrossRef] [PubMed]
- Berrazeg, M.; Hadjadj, L.; Ayad, A.; Drissi, M.; Rolain, J.M. First Detected Human Case in Algeria of mcr-1 Plasmid-Mediated Colistin Resistance in a 2011 Escherichia coli Isolate. Antimicrob. Agents Chemother. 2016, 60, 6996–6997. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yanat, B.; Machuca, J.; Yahia, R.D.; Touati, A.; Pascual, Á.; Rodríguez-Martínez, J.M. First Report of the Plasmid-Mediated Colistin Resistance Gene Mcr-1 in a Clinical Escherichia coli Isolate in Algeria. Int. J. Antimicrob. Agents 2016, 48, 760–761. [Google Scholar] [CrossRef]
- Chaalal, N.; Touati, A.; Yahiaoui-Martinez, A.; Aissa, M.A.; Sotto, A.; Lavigne, J.P.; Pantel, A. Colistin-Resistant Enterobacterales Isolated from Chicken Meat in Western Algeria. Microb. Drug Resist. 2021, 27, 991–1002. [Google Scholar] [CrossRef]
- Li, Y.; Dai, X.; Zeng, J.; Gao, Y.; Zhang, Z.; Zhang, L. Characterization of the Global Distribution and Diversified Plasmid Reservoirs of the Colistin Resistance Gene Mcr-9. Sci. Rep. 2020, 10, 8113. [Google Scholar] [CrossRef]
- Ha, V.T.T.; Tran, L.D.; Mai, N.T.T.; Hirabayashi, A.; Nguyen, S.T.; Tran, H.H.; Shibayama, K.; Suzuki, M. Potential Spread of mcr-9-Carrying IncHI2 Plasmids in Enterobacter hormaechei in Vietnam. J. Glob. Antimicrob. Resist. 2021, 27, 332–334. [Google Scholar] [CrossRef]
- Liao, W.; Cui, Y.; Quan, J.; Zhao, D.; Han, X.; Shi, Q.; Wang, Q.; Jiang, Y.; Du, X.; Li, X.; et al. High Prevalence of Colistin Resistance and Mcr-9/10 Genes in Enterobacter spp. in a Tertiary Hospital over a Decade. Int. J. Antimicrob. Agents 2022, 59, 106573. [Google Scholar] [CrossRef]
- Sadek, M.; Nariya, H.; Shimamoto, T.; Kayama, S.; Yu, L.; Hisatsune, J.; Sugai, M.; Nordmann, P.; Poirel, L.; Shimamoto, T. First Genomic Characterization of blaVIM-1 and mcr-9-Coharbouring Enterobacter hormaechei Isolated from Food of Animal Origin. Pathogens 2020, 9, 687. [Google Scholar] [CrossRef]
- Fuentes-Castillo, D.; Sellera, F.P.; Goldberg, D.W.; Fontana, H.; Esposito, F.; Cardoso, B.; Ikeda, J.; Kyllar, A.; Catão-Dias, J.L.; Lincopan, N. Colistin-Resistant Enterobacter kobei Carrying mcr-9.1 and blaCTX-M-15 infecting a critically endangered franciscana dolphin (Pontoporia blainvillei), Brazil. Transbound. Emerg. Dis. 2021, 68, 3048–3054. [Google Scholar] [CrossRef]
- Umeda, K.; Nakamura, H.; Fukuda, A.; Matsumoto, Y.; Motooka, D.; Nakamura, S.; Yasui, Y.; Yoshida, H.; Kawahara, R. Genomic Characterization of Clinical Enterobacter roggenkampii Co-Harbouring BlaIMP-1- and BlaGES-5-Encoding IncP6 and Mcr-9-Encoding IncHI2 Plasmids Isolated in Japan. J. Glob. Antimicrob. Resist. 2020, 24, 220–227. [Google Scholar] [CrossRef] [PubMed]
- Gogry, F.A.; Siddiqui, M.T.; Sultan, I.; Haq, Q.M.R. Current Update on Intrinsic and Acquired Colistin Resistance Mechanisms in Bacteria. Front. Med. 2021, 8, 677720. [Google Scholar] [CrossRef] [PubMed]
- Kieffer, N.; Royer, G.; Decousser, J.W.; Bourrel, A.S.; Palmieri, M.; de La Rosa, J.M.O.; Jacquier, H.; Denamur, E.; Nordmann, P.; Poirel, L. Mcr-9, an Inducible Gene Encoding an Acquired Phosphoethanolamine Transferase in Escherichia coli, and Its Origin. Antimicrob. Agents Chemother. 2019, 63, e00965-19, Erratum on: Antimicrob. Agents Chemother. 2019, 63, e01866-19. [Google Scholar] [CrossRef] [Green Version]
- Elbediwi, M.; Pan, H.; Zhou, X.; Rankin, S.C.; Schifferli, D.M.; Yue, M. Detection of Mcr-9-Harbouring ESBL-Producing Salmonella Newport Isolated from an Outbreak in a Large-Animal Teaching Hospital in the USA. J. Antimicrob. Chemother. 2021, 76, 1107–1109. [Google Scholar] [CrossRef]
- Doumith, M.; Godbole, G.; Ashton, P.; Larkin, L.; Dallman, T.; Day, M.; Day, M.; Muller-Pebody, B.; Ellington, M.J.; de Pinna, E.; et al. Detection of the Plasmid-Mediated Mcr-1 Gene Conferring Colistin Resistance in Human and Food Isolates of Salmonella enterica and Escherichia coli in England and Wales. J. Antimicrob. Chemother. 2016, 71, 2300–2305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eichhorn, I.; Feudi, C.; Wang, Y.; Kaspar, H.; Feßler, A.T.; Lübke-Becker, A.; Michael, G.B.; Shen, J.; Schwarz, S. Identification of Novel Variants of the Colistin Resistance Gene Mcr-3 in Aeromonas spp. from the National Resistance Monitoring Programme GERM-Vet and from Diagnostic Submissions. J. Antimicrob. Chemother. 2018, 73, 1217–1221. [Google Scholar] [CrossRef]
- Lupo, A.; Saras, E.; Madec, J.Y.; Haenni, M. Emergence of Bla CTX-M-55 Associated with FosA, RmtB and Mcr Gene Variants in Escherichia coli from Various Animal Species in France. J. Antimicrob. Chemother. 2018, 73, 867–872. [Google Scholar] [CrossRef]
- Barbieri, N.L.; Pimenta, R.L.; de Melo, D.A.; Nolan, L.K.; de Souza, M.M.S.; Logue, C.M. Mcr-1 Identified in Fecal Escherichia coli and Avian pathogenic e. coli (APEC) From Brazil. Front. Microbiol. 2021, 12, 799. [Google Scholar] [CrossRef]
- Rabello, R.F.; Bonelli, R.R.; Penna, B.A.; Albuquerque, J.P.; Souza, R.M.; Cerqueira, A.M.F. Antimicrobial Resistance in Farm Animals in Brazil: An Update Overview. Animals 2020, 10, 552. [Google Scholar] [CrossRef] [Green Version]
- Tyso, G.H.; Li, C.; Hsu, C.H.; Ayers, S.; Borenstein, S.; Mukherjee, S.; Tran, T.T.; McDermot, P.F.; Zhao, S. The Mcr-9 Gene of Salmonella and Escherichia coli Is Not Associated with Colistin Resistance in the United States. Antimicrob. Agents Chemother. 2020, 64, e00573-20. [Google Scholar] [CrossRef]
- Dallap Schaer, B.L.; Aceto, H.; Rankin, S.C. Outbreak of Salmonellosis Caused by Salmonella Enterica Serovar Newport MDR-AmpC in a Large Animal Veterinary Teaching Hospital. J. Vet. Intern. Med. 2010, 24, 1138–1146. [Google Scholar] [CrossRef] [PubMed]
- Marco-Fuertes, A.; Marin, C.; Lorenzo-Rebenaque, L.; Vega, S.; Montoro-Dasi, L. Antimicrobial Resistance in Companion Animals: A New Challenge for the One Health Approach in the European Union. Vet. Sci. 2022, 9, 208. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Lu, C.; Shen, D.; Liu, J.; Ma, Z.; Yang, B.; Ling, W.; Waigi, M.G. Elimination of the Risks of Colistin Resistance Gene (Mcr-1) in Livestock Manure during Composting. Environ. Int. 2019, 126, 61–68. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Country | Source of Isolates | Date of Isolation (mcr Gene Assayed) | Number of Isolates Tested for mcr | Identified Gene/Variant (Number of Organisms) | Sequence Type and/or Phylogroup (Virulence Genes) | Plasmid (Associated Insertion Sequence) | Additional Resistance Traits | Reference |
|---|---|---|---|---|---|---|---|---|
| Algeria | Horse manure | 2016–2018 (mcr-1-mcr-5) | - | mcr-1 (1 E. coli) | - | - | blaTEM-12 | [57] |
| Sweden | Uterus, wound, trachea, skin biopsy, urine, and thrombophlebitis | 2015–2018 (mcr-1 to mcr-10) | 56 | Mcr-9 (15 Enterobacter cloacae complex, 10 E. coli, 4 Klebsiella oxytoca, and 1 Citrobacter freundii) | E. coli: ST1861, ST2557, ST9329, ST1252, ST1423, and ST4398; | IncFIB, IncFII, IncFIC, IncFI, IncP, IncA/C, and Col (MG828) | blaSHV-12, blaTEM-1B, blaACT, blaCMY-82, blaOXY, blaSCO-1, aac(6′)-Ib, aadA2, ant(3″)-Ia, aph(3′)-Ia, aph(3″)-Ib, aadA1, catA2, dfrA19, qnrB2, qnrA1, qnrB10, tet(D), fosA, mdf(A), ere(A), sul1, and sul2 | [56] |
| Enterobacter clocae complex: ST113, ST116, ST1021, ST51, ST102, ST88, and ST254; | ||||||||
| Klebsiella oxytoca: ST238, ST37, and ST2; | ||||||||
| Citrobacter freundii: ST233 | ||||||||
| USA | Horse veterinary clinic environment | 2003–2004 (mcr-1 to mcr-10) | 31 | mcr-9 (1 Salmonella Newport) | ST45 (SPI-I and II, fim and other fimbrial colonization factors) | IncHI2 (IS26 and IS903B) | blaCMY-2, blaSHV-12, blaTEM-1B, aph(4)-Ia, aac(6’)-Iaa, aadA2, aac(3)-IV, aph(6)-Id, aph(3″)-Ia, aph(3″)-Ib, mdf(A), catA2, floR, fexA, sul1, sul2, dfrA19, tet(A), tet(D), qnrB2, and mutation in parC | [76] |
| Brazil | Lungs of dead horse | 2012 (mcr-1 to mcr-10) | 1 | mcr-5.3 (1 E. coli) | ST711 (gad, lpfA, and iss) | - | blaCTX-M-8, blaTEM-1B, aac(3)-IId, aadA2, strA, strB, sul2, and dfrA12 | [55] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Anyanwu, M.U.; Jaja, I.F.; Nwobi, O.C.; Mgbeahuruike, A.C.; Ikpendu, C.N.; Okafor, N.A.; Oguttu, J.W. Epidemiology and Traits of Mobile Colistin Resistance (mcr) Gene-Bearing Organisms from Horses. Microorganisms 2022, 10, 1499. https://doi.org/10.3390/microorganisms10081499
Anyanwu MU, Jaja IF, Nwobi OC, Mgbeahuruike AC, Ikpendu CN, Okafor NA, Oguttu JW. Epidemiology and Traits of Mobile Colistin Resistance (mcr) Gene-Bearing Organisms from Horses. Microorganisms. 2022; 10(8):1499. https://doi.org/10.3390/microorganisms10081499
Chicago/Turabian StyleAnyanwu, Madubuike Umunna, Ishmael Festus Jaja, Obichukwu Chisom Nwobi, Anthony Christian Mgbeahuruike, Chinaza Nnenna Ikpendu, Nnenna Audrey Okafor, and James Wabwire Oguttu. 2022. "Epidemiology and Traits of Mobile Colistin Resistance (mcr) Gene-Bearing Organisms from Horses" Microorganisms 10, no. 8: 1499. https://doi.org/10.3390/microorganisms10081499