
Retrospective Investigation of the Whole Genome of the Hypovirulent Listeria monocytogenes Strain of ST201, CC69, Lineage III, Isolated from a Piglet with Fatal Neurolisteriosis


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
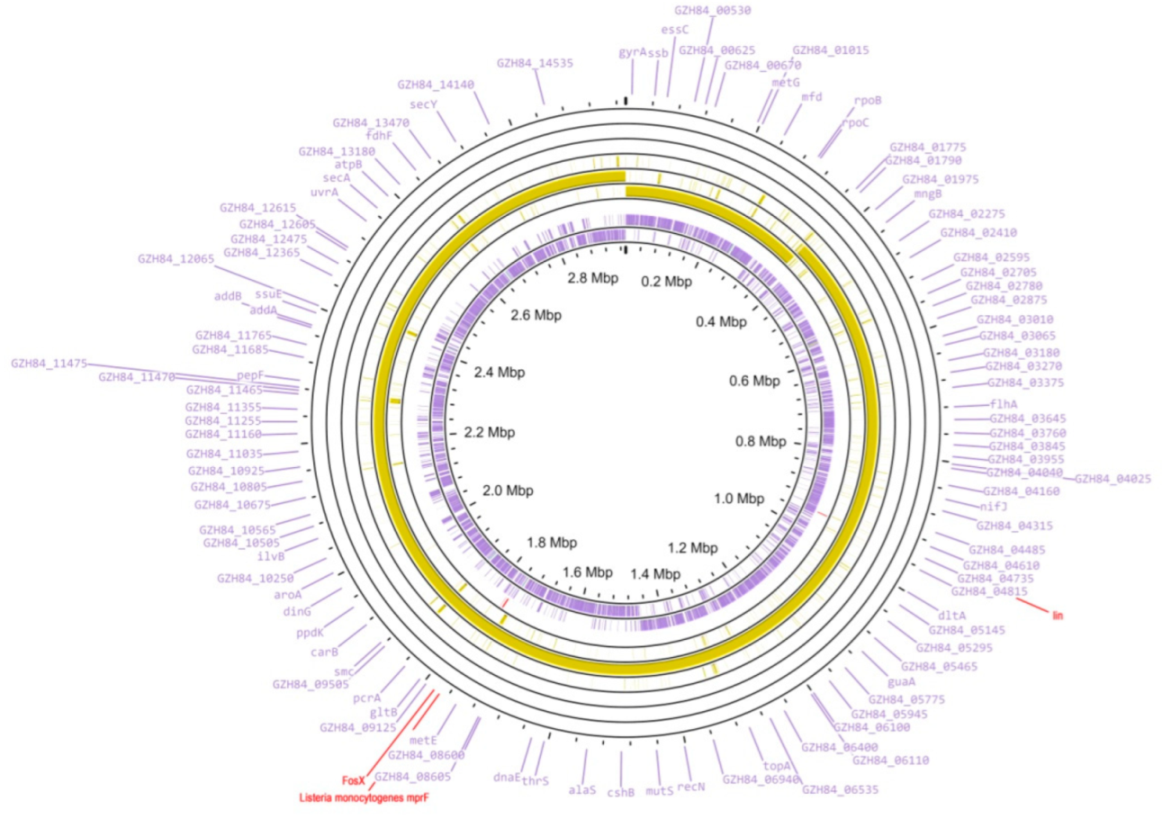
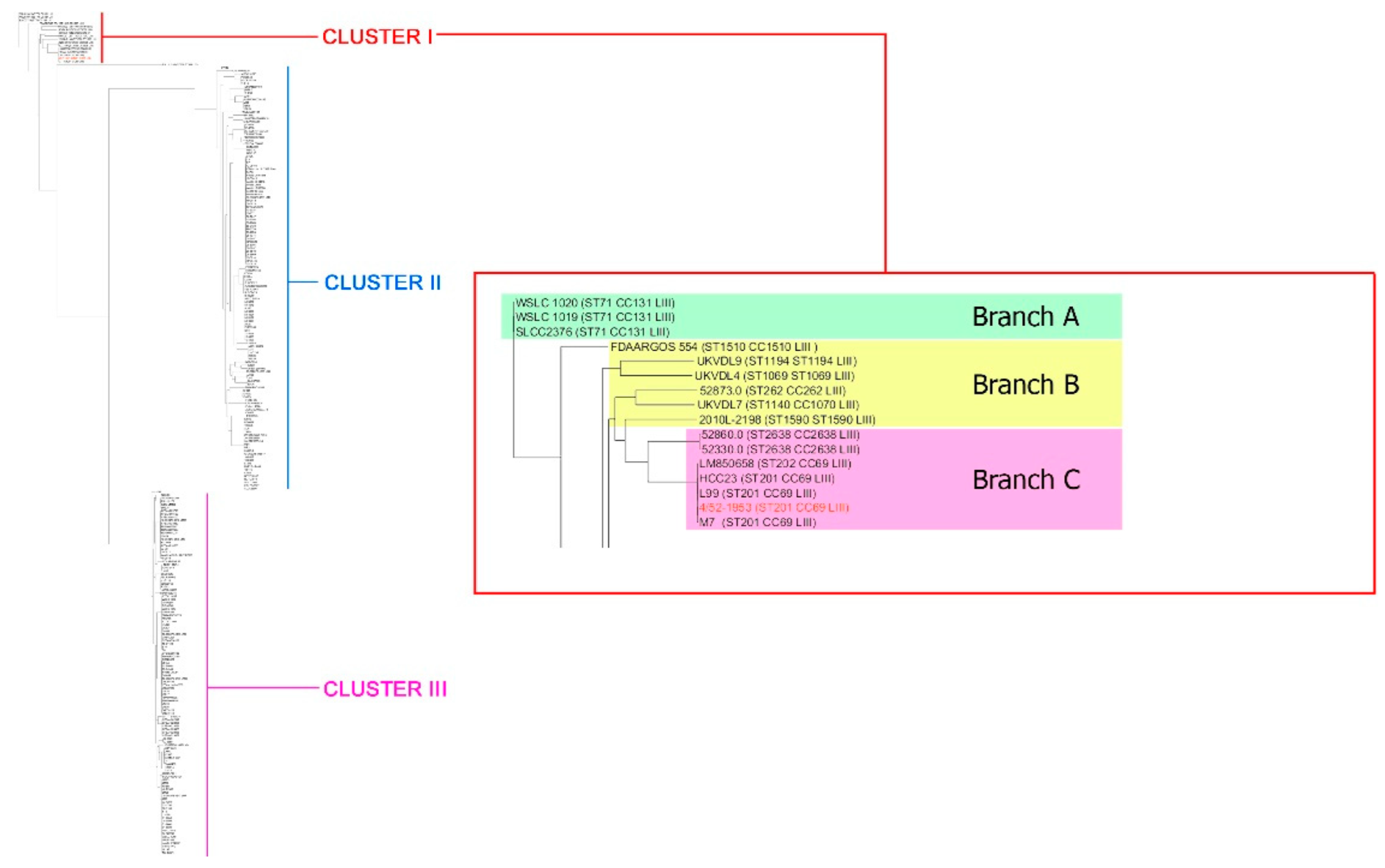
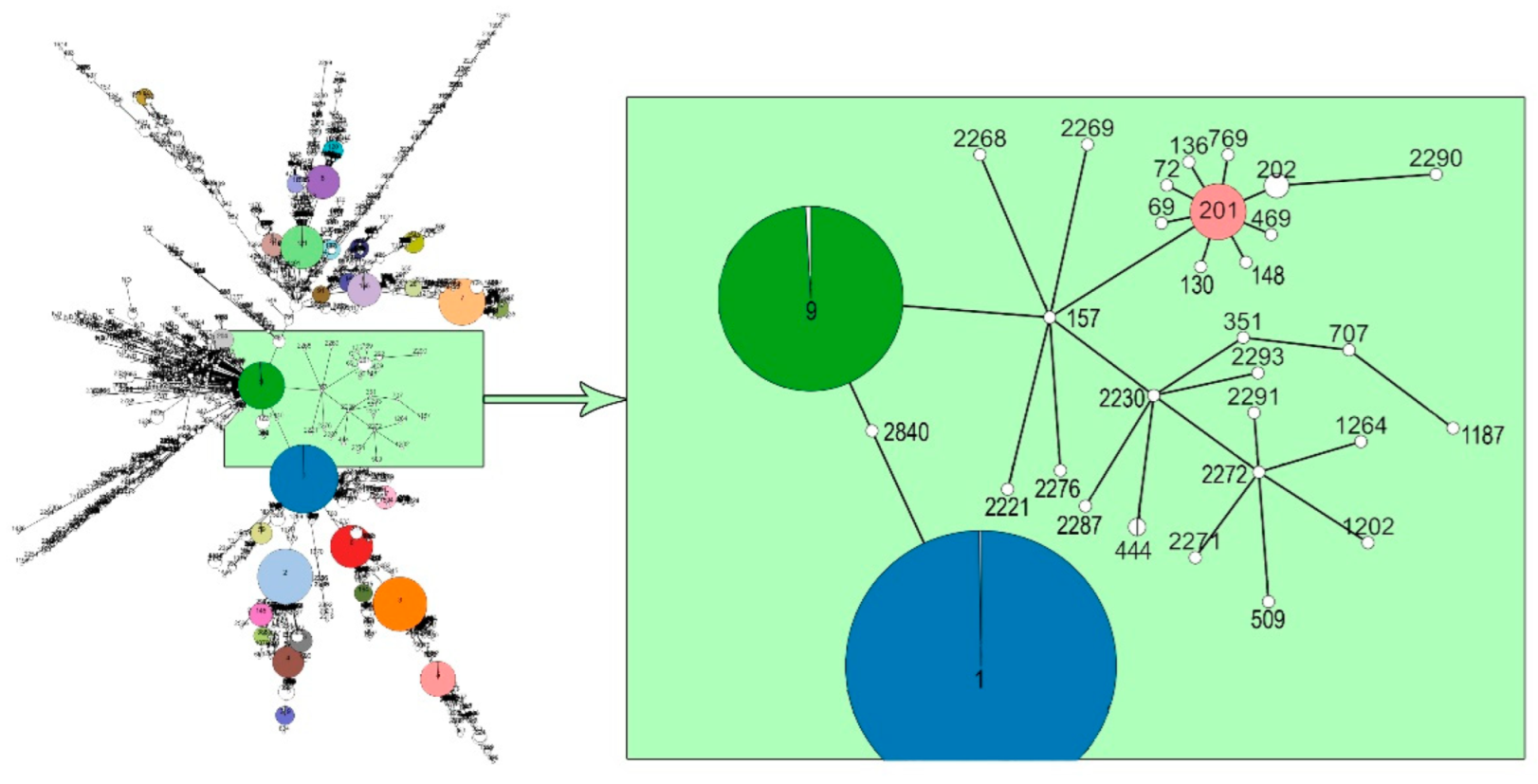
3. Results and Discussions
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hansen, T.B.; Abdalas, S.; Al-Hilali, I.; Hansen, L.T. Predicting the effect of salt on heat tolerance of Listeria monocytogenes in meat and fish products. Int. J. Food Microbiol. 2021, 352, 109265. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos, L.R.; Alia, A.; Martin, I.; Gottardo, F.M.; Rodrigues, L.B.; Borges, K.A.; Furian, T.Q.; Cordoba, J.J. Antimicrobial activity of essential oils and natural plant extracts against Listeria monocytogenes in a dry-cured ham-based model. J. Sci. Food Agric. 2022, 102, 1729–1735. [Google Scholar] [CrossRef] [PubMed]
- Du, J.; Liu, J.; Liu, K.; Zhao, D.; Sagratini, G.; Tao, J.; Bai, Y. Development of a fluorescent test strip sensor based on surface positively-charged magnetic bead separation for the detection of Listeria monocytogenes. Anal. Methods 2022, 14, 2188–2194. [Google Scholar] [CrossRef] [PubMed]
- Felix, B.; Sevellec, Y.; Palma, F.; Douarre, P.E.; Felten, A.; Radomski, N.; Mallet, L.; Blanchard, Y.; Leroux, A.; Soumet, C.; et al. A European-wide dataset to uncover adaptive traits of Listeria monocytogenes to diverse ecological niches. Sci Data 2022, 9, 190. [Google Scholar] [CrossRef] [PubMed]
- Hellstrom, S.; Laukkanen, R.; Siekkinen, K.M.; Ranta, J.; Maijala, R.; Korkeala, H. Listeria monocytogenes contamination in pork can originate from farms. J. Food Prot. 2010, 73, 641–648. [Google Scholar] [CrossRef]
- Hazards, E.P.o.B.; Ricci, A.; Allende, A.; Bolton, D.; Chemaly, M.; Davies, R.; Fernandez Escamez, P.S.; Girones, R.; Herman, L.; Koutsoumanis, K.; et al. Listeria monocytogenes contamination of ready-to-eat foods and the risk for human health in the EU. EFSA J. 2018, 16, e05134. [Google Scholar] [CrossRef]
- Muhterem-uyar, M.; Dalmasso, M.; Sorin, A.; Manios, S.G.; Hernandez, M.; Kapetanakou, A.E.; Melero, B.; Minarovi, J.; Stessl, B.; Skandamis, P.N.; et al. Environmental sampling for Listeria monocytogenes control in food processing facilities reveals three contamination scenarios. Food Control. 2015, 51, 94–107. [Google Scholar] [CrossRef]
- Headley, S.A.; Fritzen, J.T.; Queiroz, G.R.; Oliveira, R.A.; Alfieri, A.F.; Di Santis, G.W.; Lisboa, J.A.; Alfieri, A.A. Molecular characterization of encephalitic bovine listeriosis from southern Brazil. Trop. Anim. Health Prod. 2014, 46, 19–25. [Google Scholar] [CrossRef]
- CDC. National Center for Emerging and Zoonotic Infectious Diseases. National Enteric Disease Surveillance: The Listeria Initiative. 2016; pp. 1–2. Available online: https://www.cdc.gov/listeria/pdf/listeriainitiativeoverview_508.pdf (accessed on 3 June 2022).
- Spyrou, N.; Anderson, M.; Foale, R. Listeria endocarditis: Current management and patient outcome—World literature review. Heart 1997, 77, 380–383. [Google Scholar] [CrossRef] [Green Version]
- Aubin, G.G.; Boutoille, D.; Bourcier, R.; Caillon, J.; Lepelletier, D.; Bemer, P.; Corvec, S. Unusual Case of Spondylodiscitis due to Listeria monocytogenes. J. Bone Jt Infect. 2016, 1, 7–9. [Google Scholar] [CrossRef]
- Randrianarisoa, R.M.F.; Ramanandafy, H.; Mania, A.; Benelhadj, A.D.; Clement, M.; Sabbagh, L.; Vernet, P.; Monjanel, H.; Trouillier, S. Listeria endocarditis and spondylodiscitis: A case report and review of the literature. Clin. Case Rep. 2022, 10, e05899. [Google Scholar] [CrossRef] [PubMed]
- Schaefer, K.; Austhof, E.; Boyd, K.; Armstrong, A.; Hoffman, S.; Pogreba-Brown, K. Septicemia Due to Listeria monocytogenes Infection: A systematic review and meta-analysis. Foodborne Pathog. Dis. 2022, 19, 104–114. [Google Scholar] [CrossRef] [PubMed]
- de Noordhout, C.M.; Devleesschauwer, B.; Angulo, F.J.; Verbeke, G.; Haagsma, J.; Kirk, M.; Havelaar, A.; Speybroeck, N. The global burden of listeriosis: A systematic review and meta-analysis. Lancet Infect. Dis. 2014, 14, 1073–1082. [Google Scholar] [CrossRef] [Green Version]
- Charlier, C.; Perrodeau, E.; Leclercq, A.; Cazenave, B.; Pilmis, B.; Henry, B.; Lopes, A.; Maury, M.M.; Moura, A.; Goffinet, F.; et al. Clinical features and prognostic factors of listeriosis: The MONALISA national prospective cohort study. Lancet Infect. Dis. 2017, 17, 510–519. [Google Scholar] [CrossRef]
- Truchet, L.; Walland, J.; Wuthrich, D.; Boujon, C.L.; Posthaus, H.; Bruggmann, R.; Schupbach-Regula, G.; Oevermann, A.; Seuberlich, T. Neuropathological survey reveals underestimation of the prevalence of neuroinfectious diseases in cattle in Switzerland. Vet. Microbiol. 2017, 208, 137–145. [Google Scholar] [CrossRef] [PubMed]
- Oevermann, A.; Botteron, C.; Seuberlich, T.; Nicolier, A.; Friess, M.; Doherr, M.G.; Heim, D.; Hilbe, M.; Zimmer, K.; Zurbriggen, A.; et al. Neuropathological survey of fallen stock: Active surveillance reveals high prevalence of encephalitic listeriosis in small ruminants. Vet. Microbiol. 2008, 130, 320–329. [Google Scholar] [CrossRef]
- Lu, L.; Li, P.; Pan, T.; Feng, X. Pathogens responsible for early-onset sepsis in Suzhou, China. Jpn J. Infect. Dis 2020, 73, 148–152. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization. Listeriosis, fact- sheets. WHO. 2018. Available online: https://www.who.int/news-room/fact-sheets/detail/listeriosis (accessed on 3 June 2022).
- Shakuntala, I.; Prince Milton, A.A.; Das, S.; Ghatak, S.; Sanjukta, R.; Puro, K.U.; Pegu, R.K.; Doijad, S.P.; Barbuddhe, S.B.; Sen, A. Pulsed-field gel electrophoresis fingerprinting of Listeria monocytogenes isolates recovered from foods of animal origin and fishes in North-Eastern India. Vet. Res. Forum 2022, 13, 133–139. [Google Scholar] [CrossRef]
- Liu, D.; Lawrence, M.L.; Wiedmann, M.; Gorski, L.; Mandrell, R.E.; Ainsworth, A.J.; Austin, F.W. Listeria monocytogenes subgroups IIIA, IIIB, and IIIC delineate genetically distinct populations with varied pathogenic potential. J. Clin. Microbiol. 2006, 44, 4229–4233. [Google Scholar] [CrossRef] [Green Version]
- Ward, T.J.; Ducey, T.F.; Usgaard, T.; Dunn, K.A.; Bielawski, J.P. Multilocus genotyping assays for single nucleotide polymorphism-based subtyping of Listeria monocytogenes isolates. Appl. Environ. Microbiol. 2008, 74, 7629–7642. [Google Scholar] [CrossRef] [Green Version]
- Rasmussen, O.F.; Skouboe, P.; Dons, L.; Rossen, L.; Olsen, J.E. Listeria monocytogenes exists in at least three evolutionary lines: Evidence from flagellin, invasive associated protein and listeriolysin O genes. Microbiology 1995, 141, 2053–2061. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roberts, A.; Nightingale, K.; Jeffers, G.; Fortes, E.; Kongo, J.M.; Wiedmann, M. Genetic and phenotypic characterization of Listeria monocytogenes lineage III. Microbiology 2006, 152, 685–693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orsi, R.H.; den Bakker, H.C.; Wiedmann, M. Listeria monocytogenes lineages: Genomics, evolution, ecology, and phenotypic characteristics. Int. J. Med. Microbiol. 2011, 301, 79–96. [Google Scholar] [CrossRef] [PubMed]
- Camargo, A.C.; Woodward, J.J.; Nero, L.A. The continuous challenge of characterizing the foodborne pathogen Listeria monocytogenes. Foodborne Pathog. Dis. 2016, 13, 405–416. [Google Scholar] [CrossRef]
- Van Walle, I.; Bjorkman, J.T.; Cormican, M.; Dallman, T.; Mossong, J.; Moura, A.; Pietzka, A.; Ruppitsch, W.; Takkinen, J.; European Listeria WGS typing group. Retrospective validation of whole genome sequencing-enhanced surveillance of listeriosis in Europe, 2010 to 2015. Euro Surveill 2018, 23, 1700798. [Google Scholar] [CrossRef] [Green Version]
- Acciari, V.A.; Ruolo, A.; Torresi, M.; Ricci, L.; Pompei, A.; Marfoglia, C.; Valente, F.M.; Centorotola, G.; Conte, A.; Salini, R.; et al. Genetic diversity of Listeria monocytogenes strains contaminating food and food producing environment as single based sample in Italy (retrospective study). Int. J. Food Microbiol. 2022, 366, 109562. [Google Scholar] [CrossRef]
- Nichols, M.; Conrad, A.; Whitlock, L.; Stroika, S.; Strain, E.; Weltman, A.; Johnson, L.; DeMent, J.; Reporter, R.; Williams, I. Short communication: Multistate outbreak of Listeria monocytogenes infections retrospectively linked to unpasteurized milk using whole-genome sequencing. J. Dairy Sci 2020, 103, 176–178. [Google Scholar] [CrossRef] [Green Version]
- Papic, B.; Kusar, D.; Zdovc, I.; Golob, M.; Pate, M. Retrospective investigation of listeriosis outbreaks in small ruminants using different analytical approaches for whole genome sequencing-based typing of Listeria monocytogenes. Infect. Genet. Evol. 2020, 77, 104047. [Google Scholar] [CrossRef]
- Chenal-Francisque, V.; Lopez, J.; Cantinelli, T.; Caro, V.; Tran, C.; Leclercq, A.; Lecuit, M.; Brisse, S. Worldwide distribution of major clones of Listeria monocytogenes. Emerg. Infect. Dis. 2011, 17, 1110–1112. [Google Scholar] [CrossRef]
- Maesaar, M.; Mamede, R.; Elias, T.; Roasto, M. Retrospective use of whole-genome sequencing expands the multicountry outbreak cluster of Listeria monocytogenes ST1247. Int. J. Genom. 2021, 2021, 6636138. [Google Scholar] [CrossRef]
- Lopez-Valladares, G.; Tham, W.; Parihar, V.S.; Helmersson, S.; Andersson, B.; Ivarsson, S.; Johansson, C.; Ringberg, H.; Tjernberg, I.; Henriques-Normark, B.; et al. Human isolates of Listeria monocytogenes in Sweden during half a century (1958–2010). Epidemiol. Infect. 2014, 142, 2251–2260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bespalova, T.Y.; Mikhaleva, T.V.; Meshcheryakova, N.Y.; Kustikova, O.V.; Matovic, K.; Dmitric, M.; Zaitsev, S.S.; Khizhnyakova, M.A.; Feodorova, V.A. Novel sequence types of Listeria monocytogenes of different origin obtained in the Republic of Serbia. Microorganisms 2021, 9, 1289. [Google Scholar] [CrossRef] [PubMed]
- Stover, B.C.; Muller, K.F. TreeGraph 2: Combining and visualizing evidence from different phylogenetic analyses. BMC Bioinform. 2010, 11, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Z.; Alikhan, N.F.; Sergeant, M.J.; Luhmann, N.; Vaz, C.; Francisco, A.P.; Carrico, J.A.; Achtman, M. GrapeTree: Visualization of core genomic relationships among 100,000 bacterial pathogens. Genome Res. 2018, 28, 1395–1404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuenne, C.; Billion, A.; Mraheil, M.A.; Strittmatter, A.; Daniel, R.; Goesmann, A.; Barbuddhe, S.; Hain, T.; Chakraborty, T. Reassessment of the Listeria monocytogenes pan-genome reveals dynamic integration hotspots and mobile genetic elements as major components of the accessory genome. BMC Genom. 2013, 14, 47. [Google Scholar] [CrossRef] [Green Version]
- Albrecht, T.M.; Kucerova, Z.; D’Orazio, S.E.F. Genome sequences of neurotropic lineage III Listeria monocytogenes isolates UKVDL9 and 2010L-2198. Microbiol. Resour. Announc. 2021, 10, e00228-21. [Google Scholar] [CrossRef] [PubMed]
- Heidarlo, M.N.; Lotfollahi, L.; Yousefi, S.; Lohrasbi, V.; Irajian, G.; Talebi, M. Analysis of virulence genes and molecular typing of Listeria monocytogenes isolates from human, food, and livestock from 2008 to 2016 in Iran. Trop. Anim. Health Prod. 2021, 53, 127. [Google Scholar] [CrossRef]
- Moura, A.; Criscuolo, A.; Pouseele, H.; Maury, M.M.; Leclercq, A.; Tarr, C.; Bjorkman, J.T.; Dallman, T.; Reimer, A.; Enouf, V.; et al. Whole genome-based population biology and epidemiological surveillance of Listeria monocytogenes. Nat. Microbiol. 2016, 2, 16185. [Google Scholar] [CrossRef]
- Chen, J.; Luo, X.; Jiang, L.; Jin, P.; Wei, W.; Liu, D.; Fang, W. Molecular characteristics and virulence potential of Listeria monocytogenes isolates from Chinese food systems. Food Microbiol. 2009, 26, 103–111. [Google Scholar] [CrossRef]
- Chen, J.; Xia, Y.; Cheng, C.; Fang, C.; Shan, Y.; Jin, G.; Fang, W. Genome sequence of the nonpathogenic Listeria monocytogenes serovar 4a strain M7. J. Bacteriol. 2011, 193, 5019–5020. [Google Scholar] [CrossRef] [Green Version]
- Cao, X.; Wang, Y.; Wang, Y.; Ye, C. Isolation and characterization of Listeria monocytogenes from the black-headed gull feces in Kunming, China. J. Infect. Public Health 2018, 11, 59–63. [Google Scholar] [CrossRef] [PubMed]
- Gan, L.; Cao, X.; Wang, Y.; Wang, Y.; Jiang, H.; Lan, R.; Xu, J.; Ye, C. Carriage and potential long distance transmission of Listeria monocytogenes by migratory black-headed gulls in Dianchi Lake, Kunming. Emerg. Microbes Infect. 2019, 8, 1195–1204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caruso, M.; Fraccalvieri, R.; Pasquali, F.; Santagada, G.; Latorre, L.M.; Difato, L.M.; Miccolupo, A.; Normanno, G.; Parisi, A. Antimicrobial susceptibility and multilocus sequence typing of Listeria monocytogenes isolated over 11 years from food, humans, and the environment in Italy. Foodborne Pathog. Dis. 2020, 17, 284–294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dar, D.; Shamir, M.; Mellin, J.R.; Koutero, M.; Stern-Ginossar, N.; Cossart, P.; Sorek, R. Term-seq reveals abundant ribo-regulation of antibiotics resistance in bacteria. Science 2016, 352, aad9822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thedieck, K.; Hain, T.; Mohamed, W.; Tindall, B.J.; Nimtz, M.; Chakraborty, T.; Wehland, J.; Jansch, L. The MprF protein is required for lysinylation of phospholipids in listerial membranes and confers resistance to cationic antimicrobial peptides (CAMPs) on Listeria monocytogenes. Mol. Microbiol. 2006, 62, 1325–1339. [Google Scholar] [CrossRef] [Green Version]
- Fillgrove, K.L.; Pakhomova, S.; Schaab, M.R.; Newcomer, M.E.; Armstrong, R.N. Structure and mechanism of the genomically encoded fosfomycin resistance protein, FosX, from Listeria monocytogenes. Biochemistry 2007, 46, 8110–8120. [Google Scholar] [CrossRef]
- Haubert, L.; Kremer, F.S.; da Silva, W.P. Whole-genome sequencing identification of a multidrug-resistant Listeria monocytogenes serotype 1/2a isolated from fresh mixed sausage in southern Brazil. Infect. Genet. Evol. 2018, 65, 127–130. [Google Scholar] [CrossRef]
- Weisblum, B. Erythromycin resistance by ribosome modification. Antimicrob. Agents Chemother. 1995, 39, 577–585. [Google Scholar] [CrossRef] [Green Version]
- Schubert, W.D.; Urbanke, C.; Ziehm, T.; Beier, V.; Machner, M.P.; Domann, E.; Wehland, J.; Chakraborty, T.; Heinz, D.W. Structure of internalin, a major invasion protein of Listeria monocytogenes, in complex with its human receptor E-cadherin. Cell 2002, 111, 825–836. [Google Scholar] [CrossRef] [Green Version]
- Bierne, H.; Cossart, P. InlB, a surface protein of Listeria monocytogenes that behaves as an invasin and a growth factor. J. Cell Sci. 2002, 115, 3357–3367. [Google Scholar] [CrossRef]
- Astashkin, E.I.; Alekseeva, E.A.; Borzenkov, V.N.; Kislichkina, A.A.; Mukhina, T.N.; Platonov, M.E.; Svetoch, E.A.; Shepelin, A.P.; Fursova, N.K. Molecular genetic characteristics of polyresistant Listeria monocytogenes strains and identification of new sequence types. Mol. Genet. Microbiol. Virol. 2021, 39, 3–13. [Google Scholar] [CrossRef]
- Maury, M.M.; Bracq-Dieye, H.; Huang, L.; Vales, G.; Lavina, M.; Thouvenot, P.; Disson, O.; Leclercq, A.; Brisse, S.; Lecuit, M. Hypervirulent Listeria monocytogenes clones’ adaption to mammalian gut accounts for their association with dairy products. Nat. Commun. 2019, 10, 2488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Autret, N.; Raynaud, C.; Dubail, I.; Berche, P.; Charbit, A. Identification of the agr locus of Listeria monocytogenes: Role in bacterial virulence. Infect. Immun. 2003, 71, 4463–4471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sumrall, E.; Klumpp, J.; Shen, Y.; Loessner, M.J. Genome Sequences of Five Nonvirulent Listeria monocytogenes Serovar 4 Strains. Genome Announc. 2016, 4, e00179-16. [Google Scholar] [CrossRef] [Green Version]
- Senay, T.E.; Ferrell, J.L.; Garrett, F.G.; Albrecht, T.M.; Cho, J.; Alexander, K.L.; Myers-Morales, T.; Grothaus, O.F.; D’Orazio, S.E.F. Neurotropic Lineage III Strains of Listeria monocytogenes Disseminate to the Brain without Reaching High Titer in the Blood. MSphere 2020, 5, e00871-20. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zaitsev, S.S.; Khizhnyakova, M.A.; Feodorova, V.A. Retrospective Investigation of the Whole Genome of the Hypovirulent Listeria monocytogenes Strain of ST201, CC69, Lineage III, Isolated from a Piglet with Fatal Neurolisteriosis. Microorganisms 2022, 10, 1442. https://doi.org/10.3390/microorganisms10071442
Zaitsev SS, Khizhnyakova MA, Feodorova VA. Retrospective Investigation of the Whole Genome of the Hypovirulent Listeria monocytogenes Strain of ST201, CC69, Lineage III, Isolated from a Piglet with Fatal Neurolisteriosis. Microorganisms. 2022; 10(7):1442. https://doi.org/10.3390/microorganisms10071442
Chicago/Turabian StyleZaitsev, Sergey S., Mariya A. Khizhnyakova, and Valentina A. Feodorova. 2022. "Retrospective Investigation of the Whole Genome of the Hypovirulent Listeria monocytogenes Strain of ST201, CC69, Lineage III, Isolated from a Piglet with Fatal Neurolisteriosis" Microorganisms 10, no. 7: 1442. https://doi.org/10.3390/microorganisms10071442