
Control of Maize Sheath Blight and Elicit Induced Systemic Resistance Using Paenibacillus polymyxa Strain SF05

Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation of Antagonistic Bacteria from Maize
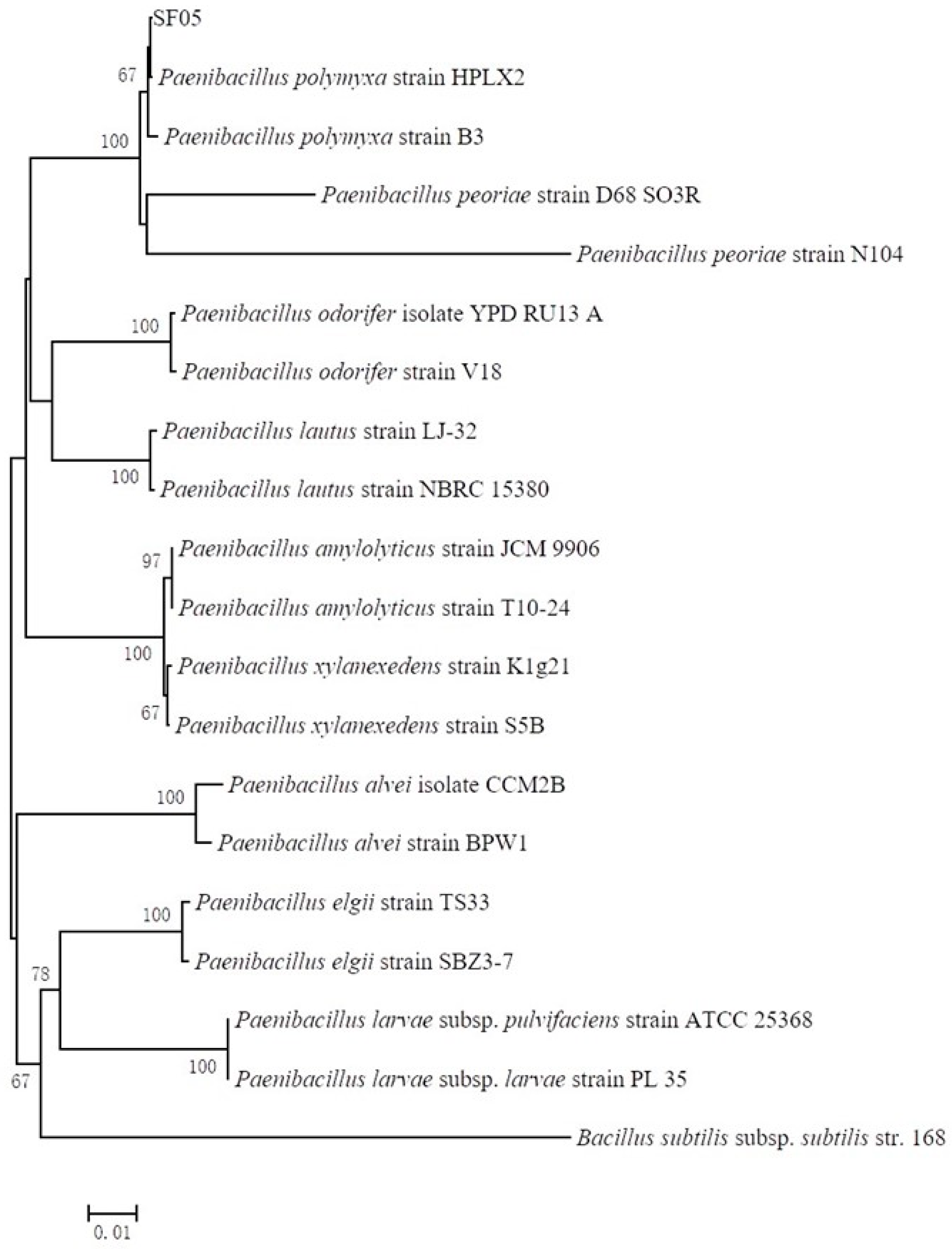
2.2. Identification of Antagonistic Bacteria SF05
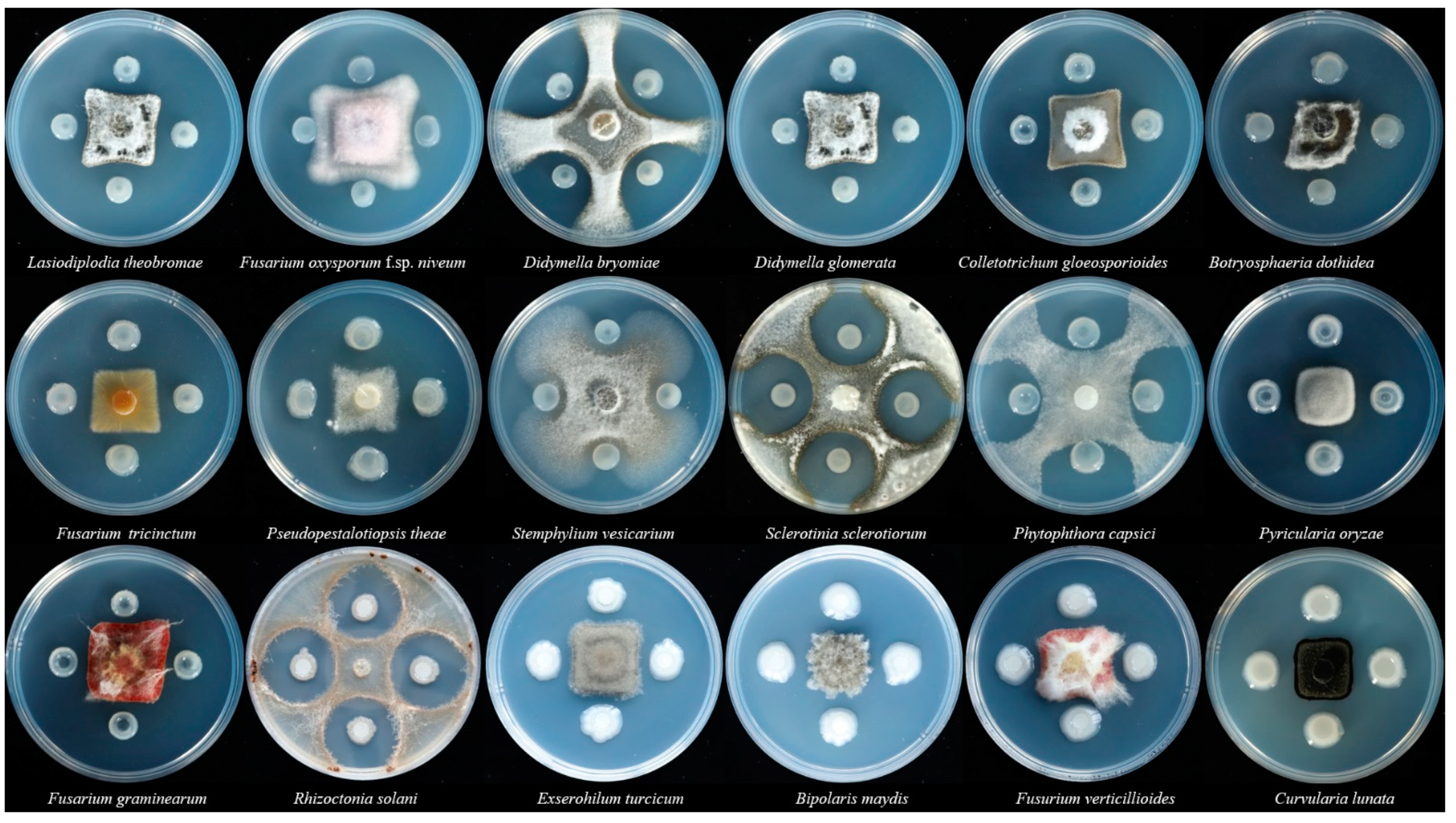
2.3. Antimicrobial Activity of Antagonistic Bacteria Strain SF05 to Pathogenic Fungi
2.4. Control Effect of P. polymyxa Strain SF05 on Banded Leaf and Sheath Blight in Field
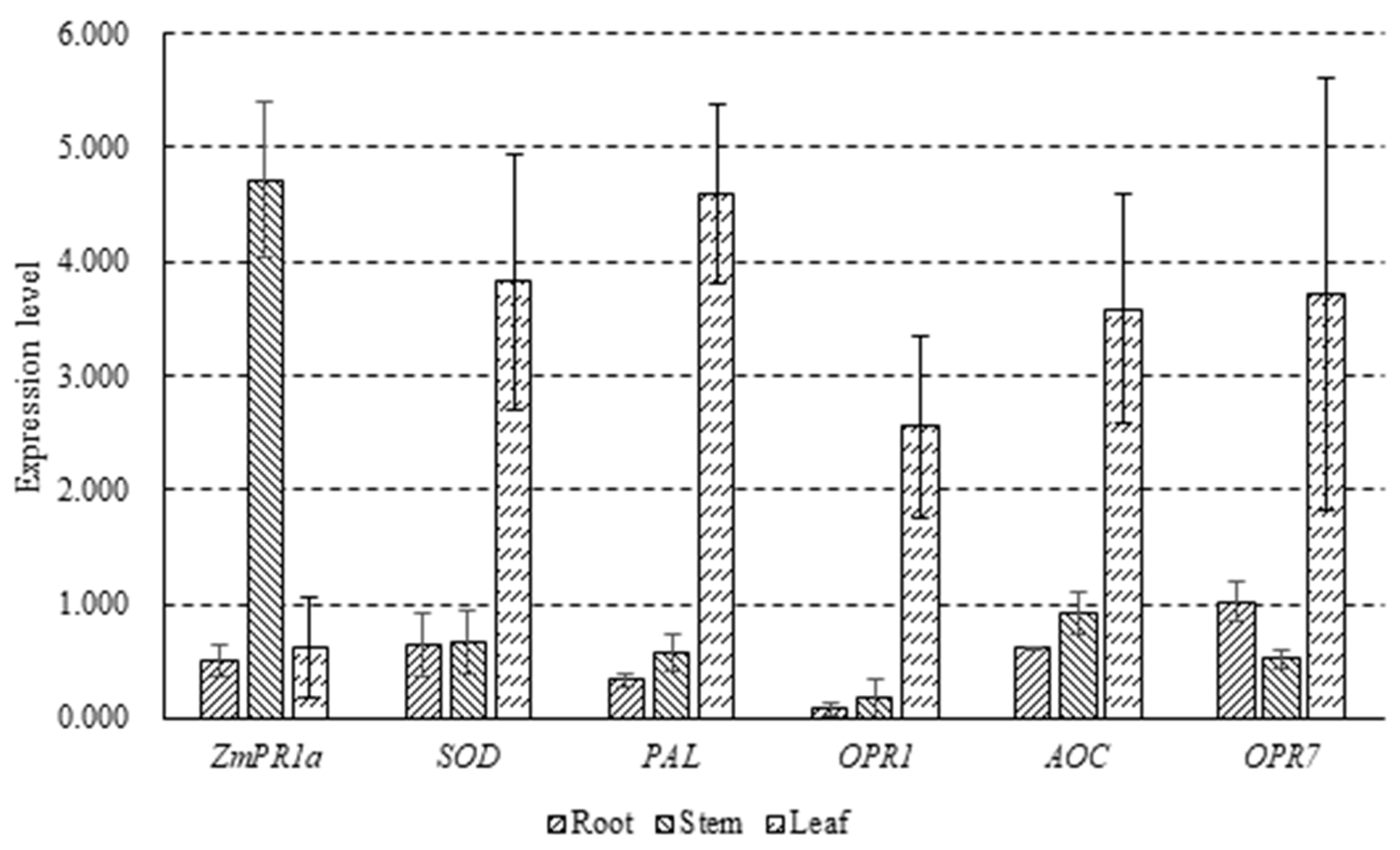
2.5. Detection of Expression Levels of Resistant Gene Induced by P. polymyxa Strain SF05 in Maize Seedling
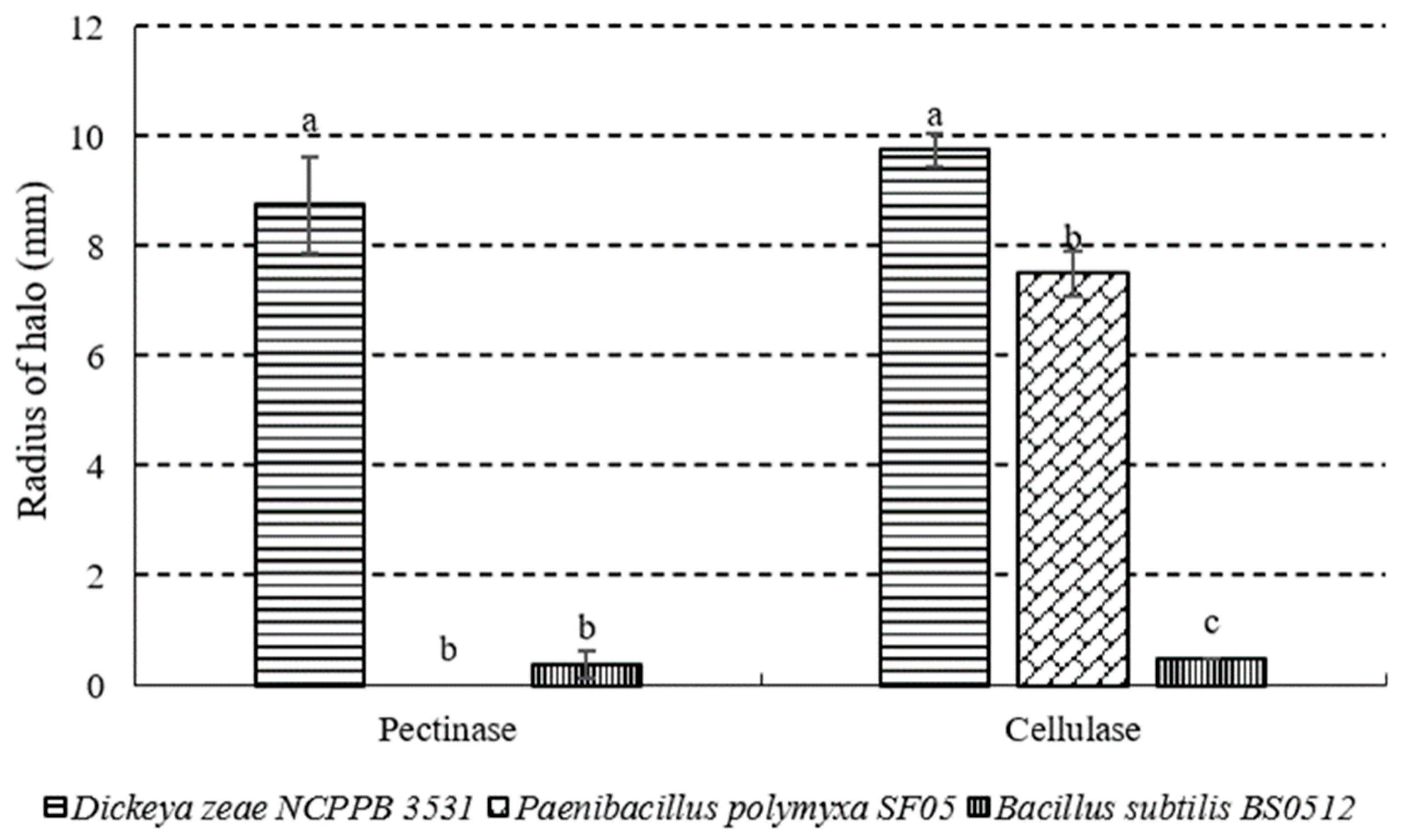
2.6. Cell Wall Degradation Enzyme Activities of P. polymyxa Strain SF05
2.7. Genome Sequencing of P. polymyxa Strain SF05
2.8. Prediction of Genes Involving Resistance Inducing Component Biosynthesis of P. polymyxa Strain SF05
2.9. Statistical Analysis
3. Results
3.1. Isolate and Identification of Antagonist Bacterium Strain SF05
3.2. Antimicrobial Activity of P. polymyxa Strain SF05 to Pathogenic Fungi
3.3. Control Effect of P. polymyxa Strain SF05 on Banded Leaf and Sheath Blight in Field
3.4. Expression Levels of Resistant Gene Induced by P. polymyxa Strain SF05 in Maize Seedling Differed in Stem and Leaf
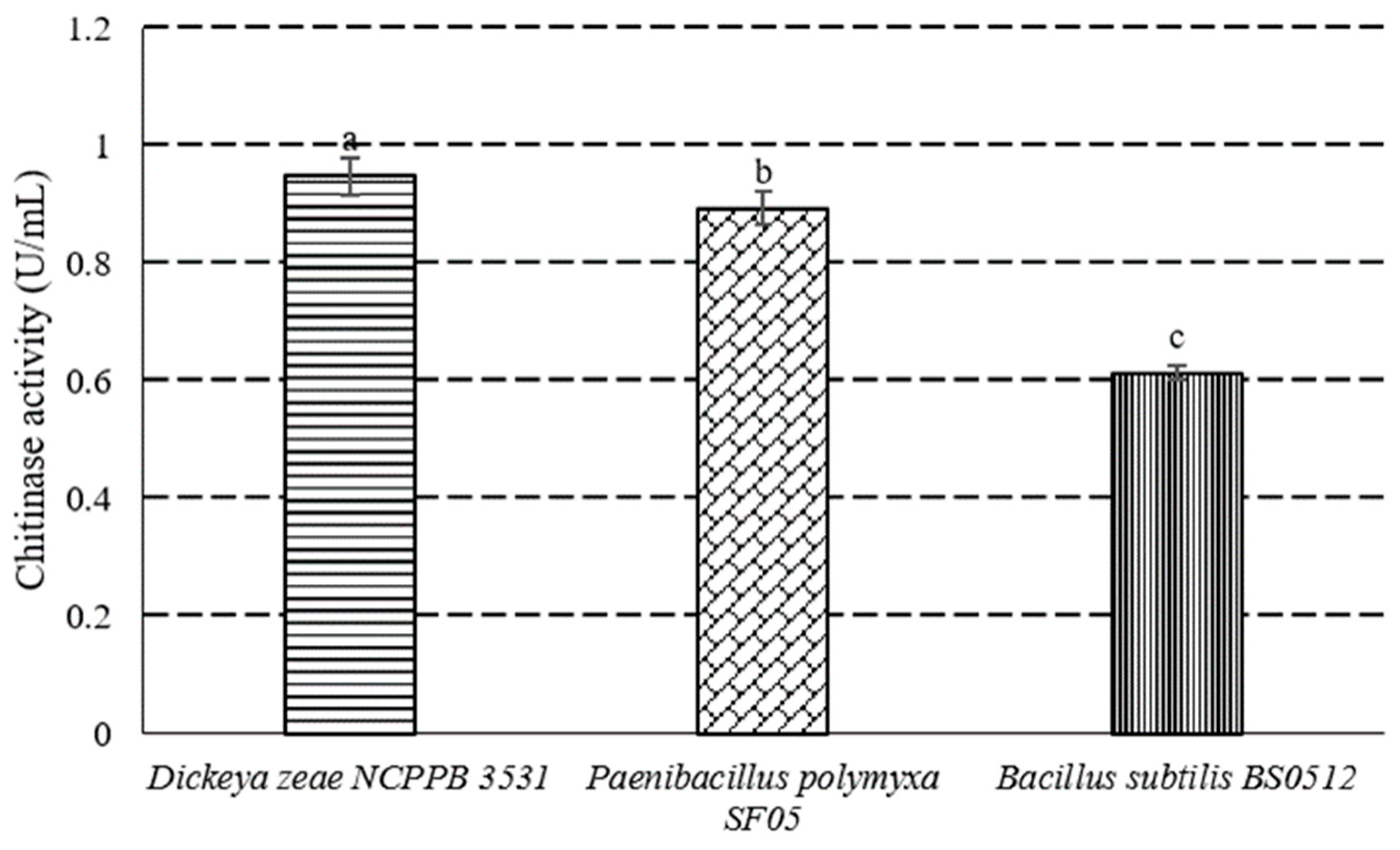
3.5. Cell Wall Degradation Enzyme Activities of P. polymyxa Strain SF05
3.6. Genome Information of P. polymyxa Strain SF05
3.7. Annotation Information of Biocontrol Mechanism of P. polymyxa Strain SF05
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Egal, S.; Hounsa, A.; Gong, Y.Y.; Turner, P.C.; Wild, C.P.; Hall, A.J.; Hell, K.; Cardwell, K.F. Dietary exposure to aflatoxin from maize and groundnut in young children from Benin and Togo, West Africa. Int. J. Food Microbiol. 2005, 104, 215–224. [Google Scholar] [CrossRef]
- Han, X.; Zhao, H.; Ren, W.; Lv, C.; Chen, C. Resistance risk assessment for fludioxonil in Bipolaris maydis. Pestic. Biochem. Physiol. 2017, 139, 32–39. [Google Scholar] [CrossRef] [PubMed]
- Da Costa, D.I.; Boller, W. Aerial and ground applications of fungicide for the control of leaf diseases in maize crop (Zea mays L.). In Proceedings of the CIGR–International Conference of Agricultural Engineering, Foz do Iguaçu, Brazil, 31 August–4 September 2008. [Google Scholar]
- Wang, G.; Han, H.; Zhao, F.; Tan, H.; Bao, F.; Lou, X. Occurrence and control of sheath blight of sweet and waxy corn in Zhejiang Province. China Plant Prot. 2016, 36, 32–36. [Google Scholar]
- Liu, H. Pathogen Identification, Pathogenic Factors and Control of Spring Maize Ear Rot disease in The Northern Region. Master s Thesis, Northeast Agricultural University, Harbin, China, 2018. [Google Scholar]
- Carvalho, F.P. Pesticides, environment, and food safety. Food Energy Secur. 2017, 6, 48–60. [Google Scholar] [CrossRef]
- Ryu, C.-M.; Murphy, J.F.; Mysore, K.S.; Kloepper, J.W. Plant growth-promoting rhizobacteria systemically protect Arabidopsis thaliana against Cucumber mosaic virus by a salicylic acid and NPR1-independent and jasmonic acid-dependent signaling pathway. Plant J. 2004, 39, 381–392. [Google Scholar] [CrossRef] [PubMed]
- Ryu, C.-M.; Kim, J.; Choi, O.; Kim, S.H.; Park, C.S. Improvement of biological control capacity of Paenibacillus polymyxa E681 by seed pelleting on sesame. Biol. Control 2006, 39, 282–289. [Google Scholar] [CrossRef]
- Son, S.H.; Khan, Z.; Kim, S.G.; Kim, Y.H. Plant growth-promoting rhizobacteria, Paenibacillus polymyxa and Paenibacillus lentimorbus suppress disease complex caused by root-knot nematode and fusarium wilt fungus. J. Appl. Microbiol. 2009, 107, 524–532. [Google Scholar] [CrossRef]
- Heulin, T.; Berge, O.; Mavingui, P.; Gouzoux, L.; Hebbar, P.; Balandreau, J. Bacillus polymyxa and Rahnella aquatilis, the dominant N2-fixing bacteria associated with wheat roots in French soils. Eur. J. Soil Biol. 1994, 30, 35–42. [Google Scholar]
- Choi, S.K.; Park, S.Y.; Kim, R.; Lee, C.H.; Kim, J.F.; Park, S.H. Identification and functional analysis of the fusaricidin biosynthetic gene of Paenibacillus polymyxa E681. Biochem. Biophys. Res. Commun. 2008, 365, 89–95. [Google Scholar] [CrossRef]
- Abdallah, Y.; Yang, M.; Zhang, M.; Masum, M.M.I.; Ogunyemi, S.O.; Hossain, A.; An, Q.; Yan, C.; Li, B. Plant growth promotion and suppression of bacterial leaf blight in rice by Paenibacillus polymyxa Sx3. Lett. Appl. Microbiol. 2019, 68, 423–429. [Google Scholar] [CrossRef]
- Raza, W.; Yang, W.; Shen, Q. Paenibacillus polymyxa: Antibiotics, hydrolytic enzymes and hazard assessment. J. Plant. Pathol. 2008, 90, 419–430. [Google Scholar]
- Timmusk, S.; Wagner, E.G. The plant-growth-promoting rhizobacterium Paenibacillus polymyxa induces changes in Arabidopsis thaliana gene expression: A possible connection between biotic and abiotic stress responses. Mol. Plant Microbe Interact. 1999, 12, 951–959. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tong, Y.; Guo, G.; Xu, J.; Ji, Z.; Chen, X. Mechanisms of resistance to gray mould in tomato plant induced by antagonistic bacteria. Acta Phytopathol. Sin. 2004, 34, 507–511. [Google Scholar]
- Polz, M.F.; Cavanaugh, C.M. Bias in template-to-product ratios in multitemplate PCR. Appl. Environ. Microbiol. 1998, 64, 3724–3730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, N. Transcriptomic Analysis of the Functions of BR, JA, and SA Involved in Maize Chlorotic Mottle Virus Infection; Chinese Academy of Agricultural Sciences: Beijing, China, 2019. [Google Scholar]
- Fan, L.; Fu, K.; Yu, C.; Li, Y.; Li, Y.; Chen, J. Thc6 protein, isolated from Trichoderma harzianum, can induce maize defense response against Curvularia Lunata. J. Basic Microbiol. 2015, 55, 591–600. [Google Scholar] [CrossRef]
- Wang, S.-q.; Ma, J.; Wang, M.; Wang, X.-h.; Li, Y.-Q.; Chen, J. Combined application of Trichoderma harzianum SH2303 and difenoconazole-propiconazolein controlling Southern corn leaf blight disease caused by Cochliobolus heterostrophus in maize. J. Integr. Agric. 2019, 18, 2063–2071. [Google Scholar] [CrossRef]
- Chen, B.; Tian, Y.; Zhao, Y.; Wang, J.; Xu, Z.; Li, X.; Hu, B. Bleeding canker of pears caused by Dickeya fangzhongdai: Symptoms, etiology and biology. J. Integr. Agric. 2020, 19, 889–897. [Google Scholar] [CrossRef]
- Goldenberger, D.; Perschil, I.; Ritzler, M.; Altwegg, M. A Simple Universal DNA Extraction Procedure Using Sds and Proteinase-K Is Compatible with Direct Pcr Amplification. Pcr. Meth. Appl. 1995, 4, 368–370. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Wu, J.; Yang, J.; Sun, S.; Xiao, J.; Yu, J. PGAP: Pan-genomes analysis pipeline. Bioinformatics 2012, 28, 416–418. [Google Scholar] [CrossRef] [Green Version]
- Shi, Y.; Niu, K.; Huang, B.; Liu, W.; Ma, H. Transcriptional Responses of Creeping Bentgrass to 2,3-Butanediol, a Bacterial Volatile Compound (BVC) Analogue. Molecules 2017, 22, 1318. [Google Scholar] [CrossRef] [Green Version]
- Lee, B.; Farag, M.A.; Park, H.B.; Kloepper, J.W.; Lee, S.H.; Ryu, C.M. Induced resistance by a long-chain bacterial volatile: Elicitation of plant systemic defense by a C13 volatile produced by Paenibacillus polymyxa. PLoS ONE 2012, 7, e48744. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.; Cai, M. Handbook of Systematic for Common Bacteria; Science Press: Beijing, China, 2001. [Google Scholar]
- Saravanan, R.; Nakkeeran, S.; Saranya, N.; Senthilraja, C.; Renukadevi, P.; Krishnamoorthy, A.S.; El Enshasy, H.A.; El-Adawi, H.; Malathi, V.G.; Salmen, S.H.; et al. Mining the Genome of Bacillus velezensis VB7 (CP047587) for MAMP Genes and Non-Ribosomal Peptide Synthetase Gene Clusters Conferring Antiviral and Antifungal Activity. Microorganisms 2021, 9, 2511. [Google Scholar]
- Hong, S.R.; Kim, T.Y.; Won, S.J.; Moon, J.H.; Ajuna, H.B.; Kim, K.Y.; Ahn, Y.S. Control of Fungal Diseases and Fruit Yield Improvement of Strawberry Using Bacillus velezensis CE 100. Microorganisms 2022, 10, 365. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Liang, L.; Shao, H.; Ye, X.; Yang, X.; Chen, X.; Shi, Y.; Zhang, L.; Xu, L.; Wang, J. Isolation of the Novel Strain Bacillus amyloliquefaciens F9 and Identification of Lipopeptide Extract Components Responsible for Activity against Xanthomonas citri subsp. citri. Plants 2022, 11, 457. [Google Scholar] [CrossRef] [PubMed]
- Hong, P.; Hao, W.N.; Luo, J.J.; Chen, S.H.; Hu, M.Y.; Zhong, G.H. Combination of hot water, Bacillus amyloliquefaciens HF-01 and sodium bicarbonate treatments to control postharvest decay of mandarin fruit. Postharvest Biol. Technol. 2014, 88, 96–102. [Google Scholar] [CrossRef]
- Lai, K.P.; Chen, S.H.; Hu, M.Y.; Hu, Q.B.; Geng, P.; Weng, Q.F.; Jia, J.W. Control of postharvest green mold of citrus fruit by application of endophytic Paenibacillus polymyxa strain SG-6. Postharvest Biol. Technol. 2012, 69, 40–48. [Google Scholar] [CrossRef]
- Hong, C.E.; Kwon, S.Y.; Park, J.M. Biocontrol activity of Paenibacillus polymyxa AC-1 against Pseudomonas syringae and its interaction with Arabidopsis thaliana. Microbiol. Res. 2016, 185, 13–21. [Google Scholar] [CrossRef]
- Timmusk, S. Biofilm forming Paenibacillus polymyxa antagonizes oomycete plant pathogens Phytophthora palmivora and Pythium aphanidermatum. J. Biotechnol. 2008, 136, S265–S266. [Google Scholar] [CrossRef]
- Haggag, W.M.; Timmusk, S. Colonization of peanut roots by biofilm-forming Paenibacillus polymyxa initiates biocontrol against crown rot disease. J. Appl. Microbiol. 2008, 104, 961–969. [Google Scholar] [CrossRef]
- McAuliffe, O.; Ross, R.P.; Hill, C. Lantibiotics: Structure, biosynthesis and mode of action. FEMS Microbiol. Rev. 2001, 25, 285–308. [Google Scholar] [CrossRef] [Green Version]
- Piuri, M.; Sanchez-Rivas, C.; Ruzal, S.M. A novel antimicrobial activity of a Paenibacillus polymyxa strain isolated from regional fermented sausages. Lett. Appl. Microbiol. 1998, 27, 9–13. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.; Han, H.; Hou, J.; Wang, G.; Zhao, F. Rapid screening of maize cultivars with resistance to banded leaf and sheath blight by ZmPR1a gene expression level. Mol. Plant Breed. 2021, 19, 7467–7472. [Google Scholar]
- Costerton, J.W.; Stewart, P.S.; Greenberg, E.P. Bacterial biofilms: A common cause of persistent infections. Science 1999, 284, 1318–1322. [Google Scholar] [CrossRef] [Green Version]
- Timmusk, S.; Grantcharova, N.; Wagner, E.G. Paenibacillus polymyxa invades plant roots and forms biofilms. Appl. Environ. Microbiol. 2005, 71, 7292–7300. [Google Scholar] [CrossRef] [Green Version]
- Hao, T.; Chen, S. Colonization of Wheat, Maize and Cucumber by Paenibacillus polymyxa WLY78. PLoS ONE 2017, 12, e0169980. [Google Scholar] [CrossRef]
- Pandey, K.D.; Patel, A.K.; Singh, M.; Vandana; Kumari, A.; Jalaluddin. Chapter 2—Secondary metabolites from bacteria and viruses. In Natural Bioactive Compounds; Sinha, R.P., Häder, D.-P., Eds.; Academic Press: Cambridge, MA, USA, 2021; pp. 19–40. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Source |
|---|---|
| Rhizoctonia solani | Laboratory Collection |
| Exserohilum turcicum | Laboratory Collection |
| Bipolaris maydis | Laboratory Collection |
| Fusurium verticillioides | Laboratory Collection |
| Sclerotinia sclerotiorum | Laboratory Collection |
| Curvularia lunata | Hebei Academy of Agricultural and Forestry Sciences |
| F. graminearum | Nanjing Agricultural University |
| Lasiodiplodia theobromae | Nanjing Agricultural University |
| F. oxysporum f.sp. niveum | Nanjing Agricultural University |
| Didymella bryoniae | Nanjing Agricultural University |
| D. glomerata | Nanjing Agricultural University |
| Colletotrichum gloeosporioides | Nanjing Agricultural University |
| Botryosphaeria dothidea | Nanjing Agricultural University |
| F. tricinctum | Nanjing Agricultural University |
| Pseudopestalotiopsis theae | Nanjing Agricultural University |
| Stemphylium vesicarium | Nanjing Agricultural University |
| Phytophthora capsici | Nanjing Agricultural University |
| Pyricularia oryzae | Nanjing Agricultural University |
| Primer | Sequences (5′-3′) | References |
|---|---|---|
| Actin-F | GGTTCTATTCCAGCCATCCTTCATTG | [12] |
| Actin-R | TCTCCTTGCTCATGCGGTCAC | |
| ZmPR1a-F | GGCGAGAGCCCCTACTAGAC | [13] |
| ZmPR1a-R | AAATCGCCTGCATGGTTTTA | |
| PAL-F | GAAGCTCATGTCGTCCACCTA | [12] |
| PAL-R | GTTCATGGTCAGCACCTTCTT | |
| SOD-F | AGAATAACATCCCGAAGACATC | [12] |
| SOD-R | AGCCAACAGTCCAACACAGT | |
| OPR1-F | CGTATGGGAGGCTGTTCTTG | [12] |
| OPR1-R | AGCGGTCGTATTTGTTGAGTG | |
| OPR7-F | GAGAAAGGTGGTTGATGCTGTT | [12] |
| OPR7-R | GGAGTTGGATACTTGCCATAGG | |
| AOC-F | GGGCATCTGCGTGCTCATC | [14] |
| AOC-R | ACCGCCAGGTACGACTCCTC |
| Treatment | Disease Index | Control Effect |
|---|---|---|
| Biological Treatment | 37.38 ± 1.23 b | 37.06% |
| Chemical Treatment | 32.20 ± 0.75 a | 45.79% |
| Negative Control | 59.40 ± 2.17 c | - |
| Strain | GenBank | Size (Mb) | GC% | Protein | rRNA | tRNA | Other RNA | Gene | Pseudogene |
|---|---|---|---|---|---|---|---|---|---|
| SF05 | - | 5.46 | 45.5 | 4615 | 43 | 108 | 4 | 5018 | 248 |
| ZF129 | GCA_006274405.1 | 5.82 | 45.4 | 4873 | 42 | 111 | 4 | 5115 | 85 |
| CF05 | GCA_000785455.1 | 5.76 | 45.5 | 4738 | 43 | 107 | 4 | 4966 | 74 |
| YC0573 | GCA_001874425.3 | 6.13 | 45.6 | 5091 | 37 | 102 | 4 | 5442 | 208 |
| Sb3-1 | GCA_000819665.1 | 5.83 | 45.5 | 4812 | 46 | 109 | 4 | 5097 | 126 |
| HY96-2 | GCA_002893885.1 | 5.75 | 45.6 | 4663 | 42 | 110 | 4 | 4955 | 136 |
| CJX518 | GCA_014854715.1 | 5.69 | 45.4 | 4749 | 42 | 111 | 4 | 5034 | 128 |
| JE201 | GCA_019852195.1 | 6.17 | 45.3 | 5145 | 46 | 111 | 4 | 5425 | 119 |
| EBL06 | GCA_000955925.1 | 5.68 | 45.6 | 4821 | 7 | 38 | 4 | 4964 | 94 |
| E681 | GCA_014706575.1 | 5.42 | 45.8 | 4532 | 36 | 91 | 4 | 4872 | 209 |
| VOC | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 |
|---|---|---|---|---|---|---|---|---|---|---|
| 2,3-Butanediol | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 |
| Acetone | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 |
| Acetylene | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
| Glyoxylic acid | 1 | 0 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
| Benzaldehyde | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 1 |
| Butanol | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 |
| Ethanol | 3 | 2 | 3 | 2 | 2 | 2 | 2 | 2 | 2 | 3 |
| Methanethiol | 2 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 | 3 |
| Total | 15 | 14 | 16 | 15 | 15 | 15 | 15 | 15 | 15 | 15 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, B.; Han, H.; Hou, J.; Bao, F.; Tan, H.; Lou, X.; Wang, G.; Zhao, F. Control of Maize Sheath Blight and Elicit Induced Systemic Resistance Using Paenibacillus polymyxa Strain SF05. Microorganisms 2022, 10, 1318. https://doi.org/10.3390/microorganisms10071318
Chen B, Han H, Hou J, Bao F, Tan H, Lou X, Wang G, Zhao F. Control of Maize Sheath Blight and Elicit Induced Systemic Resistance Using Paenibacillus polymyxa Strain SF05. Microorganisms. 2022; 10(7):1318. https://doi.org/10.3390/microorganisms10071318
Chicago/Turabian StyleChen, Bin, Hailiang Han, Junfeng Hou, Fei Bao, Heping Tan, Xiaocheng Lou, Guiyue Wang, and Fucheng Zhao. 2022. "Control of Maize Sheath Blight and Elicit Induced Systemic Resistance Using Paenibacillus polymyxa Strain SF05" Microorganisms 10, no. 7: 1318. https://doi.org/10.3390/microorganisms10071318