
A Chikungunya Virus Multiepitope Recombinant Protein Expressed from the Binary System Insect Cell/Recombinant Baculovirus Is Useful for Laboratorial Diagnosis of Chikungunya


 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Construction of Recombinant Baculovirus Containing R1 Gene
2.2. Expression and Recombinant OBs Purification
2.3. Scanning Electron Microscopy and Recombinant OBs Synthesis Analysis
2.4. Ethical Statement and Human Serum Samples
2.5. Multiepitope Recombinant Protein-Based IgG-ELISA for the Detection of Anti-CHIKV Specific IgG Antibodies (MULTREC IgG-ELISA)
3. Results
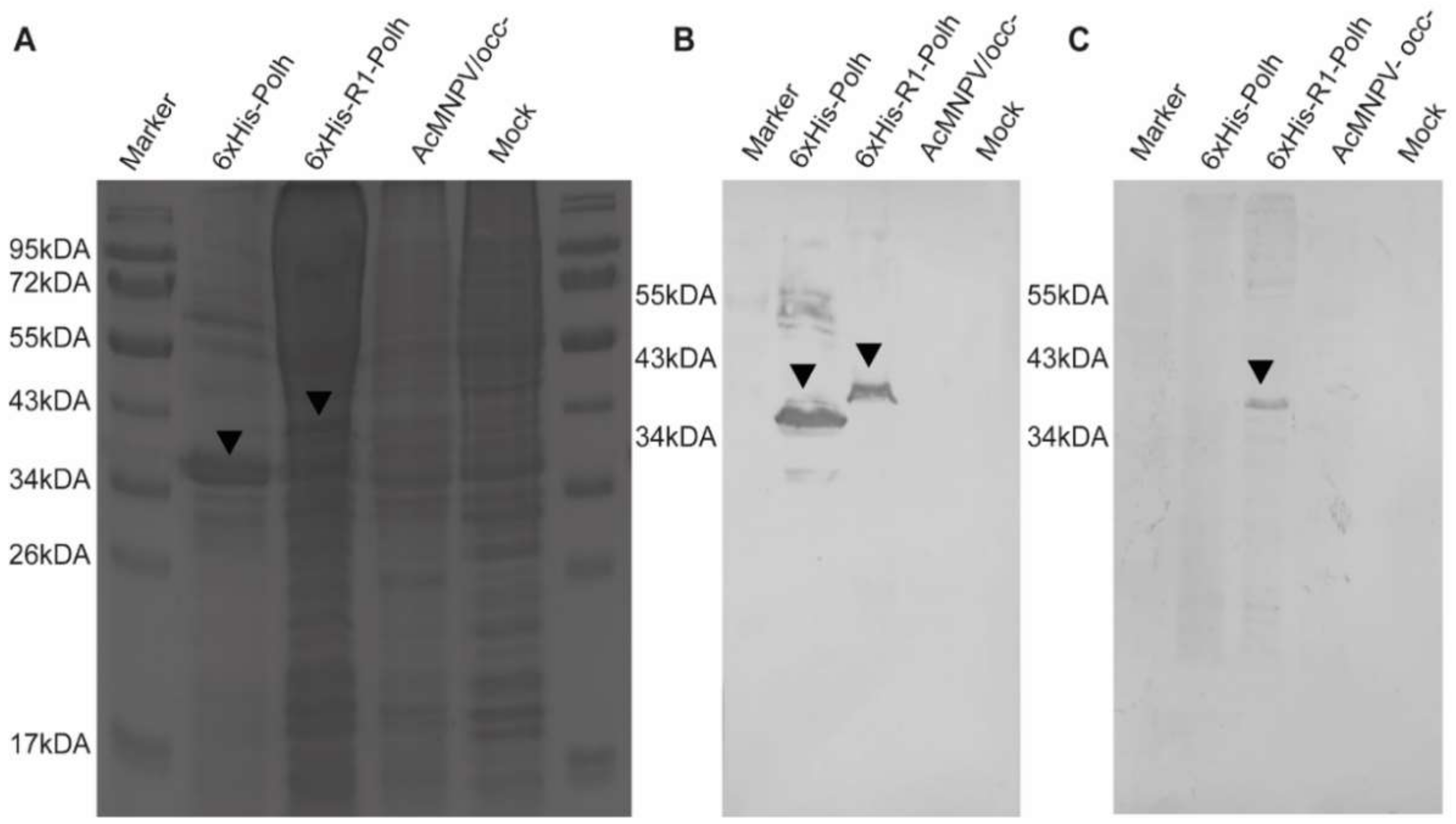
3.1. Fusion of the CHIKV Multi-Epitope Gene-Coding Sequence at Amino-Terminal Region of the Modified Polh
3.2. Ultrastructural Analysis of the CHIKV Antigen-Containing Recombinant Crystal Revealed Smaller Crystals Than the Parental Version
3.3. CHIKV Antigen-Containing Recombinant Crystal Immunoblot
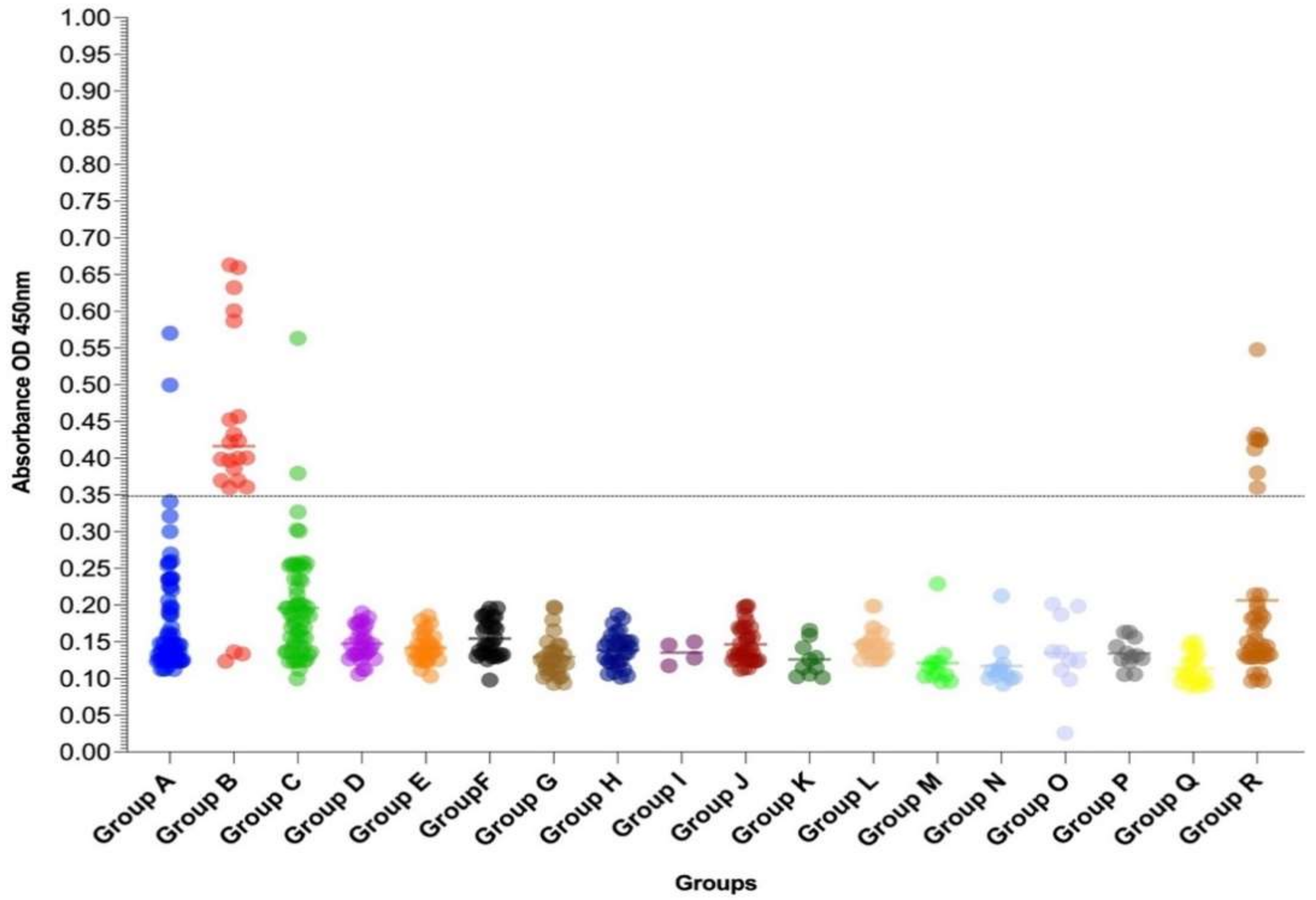
3.4. Multiepitope Recombinant Protein-Based IgG-ELISA for the Detection of Anti-CHIKV Specific IgG Antibodies (MULTREC IgG-ELISA)
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chikungunya Cases Identified Through Passive Surveillance and Household Investigations—Puerto Rico, May 5–August 12, 2014. Available online: https://www.cdc.gov/mmwr/preview/mmwrhtml/mm6348a1.htm (accessed on 2 December 2021).
- Vairo, F.; Haider, N.; Kock, R.; Ntoumi, F.; Ippolito, G.; Zumla, A. Chikungunya: Epidemiology, Pathogenesis, Clinical Features, Management, and Prevention. Infect. Dis. Clin. N. Am. 2019, 33, 1003–1025. [Google Scholar] [CrossRef] [PubMed]
- Puntasecca, C.J.; King, C.H.; Labeaud, A.D. Measuring the global burden of chikungunya and Zika viruses: A systematic review. PLoS Negl. Trop. Dis. 2021, 15, e0009055. [Google Scholar] [CrossRef] [PubMed]
- Zeller, H.; Van Bortel, W.; Sudre, B. Chikungunya: Its History in Africa and Asia and Its Spread to New Regions in 2013–2014. J. Infect. Dis. 2016, 214, S436–S440. [Google Scholar] [CrossRef] [PubMed]
- Powers, A.M. Genomic evolution and phenotypic distinctions of Chikungunya viruses causing the Indian Ocean outbreak. Exp. Biol. Med. 2011, 236, 909–914. [Google Scholar] [CrossRef]
- Neuvonen, M.; Kazlauskas, A.; Martikainen, M.; Hinkkanen, A.; Ahola, T.; Saksela, K. SH3 domain-mediated recruitment of host cell amphiphysins by alphavirus nsP3 promotes viral RNA replication. PLoS Pathog. 2011, 7, e1002383. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.F.; Sawicki, S.G.; Sawicki, D.L. Alphavirus nsP3 functions to form replication complexes transcribing negative-strand RNA. J. Virol. 1994, 68, 6466–6475. [Google Scholar] [CrossRef] [Green Version]
- Teppor, M.; Žusinaite, E.; Merits, A. Phosphorylation Sites in the Hypervariable Domain in Chikungunya Virus nsP3 Are Crucial for Viral Replication. J. Virol. 2021, 95, e02276-20. [Google Scholar] [CrossRef]
- Götte, B.; Liu, L.; McInerney, G.M. The Enigmatic Alphavirus Non-Structural Protein 3 (nsP3) Revealing Its Secrets at Last. Viruses 2018, 10, 105. [Google Scholar] [CrossRef] [Green Version]
- Nandi, I.; Gupta, A.; Chaudhary, V.K.; Gupta, V.; Gabrani, R.; Gupta, S. Expression, purification and functional characterization of recombinant hypervariable region (HVR) of Chikungunya virus nsP3 protein. 3 Biotech 2019, 9, 235. [Google Scholar] [CrossRef]
- Nowee, G.; Bakker, J.W.; Geertsema, C.; Ros, V.I.D.; Göertz, G.P.; Fros, J.J.; Pijlman, G.P. A Tale of 20 Alphaviruses; Inter-species Diversity and Conserved Interactions Between Viral Non-structural Protein 3 and Stress Granule Proteins. Front. Cell Dev. Biol. 2021, 9, 625711. [Google Scholar] [CrossRef]
- Petitdemange, C.; Wauquier, N.; Vieillard, V. Control of immunopathology during chikungunya virus infection. J. Allergy Clin. Immunol. 2015, 135, 846–855. [Google Scholar] [CrossRef]
- Reddy, A.; Bosch, I.; Salcedo, N.; Herrera, B.B.; de Puig, H.; Narváez, C.F.; Caicedo-Borrero, D.M.; Lorenzana, I.; Parham, L.; García, K.; et al. Development and Validation of a Rapid Lateral Flow E1/E2-Antigen Test and ELISA in Patients Infected with Emerging Asian Strain of Chikungunya Virus in the Americas. Viruses 2020, 12, 971. [Google Scholar] [CrossRef]
- Thiberville, S.D.; Boisson, V.; Gaudart, J.; Simon, F.; Flahault, A.; de Lamballerie, X. Chikungunya fever: A clinical and virological investigation of outpatients on Reunion Island, South-West Indian Ocean. PLoS Negl. Trop. Dis. 2013, 7, e2004. [Google Scholar] [CrossRef]
- Rezza, G.; Weaver, S.C. Chikungunya as a paradigm for emerging viral diseases: Evaluating disease impact and hurdles to vaccine development. PLoS Negl. Trop. Dis. 2019, 13, e0006919. [Google Scholar] [CrossRef]
- Mehta, R.; Gerardin, P.; de Brito, C.A.A.; Soares, C.N.; Ferreira, M.L.B.; Solomon, T. The neurological complications of chikungunya virus: A systematic review. Rev. Med. Virol. 2018, 28, e1978. [Google Scholar] [CrossRef] [Green Version]
- Da Mello, C.S.; Cabral-Castro, M.J.; de Faria Silva, L.C.; Peralta, J.M.; Puccioni-Sohler, M. Dengue and chikungunya infection in neurologic disorders from endemic areas in Brazil. Neurol. Clin. Pract. 2020, 10, 497–502. [Google Scholar] [CrossRef]
- Pierro, A.; Rossini, G.; Gaibani, P.; Finarelli, A.C.; Moro, M.L.; Landini, M.P.; Sambri, V. Persistence of anti-chikungunya virus-specific antibodies in a cohort of patients followed from the acute phase of infection after the 2007 outbreak in Italy. N. Microbes N. Infect. 2015, 7, 23–25. [Google Scholar] [CrossRef] [Green Version]
- Azami, N.A.M.; Salleh, S.A.; Shah, S.A.; Neoh, H.M.; Othman, Z.; Zakaria, S.Z.S.; Jamal, R. Emergence of chikungunya seropositivity in healthy Malaysian adults residing in outbreak-free locations: Chikungunya seroprevalence results from the Malaysian Cohort. BMC Infect. Dis. 2013, 13, 67. [Google Scholar] [CrossRef] [Green Version]
- Nunes, M.R.T.; Silva, S.P.; Carvalho, V.L.; Vasconcelos, J.M.; Da Silva, D.E.A.; Oliveira, L.F.; Nunes Neto, J.P.; Rodrigues, S.G.; Azevedo, R.S.S.; Monteiro, H.A.O.; et al. Emergence of New Insect-Restrictive Viruses in the Amazon Region. Genome Announc. 2015, 3, e00131-15. [Google Scholar] [CrossRef] [Green Version]
- Da Costa, A.C.; Thézé, J.; Komninakis, S.C.V.; Sanz-Duro, R.L.; Felinto, M.R.L.; Moura, L.C.C.; De Oliveira Barroso, I.M.; Santos, L.E.C.; De Lemos Nunes, M.A.; Moura, A.A.; et al. Spread of Chikungunya Virus East/Central/South African Genotype in Northeast Brazil. Emerg. Infect. Dis. 2017, 23, 1742–1744. [Google Scholar] [CrossRef] [Green Version]
- Naveca, F.G.; Claro, I.; Giovanetti, M.; de Jesus, J.G.; Xavier, J.; Iani, F.C.D.M.; do Nascimento, V.A.; de Souza, V.C.; Silveira, P.P.; Lourenço, J.; et al. Genomic, epidemiological and digital surveillance of Chikungunya virus in the Brazilian Amazon. PLoS Negl. Trop. Dis. 2019, 13, e0007065. [Google Scholar] [CrossRef] [Green Version]
- Xavier, J.; Fonseca, V.; Bezerra, J.F.; do Monte Alves, M.; Mares-Guia, M.A.; Claro, I.M.; de Jesus, R.; Adelino, T.; Araújo, E.; Cavalcante, K.R.L.J.; et al. Chikungunya virus ECSA lineage reintroduction in the northeasternmost region of Brazil. Int. J. Infect. Dis. 2021, 105, 120–123. [Google Scholar] [CrossRef]
- Pereira Gusmão Maia, Z.; Mota Pereira, F.; do Carmo Said, R.F.; Fonseca, V.; Gräf, T.; de Bruycker Nogueira, F.; Brandão Nardy, V.; Xavier, J.; Lima Maia, M.; Abreu, A.L.; et al. Return of the founder Chikungunya virus to its place of introduction into Brazil is revealed by genomic characterization of exanthematic disease cases. Emerg. Microbes Infect. 2019, 9, 53–57. [Google Scholar] [CrossRef] [Green Version]
- Pacheco, O.E.; Martínez, M.; Alarcón, Á.; Bonilla, M.; Caycedo, A.; Valbuena, T.; Zabaleta, A. Estimation of underreporting of Chikungunya virus infection cases in Girardot, Colombia, from November, 2014, to May, 2015. Biomedica 2017, 37, 507–515. [Google Scholar] [CrossRef] [Green Version]
- Oidtman, R.J.; España, G.; Alex Perkins, T. Co-circulation and misdiagnosis led to underestimation of the 2015-2017 Zika epidemic in the Americas. PLoS Negl. Trop. Dis. 2021, 15, e0009208. [Google Scholar] [CrossRef]
- De Zanotto, P.M.A.; de Leite, L.C.C. The Challenges Imposed by Dengue, Zika, and Chikungunya to Brazil. Front. Immunol. 2018, 9, 1964. [Google Scholar] [CrossRef] [PubMed]
- Kumar, N.P.; Suresh, A.; Vanamail, P.; Sabesan, S.; Krishnamoorthy, K.G.; Mathew, J.; Jose, V.T.; Jambulingam, P. Chikungunya virus outbreak in Kerala, India, 2007: A seroprevalence study. Mem. Inst. Oswaldo Cruz 2011, 106, 912–916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fritzell, C.; Rousset, D.; Adde, A.; Kazanji, M.; Van Kerkhove, M.D.; Flamand, C. Current challenges and implications for dengue, chikungunya and Zika seroprevalence studies worldwide: A scoping review. PLoS Negl. Trop. Dis. 2018, 12, e0006533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Périssé, A.R.S.; Souza-Santos, R.; Duarte, R.; Santos, F.; De Andrade, C.R.; Rodrigues, N.C.P.; De Andrade Schramm, J.M.; Da Silva, E.D.; Da Silva Viana Jacobson, L.; Lemos, M.C.F.; et al. Zika, dengue and chikungunya population prevalence in Rio de Janeiro city, Brazil, and the importance of seroprevalence studies to estimate the real number of infected individuals. PLoS ONE 2020, 15, e0243239. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, N.K.; Shrivastva, A.; Pattnaik, P.; Parida, M.; Dash, P.K.; Jana, A.M.; Rao, P.V.L. Production, purification and characterization of recombinant dengue multiepitope protein. Biotechnol. Appl. Biochem. 2007, 46, 105. [Google Scholar] [CrossRef]
- Maldaner, F.R.; Aragão, F.J.L.; Dos Santos, F.B.; Franco, O.L.; Da Rocha Queiroz Lima, M.; De Oliveira Resende, R.; Vasques, R.M.; Nagata, T. Dengue virus tetra-epitope peptide expressed in lettuce chloroplasts for potential use in dengue diagnosis. Appl. Microbiol. Biotechnol. 2013, 97, 5721–5729. [Google Scholar] [CrossRef] [Green Version]
- Lopes-Luz, L.; Junqueira, I.C.; da Silveira, L.A.; de Melo Pereira, B.R.; da Silva, L.A.; Ribeiro, B.M.; Nagata, T. Dengue and Zika virus multi-epitope antigen expression in insect cells. Mol. Biol. Rep. 2020, 47, 7333–7340. [Google Scholar] [CrossRef]
- Metz, S.W.; Pijlman, G.P. Arbovirus vaccines; opportunities for the baculovirus-insect cell expression system. J. Invertebr. Pathol. 2011, 107, S16–S30. [Google Scholar] [CrossRef]
- Chua, C.L.; Sam, I.C.; Chan, Y.F. Expression and Purification of E2 Glycoprotein from Insect Cells (Sf9) for Use in Serology. Methods Mol. Biol. 2016, 1426, 51–61. [Google Scholar] [CrossRef]
- Ardisson-Araújo, D.M.P.; Rocha, J.R.; Da Costa, M.H.O.; Bocca, A.L.; Dusi, A.N.; De Oliveira Resende, R.; Ribeiro, B.M. A baculovirus-mediated strategy for full-length plant virus coat protein expression and purification. Virol. J. 2013, 10, 262. [Google Scholar] [CrossRef] [Green Version]
- Silva, L.A.; Camargo, B.R.; Araújo, A.C.; Batista, T.L.; Ribeiro, B.M.; Ardisson-Araújo, D.M.P. Easily purified baculovirus/insect-system-expressed recombinant hepatitis B virus surface antigen fused to the N- or C-terminus of polyhedrin. Arch. Virol. 2021, 167, 345–354. [Google Scholar] [CrossRef]
- O’Reilly, D.R.; Miller, L.K.; Luckow, V.A. Baculovirus Expression Vectors. A Laboratory Manual; Freeman and Company: New York, NY, USA, 1992; ISBN 0195091310. [Google Scholar]
- Lanciotti, R.S.; Kosoy, O.L.; Laven, J.J.; Panella, A.J.; Velez, J.O.; Lambert, A.J.; Campbell, G.L. Chikungunya virus in US travelers returning from India, 2006. Emerg. Infect. Dis. 2007, 13, 764–767. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention (U.S.); National Center for Emerging and Zoonotic Infectious Diseases (U.S.); Pan American Health Organization. Preparedness and Response for Chikungunya Virus: Introduction in the Americas; Centers for Disease Control and Prevention (U.S.); National Center for Emerging and Zoonotic Infectious Diseases (U.S.); Pan American Health Organization: Atlanta, GA, USA, 2011; p. 150. [Google Scholar]
- Miagostovich, M.P.; Nogueira, R.M.R.; Dos Santos, F.B.; Schatzmayr, H.G.; Araújo, E.S.M.; Vorndam, V. Evaluation of an IgG enzyme-linked immunosorbent assay for dengue diagnosis. J. Clin. Virol. 1999, 14, 183–189. [Google Scholar] [CrossRef]
- Lanciotti, R.S.; Calisher, C.H.; Gubler, D.J.; Chang, G.J.; Vorndam, A.V. Rapid detection and typing of dengue viruses from clinical samples by using reverse transcriptase-polymerase chain reaction. J. Clin. Microbiol. 1992, 30, 545–551. [Google Scholar] [CrossRef] [Green Version]
- Lanciotti, R.S.; Kosoy, O.L.; Laven, J.J.; Velez, J.O.; Lambert, A.J.; Johnson, A.J.; Stanfield, S.M.; Duffy, M.R. Genetic and serologic properties of Zika virus associated with an epidemic, Yap State, Micronesia, 2007. Emerg. Infect. Dis. 2008, 14, 1232–1239. [Google Scholar] [CrossRef]
- Dos Santos, F.B.; Nogueira, R.M.R.; Lima, M.R.Q.; De Simone, T.S.; Schatzmayr, H.G.; Lemes, E.M.B.; Harris, E.; Miagostovich, M.P. Recombinant polypeptide antigen-based immunoglobulin G enzyme-linked immunosorbent assay for serodiagnosis of dengue. Clin. Vaccine Immunol. 2007, 14, 641–643. [Google Scholar] [CrossRef] [Green Version]
- Pialoux, G.; Gaüzère, B.A.; Jauréguiberry, S.; Strobel, M. Chikungunya, an epidemic arbovirosis. Lancet Infect. Dis. 2007, 7, 319–327. [Google Scholar] [CrossRef]
- Honório, N.A.; Câmara, D.C.P.; Calvet, G.A.; Brasil, P. Chikungunya: An arbovirus infection in the process of establishment and expansion in Brazil. Cad. Saude Publica 2015, 31, 906–908. [Google Scholar] [CrossRef] [Green Version]
- Dash, M.; Mohanty, I.; Padhi, S. Laboratory diagnosis of chikungunya virus: Do we really need it? Indian J. Med. Sci. 2011, 65, 83–91. [Google Scholar] [CrossRef]
- Yap, G.; Pok, K.Y.; Lai, Y.L.; Hapuarachchi, H.C.; Chow, A.; Leo, Y.S.; Tan, L.K.; Ng, L.C. Evaluation of Chikungunya diagnostic assays: Differences in sensitivity of serology assays in two independent outbreaks. PLoS Negl. Trop. Dis. 2010, 4, e753. [Google Scholar] [CrossRef] [PubMed]
- Johnson, B.W.; Goodman, C.H.; Holloway, K.; De Salazar, P.M.; Valadere, A.M.; Drebot, M.A. Evaluation of Commercially Available Chikungunya Virus Immunoglobulin M Detection Assays. Am. J. Trop. Med. Hyg. 2016, 95, 182–192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mendoza, E.J.; Robinson, A.; Dimitrova, K.; Mueller, N.; Holloway, K.; Makowski, K.; Wood, H. Combining anti-IgM and IgG immunoassays for comprehensive chikungunya virus diagnostic testing. Zoonoses Public Health 2019, 66, 909–917. [Google Scholar] [CrossRef] [PubMed]
- Kikuti, M.; Tauro, L.B.; Moreira, P.S.S.; Nascimento, L.C.J.; Portilho, M.M.; Soares, G.C.; Weaver, S.C.; Reis, M.G.; Kitron, U.; Ribeiro, G.S. Evaluation of two commercially available chikungunya virus IgM enzyme-linked immunoassays (ELISA) in a setting of concomitant transmission of chikungunya, dengue and Zika viruses. Int. J. Infect. Dis. 2020, 91, 38–43. [Google Scholar] [CrossRef] [Green Version]
- Da Rocha Queiroz Lima, M.; de Lima, R.C.; de Azeredo, E.L.; Dos Santos, F.B. Analysis of a Routinely Used Commercial Anti-Chikungunya IgM ELISA Reveals Cross-Reactivities with Dengue in Brazil: A New Challenge for Differential Diagnosis? Diagnostics 2021, 11, 819. [Google Scholar] [CrossRef]
- Ohst, C.; Saschenbrecker, S.; Stiba, K.; Steinhagen, K.; Probst, C.; Radzimski, C.; Lattwein, E.; Komorowski, L.; Stöcker, W.; Schlumberger, W. Reliable Serological Testing for the Diagnosis of Emerging Infectious Diseases. Adv. Exp. Med. Biol. 2018, 1062, 19–43. [Google Scholar] [CrossRef]
- Kim, Y.H.; Kim, T.Y.; Park, J.S.; Park, J.S.; Lee, J.; Moon, J.; Chong, C.K.; Junior, I.N.; Ferry, F.R.; Ahn, H.J.; et al. Development and Clinical Evaluation of a Rapid Diagnostic Test for Yellow Fever Non-Structural Protein 1. Korean J. Parasitol. 2019, 57, 283–290. [Google Scholar] [CrossRef]
- Cho, B.; Jeon, B.Y.; Kim, J.; Noh, J.; Kim, J.; Park, M.; Park, S. Expression and evaluation of Chikungunya virus E1 and E2 envelope proteins for serodiagnosis of Chikungunya virus infection. Yonsei Med. J. 2008, 49, 828–835. [Google Scholar] [CrossRef] [Green Version]
- Warter, L.; Lee, C.Y.; Thiagarajan, R.; Grandadam, M.; Lebecque, S.; Lin, R.T.P.; Bertin-Maghit, S.; Ng, L.F.P.; Abastado, J.-P.; Desprès, P.; et al. Chikungunya virus envelope-specific human monoclonal antibodies with broad neutralization potency. J. Immunol. 2011, 186, 3258–3264. [Google Scholar] [CrossRef]
- Weber, C.; Berberich, E.; von Rhein, C.; Henß, L.; Hildt, E.; Schnierle, B.S. Identification of Functional Determinants in the Chikungunya Virus E2 Protein. PLoS Negl. Trop. Dis. 2017, 11, e0005318. [Google Scholar] [CrossRef]
- Khan, M.; Dhanwani, R.; Kumar, J.S.; Rao, P.V.L.; Parida, M. Comparative evaluation of the diagnostic potential of recombinant envelope proteins and native cell culture purified viral antigens of Chikungunya virus. J. Med. Virol. 2014, 86, 1169–1175. [Google Scholar] [CrossRef]
- Kam, Y.-W.; Lee, W.W.L.; Simarmata, D.; Harjanto, S.; Teng, T.-S.; Tolou, H.; Chow, A.; Lin, R.T.P.; Leo, Y.-S.; Rénia, L.; et al. Longitudinal analysis of the human antibody response to Chikungunya virus infection: Implications for serodiagnosis and vaccine development. J. Virol. 2012, 86, 13005–13015. [Google Scholar] [CrossRef] [Green Version]
- Kumar, P.; Pok, K.Y.; Tan, L.K.; Angela, C.; Leo, Y.S.; Ng, L.C. Development and evaluation of baculovirus-expressed Chikungunya virus E1 envelope proteins for serodiagnosis of Chikungunya infection. J. Virol. Methods 2014, 206, 67–75. [Google Scholar] [CrossRef]
- Fumagalli, M.J.; De Souza, W.M.; Espósito, D.L.A.; Silva, A.; Romeiro, M.F.; Martinez, E.Z.; Da Fonseca, B.A.L.; Figueiredo, L.T.M. Enzyme-linked immunosorbent assay using recombinant envelope protein 2 antigen for diagnosis of Chikungunya virus. Virol. J. 2018, 15, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Bagno, F.F.; Godói, L.C.; Figueiredo, M.M.; Rodrigues Sérgio, S.A.; de Fátima Silva Moraes, T.; de Castro Salazar, N.; Kim, Y.C.; Reyes-Sandoval, A.; Guimarães da Fonseca, F. Chikungunya E2 Protein Produced in E. coli and HEK293-T Cells-Comparison of Their Performances in ELISA. Viruses 2020, 12, 939. [Google Scholar] [CrossRef]
- Vasques, R.M.; Lacorte, C.; da Luz, L.L.; Aranda, M.A.; Nagata, T. Development of a new tobamovirus-based viral vector for protein expression in plants. Mol. Biol. Rep. 2019, 46, 97–103. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GROUPS a | Year | REC IgG-ELISA Result/Tested (%) | Kit Euroimmun Anti-Chikungunya Virus IgG Result/Tested (%) | ||
|---|---|---|---|---|---|
| NEGATIVE | POSITIVE | NEGATIVE | POSITIVE | ||
| A (CHIKV IgM positive cases, n = 77) | 2018–2019 | 74/77 (96.10) | 3/77 (3.89) | 77/77 (100) | 0/77 (0) |
| B (CHIKV IgG positive cases, n = 22) | 2018–2019 | 3/22 (13.63) | 19/22 (86.36) | 0/22 (0) | 22/22 (100) |
| C (CHIKV qRT-PCR positive cases, n = 62) | 2018–2019 | 60/62 (96.77) | 2/62 (3.22) | 59/62 (95.16) | 3/62 (4.83) |
| Total for Groups A–C, n = 161 | 137/161 (85.09) | 24/161 (14.90) | 136/161 (84.47) | 25/161 (15.52) | |
| D (DENV-1 cases, n = 30) | 1997–2001 | 30/30 (100) | 0/30 | 30/30 (100) | 0/30 |
| E (DENV-2 cases, n = 30) | 1998–2010 | 30/30 (100) | 0/30 | 30/30 (100) | 0/30 |
| F (DENV-3 cases, n = 30) | 2001–2007 | 30/30 (100) | 0/30 | 30/30 (100) | 0/30 |
| G (DENV-4 cases, n = 30) | 2012–2017 | 30/30 (100) | 0/30 | 30/30 (100) | 0/30 |
| H (DENV IgM positive and IgG negative cases, n = 30) | 1997–2004 | 30/30 (100) | 0/30 | 30/30 (100) | 0/30 |
| I (DENV IgG positive cases, n = 04) | 1999–2001 | 4/4 (100) | 0/4 | 4/4 (100) | 0/4 |
| J (ZIKV qRT-PCR positive cases, n = 35) | 2016–2017 | 35/35 (100) | 0/35 | 35/35 (100) | 0/35 |
| K (YFV cases, n = 10) | 1997–2004 | 10/10 (100) | 0/10 | 10/10 (100) | 0/10 |
| L (YFV vaccinee cases, n = 24) | 1999–2019 | 24/24 (100) | 0/24 | 24/24 (100) | 0/24 |
| M (Rubella cases, n = 12) | 2004 | 12/12 (100) | 0/12 | 12/12 (100) | 0/12 |
| N (Measles cases; n = 12) | 2004 | 12/12 (100) | 0/12 | 12/12 (100) | 0/12 |
| O (Hepatitis C cases, n = 10) | 2013 | 10/10(100) | 0/10 | 10/10 (100) | 0/10 |
| P (Leptospirosis cases, n = 12) | Not available | 12/12 (100) | 0/12 | 12/12 (100) | 0/12 |
| Q (Healthy individuals, n = 22) | Not available | 22/22 (100) | 0/22 | 22/22 (100) | 0/22 |
| R (Arboviruses negative cases, n = 43) | 2000–2004 | 35/43 (81.39) | 08/43 (18.60) | 38/43 (88.37) | 05/43 (11.62) |
| Total for Groups D–R, n = 334 | 326/334 (97.60) | 08/334 (02.39) | 329/334 (98.5) | 05/334 (1.49) | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Silva, L.A.d.; Lima, M.d.R.Q.; de Camargo, B.R.; Guimarães, D.K.d.S.C.; Barbastefano, A.A.L.; Lima, R.C.d.; Damasco, P.V.; Cunha, R.V.d.; de Souza, L.J.; de Azeredo, E.L.; et al. A Chikungunya Virus Multiepitope Recombinant Protein Expressed from the Binary System Insect Cell/Recombinant Baculovirus Is Useful for Laboratorial Diagnosis of Chikungunya. Microorganisms 2022, 10, 1451. https://doi.org/10.3390/microorganisms10071451
Silva LAd, Lima MdRQ, de Camargo BR, Guimarães DKdSC, Barbastefano AAL, Lima RCd, Damasco PV, Cunha RVd, de Souza LJ, de Azeredo EL, et al. A Chikungunya Virus Multiepitope Recombinant Protein Expressed from the Binary System Insect Cell/Recombinant Baculovirus Is Useful for Laboratorial Diagnosis of Chikungunya. Microorganisms. 2022; 10(7):1451. https://doi.org/10.3390/microorganisms10071451
Chicago/Turabian StyleSilva, Leonardo Assis da, Monique da Rocha Queiroz Lima, Brenda Rabello de Camargo, Dyeferson Kened da Silva Coelho Guimarães, Anabele Azevedo Lima Barbastefano, Raquel Curtinhas de Lima, Paulo Vieira Damasco, Rivaldo Venâncio da Cunha, Luiz José de Souza, Elzinandes Leal de Azeredo, and et al. 2022. "A Chikungunya Virus Multiepitope Recombinant Protein Expressed from the Binary System Insect Cell/Recombinant Baculovirus Is Useful for Laboratorial Diagnosis of Chikungunya" Microorganisms 10, no. 7: 1451. https://doi.org/10.3390/microorganisms10071451