
Endophytic Bacteria and Essential Oil from Origanum vulgare ssp. vulgare Share Some VOCs with an Antibacterial Activity

 , , , , and
, , , , and 
Abstract
:1. Introduction
2. Results
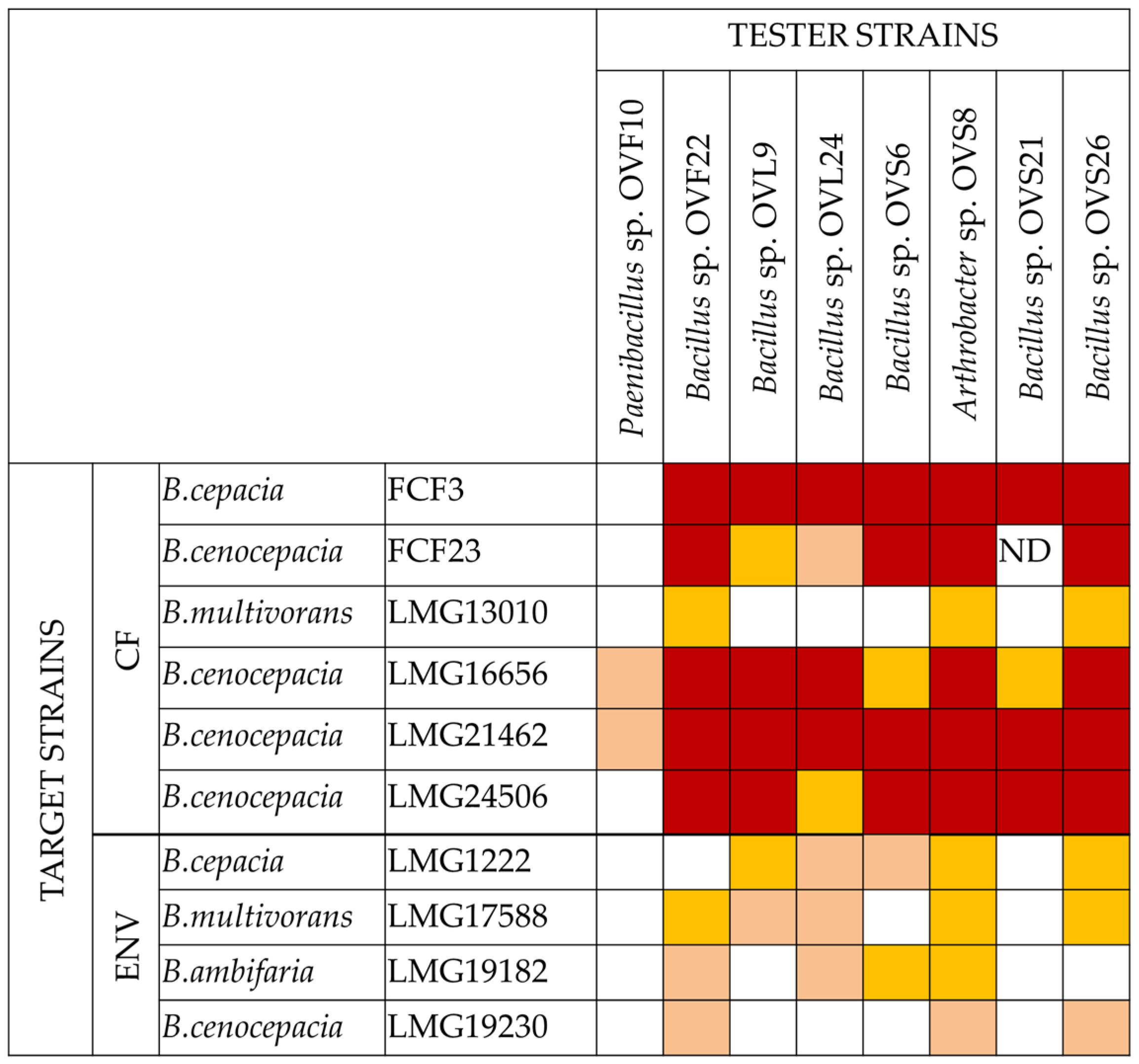
2.1. VOCs Synthesized by O. vulgare Endophytes Possess Antibacterial Activity
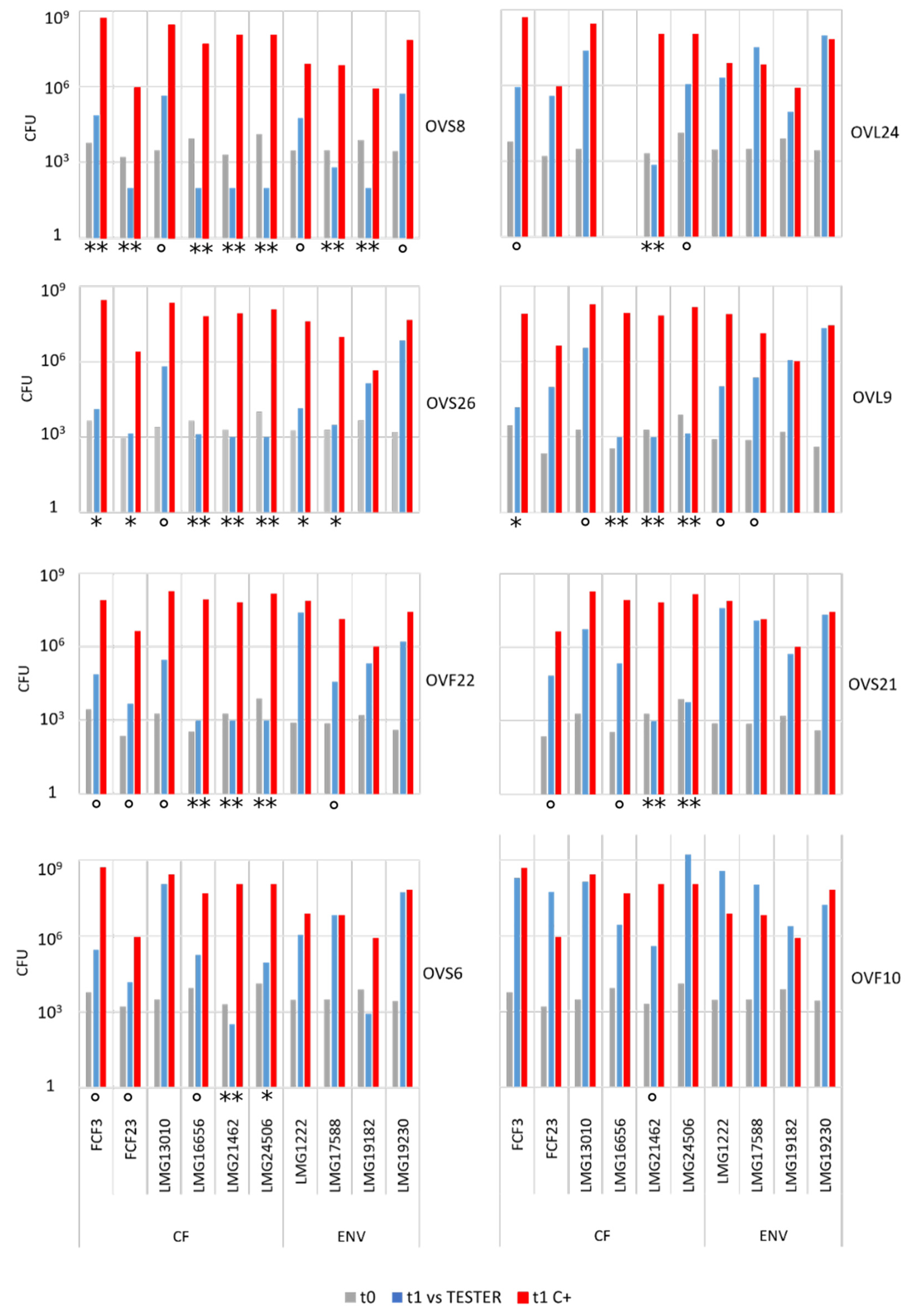
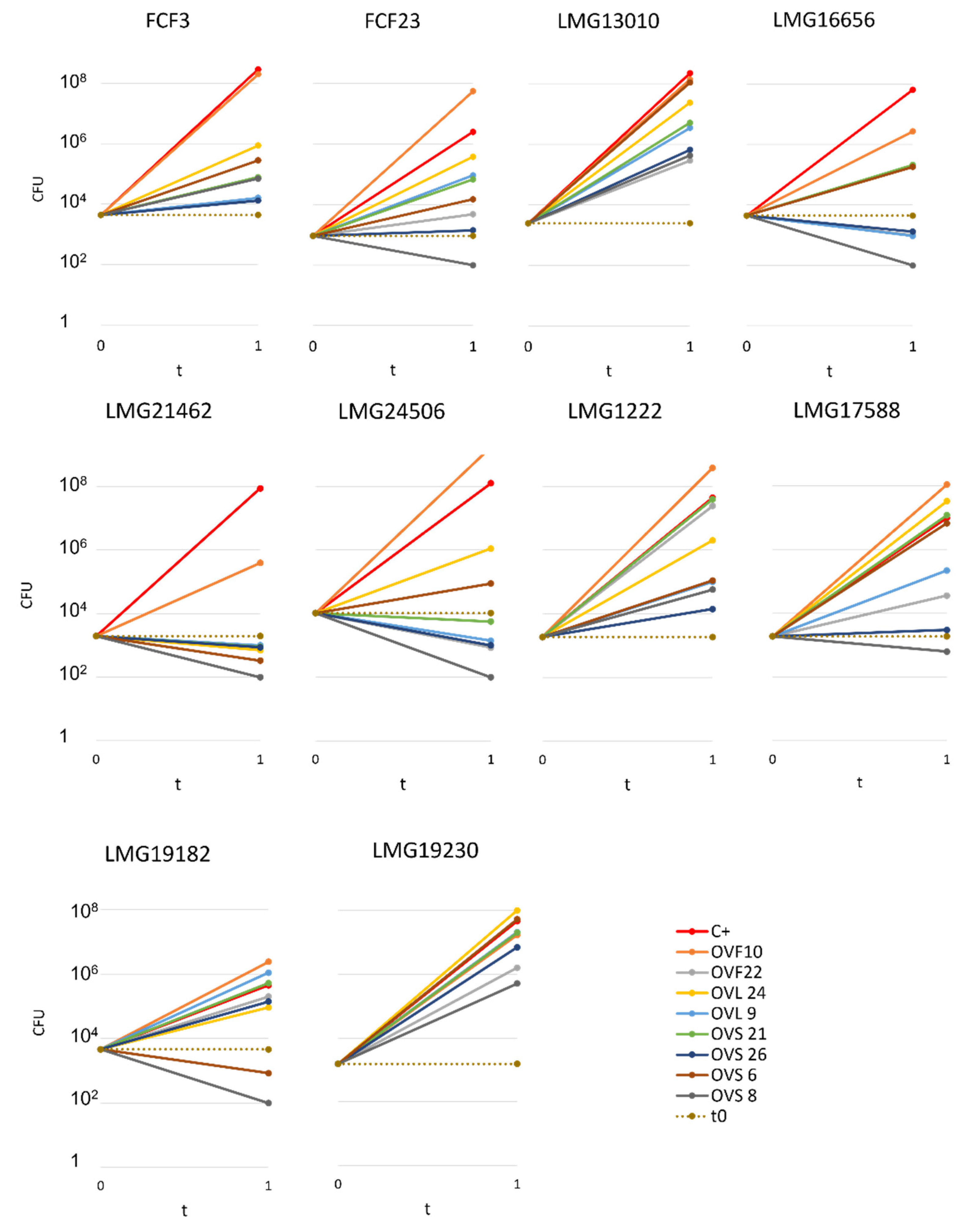
2.2. Quantitative Analysis of VOCs Antibacterial Activity
- (i)
- All the tester strains were able to inhibit the growth of at least one of the target strains.
- (ii)
- The VOCs synthesized by the eight endophytes exhibited a different ability to interfere with the growth of target strains.
- (iii)
- The most active endophytic strain was Arthrobacter sp. OVS8, whose VOCs were able to strongly antagonize the growth of all the Bcc strains, showing a bactericidal activity against seven out of ten Bcc strains (see Figure 2 and Figure 3), and a growth reducing activity on the remaining three targets. Overall, the mean number of viable cells of target strains was 0.17% in the presence of strain OVS8 with respect to the control plates (i.e., in the absence of the tester strain).
- (iv)
- On the other site, Paenibacillus sp. OVF10 exhibited the weakest antibacterial potential, being able to antagonize only the growth of LMG16656 and LMG21462 strains. These two targets, together with LMG24506, appeared to be the most sensitive to the VOCs produced by all the endophytes. It is also quite interesting that OVF10 is the only tester strain able to stimulate the growth of four targets (FCF23, LMG24506, LMG1222, and LMG17588).
- (v)
- The remaining six tester strains, all belonging to the Bacillus genus, showed a different inhibitory effect on Bcc strains. All Bacillus strains induced a bactericidal effect against at least one of the target strains, with OVS26 and OVF22 exerting the strongest antibacterial activity.
- (vi)
- The clinical Bcc strains isolated from CF patients were more sensitive to the endophytic VOCs than the environmental ones.
- (vii)
- The two B. multivorans strains (one of clinical origin and the other one isolated from the environment) were less sensitive to the endophytic VOCs than the other eight Bcc strains belonging to the B. cepacia, B. cenocepacia, and B. ambifaria species.
2.3. SPME-GC/MS Analysis of VOCs
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains and Growth Conditions
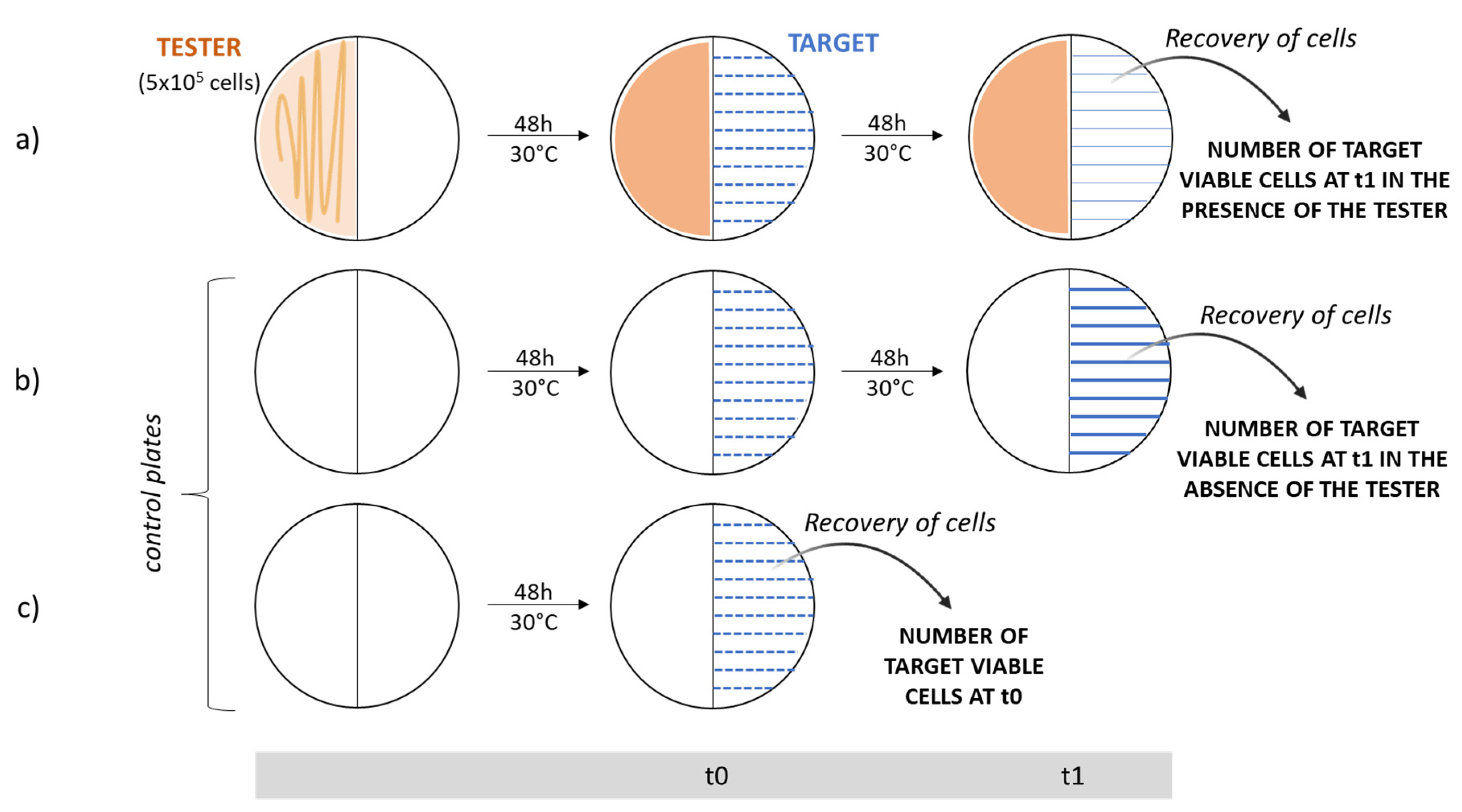
4.2. Analysis of the Antibacterial VOC Production through Cross-Streaking
4.3. Quantitative Analysis of VOC Antibacterial Activities
- i.
- Single colonies from each endophyte were separately inoculated in 10 mL of fresh tryptic soy broth (TSB) medium (BioLife) and incubated at 30 °C overnight. The next morning, cells were counted in a Burker chamber and diluted in saline solution (0.9% w/v NaCl) in order to evenly spread about 5 × 105 cells on one half of a TSA Petri plate with septum. A viable count of the cells plated was performed.
- ii.
- Plates were incubated at 30 °C for 48 h to allow the production of volatile antibacterial organic compounds.
- iii.
- Afterwards, a few colonies of each target strain (previously isolated and grown at 37 °C for 48 h on LB plates) were suspended in 100 µL of saline solution (0.9% w/v NaCl); then, a 1:100 dilution was prepared. The dilution was streaked 10 times perpendicularly to the septum on the opposite half of Petri dishes where the endophytes had grown.
- iv.
- The same procedure was repeated on two TSA plates in the absence of the tester strains; these plates represented the control ones, which permitted us to determine the number of viable cells grown in the absence of the tester at the beginning (t0) and at the end of the experiment (t1).
- v.
- All these plates were incubated at 30 °C for 2 days.
- vi.
- To determine the number of cells just plated, the same dilutions were streaked 10 times on half of a Petri dish and cells were immediately recovered (t0) in 2 mL of saline solution with a spatula. The suspensions obtained were spread onto LB plates and incubated at 37 °C for 48 h to determine the viable titer.
- vii.
- After 2 days, from cross-streaking plates and growth control plates, target strain cells were recovered in saline solution, as previously described. The suspensions obtained were properly diluted, spread onto LB plates, and incubated at 37 °C for 48 h for the viable count.

4.4. Identification of VOCs by Means of SPME-GC/MS
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Vining, L.C. Functions of secondary metabolites. Annu. Rev. Microbiol. 1990, 44, 395–427. [Google Scholar] [CrossRef] [PubMed]
- Romoli, R.; Papaleo, M.C.; De Pascale, D.; Tutino, M.L.; Michaud, L.; LoGiudice, A.; Fani, R.; Bartolucci, G. GC–MS volatolomic approach to study the antimicrobial activity of the Antarctic bacterium Pseudoalteromonas sp. TB41. Metabolomics 2014, 10, 42–51. [Google Scholar] [CrossRef]
- Schulz-Bohm, K.; Martín-Sánchez, L.; Garbeva, P. Microbial Volatiles: Small Molecules with an Important Role in Intra and Inter-Kingdom Interactions. Front. Microbiol. 2017, 8, 2484. [Google Scholar] [CrossRef] [PubMed]
- Claeson, A. Volatile Organic Compounds from Microorganisms-Identification and Health Effects. Ph.D. Thesis, Umeå Universitet, Umeå, Sweden, 2006. [Google Scholar]
- Sunesson, A.; Nilsson, C.; Carlson, R.; Blomquist, G.; Andersson, B. Influence of temperature, oxygen and carbon dioxide levels on the production of volatile metabolites from Streptomyces albidoflavus cultivated on gypsum board and tryptone glucose extract agar. Ann. Occup. Hyg. 1997, 41, 393–413. [Google Scholar] [CrossRef] [Green Version]
- Leff, J.W.; Fierer, N. Volatile organic compound (VOC) emissions from soil and litter samples. Soil Biol. Biochem. 2008, 40, 1629–1636. [Google Scholar] [CrossRef]
- Maffei, M.E.; Gertsch, J.; Appendino, G. Plant volatiles: Production, function and pharmacology. Nat. Prod. Rep. 2011, 28, 1359. [Google Scholar] [CrossRef]
- Kanchiswamy, C.N.; Malnoy, M.; Maffei, M. Chemical diversity of microbial volatiles and their potential for plant growth and productivity. Front. Plant Sci. 2015, 6, 151. [Google Scholar] [CrossRef] [Green Version]
- Effmert, U.; Kalderás, J.; Warnke, R.; Piechulla, B. Volatile mediated interactions between bacteria and fungi in the soil. J. Chem. Ecol. 2012, 38, 665–703. [Google Scholar] [CrossRef]
- Thomas, G.; Withall, D.; Birkett, M. Harnessing microbial volatiles to replace pesticides and fertilizers. Microb. Biotechnol. 2020, 13, 1366–1376. [Google Scholar] [CrossRef]
- Audrain, B.; Farag, M.A.; Ryu, C.-M.; Ghigo, J.-M. Role of bacterial volatile compounds in bacterial biology. FEMS Microbiol. Rev. 2015, 39, 222–233. [Google Scholar] [CrossRef] [Green Version]
- Kai, M.; Effmert, U.; Berg, G.; Piechulla, B. Volatiles of bacterial antagonists inhibit mycelial growth of the plant pathogen Rhizoctonia solani. Arch. Microbial. 2007, 187, 351–360. [Google Scholar] [CrossRef]
- Minerdi, D.; Maggini, V.; Fani, R. Volatile organic compounds: From figurants to leading actors in fungal symbiosis. FEMS Microbiol. Ecol. 2021, 97, fiab067. [Google Scholar] [CrossRef]
- Papaleo, M.C.; Fondi, M.; Maida, I.; Perrin, E.; Giudice, A.L.; Michaud, L.; Mangano, S.; Bartolucci, G.; Romoli, R.; Fani, R. Sponge associated microbial Antarctic communities exhibiting antimicrobial activity against Burkholderia cepacia complex bacteria. Biotechnol. Adv. 2012, 30, 272–293. [Google Scholar] [CrossRef]
- Romoli, R.; Papaleo, M.; de Pascale, D.; Tutino, M.; Michaud, L.; LoGiudice, A.; Fani, R.; Bartolucci, G. Characterization of the volatile profile of Antarctic bacteria by using solid phase microextraction-gas chromatography-mass spectrometry. J. Mass Spectrom. 2011, 46, 1051–1059. [Google Scholar] [CrossRef]
- Papaleo, M.C.; Romoli, R.; Bartolucci, G.; Maida, I.; Perrin, E.; Fondi, M.; Orlandini, V.; Mengoni, A.; Emiliani, G.; Tutino, M.L.; et al. Bioactive volatile organic compounds from Antarctic (sponges) bacteria. New Biotechnol. 2013, 30, 824–838. [Google Scholar] [CrossRef]
- Sannino, F.; Parrilli, E.; Apuzzo, G.A.; de Pascale, D.; Tedesco, P.; Maida, I.; Perrin, E.; Fondi, M.; Fani, R.; Marino, G.; et al. Pseudoalteromonas haloplanktis produces methylamine, a volatile compound active against Burkholderia cepacia complex strains. New Biotechnol. 2017, 35, 13–18. [Google Scholar] [CrossRef]
- Islam, N.; Choi, J.; Baek, K.-H. Antibacterial Activities of Endophytic Bacteria Isolated from Taxus brevifolia against Foodborne Pathogenic Bacteria. Foodborne Pathog. Dis. 2018, 15, 269–276. [Google Scholar] [CrossRef]
- Cardoso, V.; Campos, F.; Santos, A.; Ottoni, M.; Rosa, C.; Almeida, V.; Grael, C.; de Almeida, V.G. Biotechnological applications of the medicinal plant Pseudobrickellia brasiliensis and its isolated endophytic bacteria. J. Appl. Microbiol. 2020, 129, 926–934. [Google Scholar] [CrossRef]
- Chiellini, C.; Maida, I.; Maggini, V.; Bosi, E.; Mocali, S.; Emiliani, G.; Perrin, E.; Firenzuoli, F.; Mengoni, A.; Fani, R. Preliminary data on antibacterial activity of Echinacea purpurea-associated bacterial communities against Burkholderia cepacia complex strains, opportunistic pathogens of Cystic Fibrosis patients. Microbiol. Res. 2017, 196, 34–43. [Google Scholar] [CrossRef]
- Bakkali, F.; Averbeck, S.; Averbeck, D.; Idaomar, M. Biological effects of essential oils—A review. Food Chem. Toxicol. 2008, 46, 446–475. [Google Scholar] [CrossRef]
- Swamy, M.K.; Akhtar, M.S.; Sinniah, U.R. Antimicrobial properties of plant essential oils against human pathogens and their mode of action: An updated review. Evid. Based Complement. Altern. Med. 2016, 2016, 3012462. [Google Scholar] [CrossRef]
- Castronovo, L.M.; Vassallo, A.; Mengoni, A.; Miceli, E.; Bogani, P.; Firenzuoli, F.; Fani, R.; Maggini, V. Medicinal plants and their bacterial microbiota: A review on antimicrobial compounds production for plant and human health. Pathogens 2021, 10, 106. [Google Scholar] [CrossRef]
- Maggini, V.; Miceli, E.; Fagorzi, C.; Maida, I.; Fondi, M.; Perrin, E.; Mengoni, A.; Bogani, P.; Chiellini, C.; Mocali, S.; et al. Antagonism and antibiotic resistance drive a species-specific plant microbiota differentiation in Echinacea spp. FEMS Microbiol. Ecol. 2018, 94, fiy118. [Google Scholar] [CrossRef] [PubMed]
- Maggini, V.; De Leo, M.; Granchi, C.; Tuccinardi, T.; Mengoni, A.; Gallo, E.R.; Biffi, S.; Fani, R.; Pistelli, L.; Firenzuoli, F.; et al. The influence of Echinacea purpurea leaf microbiota on chicoric acid level. Sci. Rep. 2019, 9, 10897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bitas, V.; Kim, H.-S.; Bennett, J.W.; Kang, S. Sniffing on microbes: Diverse roles of microbial volatile organic compounds in plant health. Mol. Plant-Microbe Interact. 2013, 26, 835–843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castronovo, L.M.; Calonico, C.; Ascrizzi, R.; Del Duca, S.; Delfino, V.; Chioccioli, S.; Vassallo, A.; Strozza, I.; De Leo, M.; Biffi, S.; et al. The Cultivable Bacterial Microbiota Associated to the Medicinal Plant Origanum vulgare L.: From Antibiotic Resistance to Growth-Inhibitory Properties. Front. Microbiol. 2020, 11, 862. [Google Scholar] [CrossRef] [PubMed]
- Sousa, E.T.; Rodrigues, F.D.M.; Martins, C.C.; de Oliveira, F.S.; Pereira, P.A.D.P.; de Andrade, J.B. Multivariate optimization and HS-SPME/GC-MS analysis of VOCs in red, yellow and purple varieties of Capsicum chinense sp. peppers. Microchem. J. 2006, 82, 142–149. [Google Scholar] [CrossRef]
- Ossowicki, A.; Jafra, S.; Garbeva, P. The antimicrobial volatile power of the rhizospheric isolate Pseudomonas donghuensis P482. PLoS ONE 2017, 12, e0174362. [Google Scholar] [CrossRef] [Green Version]
- Ross, Z.; O’Gara, A.; Hill, D.; Sleigholme, H.; Maslin, D. Antimicrobial properties of garlic oil against human enteric bacteria: Evaluation of methodologies and comparisons with garlic oil sulfides and garlic powder. Appl. Environ. Microbiol. 2001, 67, 475–480. [Google Scholar] [CrossRef] [Green Version]
- Altaee, N.; Kadhim, M.J.; Hameed, I.H. Detection of volatile compounds produced by Pseudomonas aeruginosa isolated from UTI patients by gas chromatography-mass spectrometry. Int. J. Curr. Pharm. Rev. Res. 2017, 7, 8–24. [Google Scholar]
- Leyva-López, N.; Gutiérrez-Grijalva, E.P.; Vazquez-Olivo, G.; Heredia, J.B. Essential oils of oregano: Biological activity beyond their antimicrobial properties. Molecules 2017, 22, 989. [Google Scholar] [CrossRef] [Green Version]
- Caboni, P.; Ntalli, N.; Aissani, N.; Cavoski, I.; Angioni, A. Nematicidal activity of (E, E)-2, 4-decadienal and (E)-2-decenal from Ailanthus altissima against Meloidogyne javanica. J. Agric. Food Chem. 2012, 60, 1146–1151. [Google Scholar] [CrossRef]
- Stracquadanio, C.; Quiles, J.M.; Meca, G.; Cacciola, S.O. Antifungal activity of bioactive metabolites produced by Trichoderma asperellum and Trichoderma atroviride in liquid medium. J. Fungi 2020, 6, 263. [Google Scholar] [CrossRef]
- Gao, Z.; Zhang, B.; Liu, H.; Han, J.; Zhang, Y. Identification of endophytic Bacillus velezensis ZSY-1 strain and antifungal activity of its volatile compounds against Alternaria solani and Botrytis cinerea. Biol. Control 2017, 105, 27–39. [Google Scholar] [CrossRef]
- Rybakova, D.; Cernava, T.; Koberl, M.; Liebminger, S.; Etremadi, M.; Berg, G. Endophytes-assisted biocontrol: Novel insights in ecology and the mode of action of Paenibacillus. Plant Soil 2016, 405, 125–140. [Google Scholar] [CrossRef]
- Lee, B.; Farag, M.A.; Park, H.B.; Kloepper, J.W.; Lee, S.H.; Ryu, C.-M. Induced resistance by a long-chain bacterial volatile: Elicitation of plant systemic defense by a C13 volatile produced by Paenibacillus polymyxa. PLoS ONE 2012, 7, e48744. [Google Scholar] [CrossRef]
- Kusstatscher, P.; Cernava, T.; Liebminger, S.; Berg, G. Replacing conventional decontamination of hatching eggs with a natural defense strategy based on antimicrobial, volatile pyrazines. Sci. Rep. 2017, 7, 13253. [Google Scholar] [CrossRef] [Green Version]
- Cheng, W.; Yang, J.; Nie, Q.; Huang, D.; Yu, C.; Zheng, L.; Cai, M.; Thomashow, L.S.; Weller, D.M.; Yu, Z.; et al. Volatile organic compounds from Paenibacillus polymyxa KM2501-1 control Meloidogyne incognita by multiple strategies. Sci. Rep. 2017, 7, 16213. [Google Scholar] [CrossRef]
- Keswani, C.; Singh, H.B.; García-Estrada, C.; Caradus, J.; He, Y.-W.; Mezaache-Aichour, S.; Glare, T.R.; Borriss, R.; Sansinenea, E. Antimicrobial secondary metabolites from agriculturally important bacteria as next-generation pesticides. Appl. Microbiol. Biotechnol. 2020, 104, 1013–1034. [Google Scholar] [CrossRef]
- Keswani, C.; Prakash, O.; Bharti, N.; Vílchez, J.I.; Sansinenea, E.; Lally, R.D.; Borriss, R.; Singh, S.P.; Gupta, V.K.; Fraceto, L.F.; et al. Re-addressing the biosafety issues of plant growth promoting rhizobacteria. Sci. Total Environ. 2019, 690, 841–852. [Google Scholar] [CrossRef]
- Mortzfeld, F.B.; Hashem, C.; Vranková, K.; Winkler, M.; Rudroff, F. Pyrazines: Synthesis and Industrial Application of these Valuable Flavor and Fragrance Compounds. Biotechnol. J. 2020, 15, 2000064. [Google Scholar] [CrossRef]
- Lammers, A.; Lalk, M.; Garbeva, P. Air Ambulance: Antimicrobial Power of Bacterial Volatiles. Antibiotics 2022, 11, 109. [Google Scholar] [CrossRef]
- Tabacchioni, S.; Ferri, L.; Manno, G.; Mentasti, M.; Cocchi, P.; Campana, S.; Ravenni, N.; Taccetti, G.; Dalmastri, C.; Chiarini, L.; et al. Use of the gyrB gene to discriminate among species of the Burkholderia cepacia complex. FEMS Microbiol. Lett. 2008, 281, 175–182. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mean (%) | |
|---|---|
| Arthrobacter sp. OVS8 | 0.17 |
| Bacillus sp. OVS26 | 4.14 |
| Bacillus sp. OVF22 | 5.92 |
| Bacillus sp. OVL9 | 19.68 |
| Bacillus sp. OVS6 | 23.83 |
| Bacillus sp. OVS21 | 30.96 |
| Bacillus sp. OVL24 | 82.18 |
| Paenibacillus sp. OVF10 | 1492.42 |
| tR (min) | Compounds | OVL9 | OVL24 | OVS6 | OVS8 | OVS21 | OVS26 | OVF10 | OVF22 |
|---|---|---|---|---|---|---|---|---|---|
| 6.34 |  2-Butene, 2-methyl- | 25.29 | 3.57 | 13.96 | 6.81 | 3.29 | 1.63 | 2.80 | 15.25 |
| 6.54 |  Dimethyl disulfide | 20.28 | 60.51 | 35.14 | 84.55 | 68.41 | 8.41 | 0.28 | 43.00 |
| 7.95 |  Butanoic acid, 3-methyl-, methyl ester | 1.38 | 0.33 | 0.00 | 0.00 | 0.76 | 0.42 | 0.00 | 1.77 |
| 8.64 |  Propanoic acid, 2-methyl- | 2.88 | 0.00 | 0.00 | 0.00 | 0.00 | 4.10 | 0.00 | 1.81 |
| 12.39 |  o-Xylene | 0.40 | 0.36 | 1.56 | 0.07 | 0.24 | 0.35 | 0.01 | 0.30 |
| 12.99 |  p-Xylene | 0.74 | 0.59 | 3.93 | 0.13 | 0.37 | 0.78 | 0.02 | 0.59 |
| 13.44 |  Butanoic acid, 3-methyl- | 16.22 | 2.50 | 4.26 | 0.00 | 2.30 | 35.36 | 0.00 | 2.02 |
| 14.10 |  Butanoic acid, 2-methyl- | 12.26 | 0.46 | 0.00 | 0.00 | 1.59 | 30.47 | 0.00 | 0.96 |
| 14.30 |  1-Butanol, 3-methyl-, acetate | 1.07 | 0.59 | 6.31 | 0.13 | 0.26 | 1.60 | 0.01 | 0.49 |
| 14.40 |  Styrene | 2.54 | 2.75 | 3.46 | 0.57 | 12.99 | 2.81 | 0.05 | 2.48 |
| 16.08 |  Oxime-, methoxy-phenyl- | 3.36 | 3.12 | 8.29 | 0.47 | 0.55 | 3.13 | 0.04 | 6.35 |
| 16.11 |  Pyrazine, 2,5-dimethyl- | 8.12 | 7.19 | 3.36 | 1.33 | 6.29 | 5.15 | 0.08 | 15.30 |
| 17.35 |  α-Pinene | 0.27 | 0.17 | 0.46 | 0.04 | 0.03 | 0.12 | 0.01 | 0.19 |
| 17.99 |  Propanethioic acid, 2,2-dimethyl- | 0.61 | 13.32 | 1.10 | 1.33 | 0.55 | 0.47 | 0.01 | 1.22 |
| 19.84 |  Dimethyl trisulfide | 0.54 | 0.88 | 3.65 | 2.94 | 0.59 | 0.62 | 0.03 | 3.96 |
| 21.09 |  2-Octen-1-ol, (Z)- | 0.64 | 0.82 | 0.16 | 0.04 | 0.26 | 0.27 | 0.39 | 0.74 |
| 22.99 |  3-Carene | 0.59 | 0.35 | 1.23 | 0.13 | 0.19 | 0.35 | 0.01 | 0.45 |
| 24.20 |  p-Cymene | 0.23 | 0.22 | 1.02 | 0.07 | 0.12 | 0.21 | 0.01 | 0.24 |
| 24.58 |  γ-Terpinene | 0.46 | 0.38 | 1.89 | 0.10 | 0.17 | 0.29 | 0.01 | 0.31 |
| 24.74 |  2-Decenal | 0.77 | 0.85 | 1.15 | 0.12 | 0.54 | 0.94 | 0.11 | 0.79 |
| 25.74 |  2-methyl-5-(1-methylethyl)-Pyrazine | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 7.92 | 0.00 |
| 31.40 |  Pyrazine, 3,5-dimethyl-2-i-propyl- | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.04 | 0.00 |
| 32.29 |  2,3-Dimethyl-5-i-propylpyrazine | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.03 | 0.00 |
| 32.64 |  2-Isobutyl-3-methylpyrazine | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.01 | 0.00 |
| 34.11 |  Pyrazine, 2,3-dimethyl-5-(1-methylpropyl)- | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.21 | 0.00 |
| 36.79 |  2,3,5-Trimethyl-6-propylpyrazine | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 83.92 | 0.00 |
| 37.34 |  Octanoic acid, ethyl ester | 0.86 | 0.70 | 4.74 | 0.67 | 0.34 | 1.59 | 0.06 | 1.22 |
| 40.89 |  2-(2-Methylpropyl)-3,5,6-trimethylpyrazine | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.05 | 0.00 |
| 42.91 |  2-(2-Methylpropyl)-3-(1-methylethyl)pyrazine | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 3.56 | 0.00 |
| 44.04 |  2-Undecanone | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.13 | 0.00 |
| 50.90 |  Decanoic acid, ethyl ester | 0.48 | 0.34 | 4.34 | 0.36 | 0.17 | 0.93 | 0.02 | 0.56 |
| 61.73 |  Benzeneacetaldehyde, α-(2-methylpropylidene)- | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.08 | 0.00 |
| 66.58 |  1-Hexen-3-one, 5-methyl-1-phenyl- | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.11 | 0.00 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Polito, G.; Semenzato, G.; Del Duca, S.; Castronovo, L.M.; Vassallo, A.; Chioccioli, S.; Borsetti, D.; Calabretta, V.; Puglia, A.M.; Fani, R.; et al. Endophytic Bacteria and Essential Oil from Origanum vulgare ssp. vulgare Share Some VOCs with an Antibacterial Activity. Microorganisms 2022, 10, 1424. https://doi.org/10.3390/microorganisms10071424
Polito G, Semenzato G, Del Duca S, Castronovo LM, Vassallo A, Chioccioli S, Borsetti D, Calabretta V, Puglia AM, Fani R, et al. Endophytic Bacteria and Essential Oil from Origanum vulgare ssp. vulgare Share Some VOCs with an Antibacterial Activity. Microorganisms. 2022; 10(7):1424. https://doi.org/10.3390/microorganisms10071424
Chicago/Turabian StylePolito, Giulia, Giulia Semenzato, Sara Del Duca, Lara Mitia Castronovo, Alberto Vassallo, Sofia Chioccioli, Duccio Borsetti, Vittoria Calabretta, Anna Maria Puglia, Renato Fani, and et al. 2022. "Endophytic Bacteria and Essential Oil from Origanum vulgare ssp. vulgare Share Some VOCs with an Antibacterial Activity" Microorganisms 10, no. 7: 1424. https://doi.org/10.3390/microorganisms10071424