
Unraveling Plant Cell Death during Phytophthora Infection


Abstract
:1. Introduction
2. Programed Cell Death in Host Plants
2.1. Classification of PCD
2.2. Programed Cell Death in Host Plants
2.2.1. Transcriptional Regulation of pPCD
2.2.2. Phytohormone Signaling Pathways Involved in pPCD
2.2.3. Triggers of pPCD
3. Cell Death and Phytophthora Virulence
4. Phytophthora Effectors That Induce or Suppress Cell Death
4.1. Apoplastic Effectors
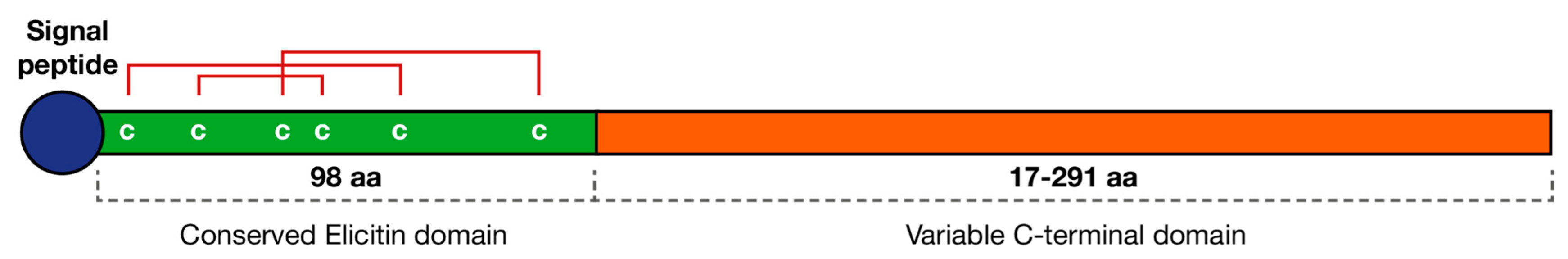
4.1.1. Elicitins
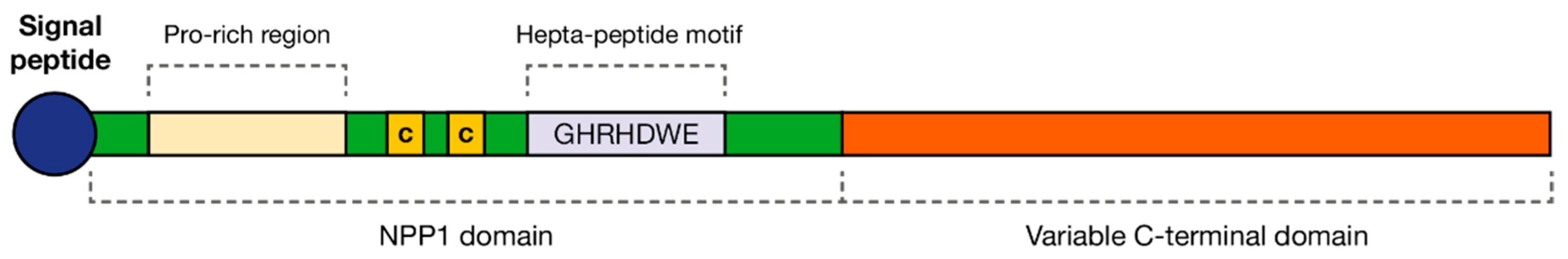
4.1.2. Nep1-like Protein (NLPs)
4.2. Cytoplasmic Effectors
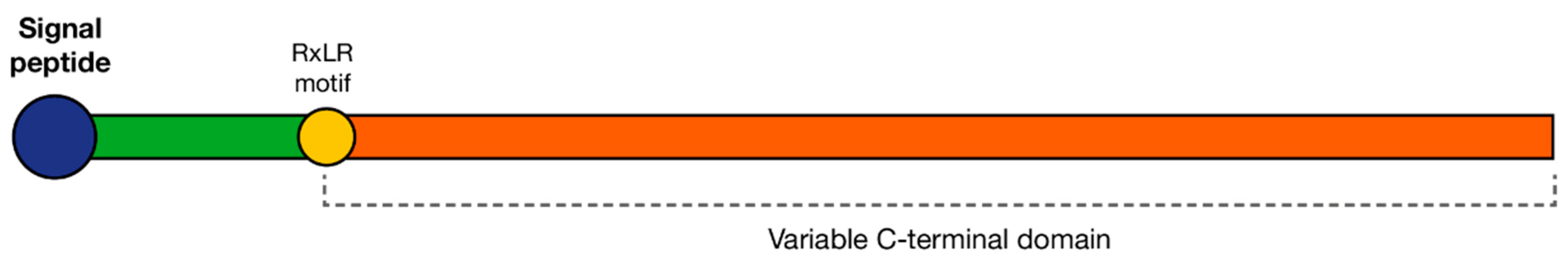
4.2.1. RxLRs
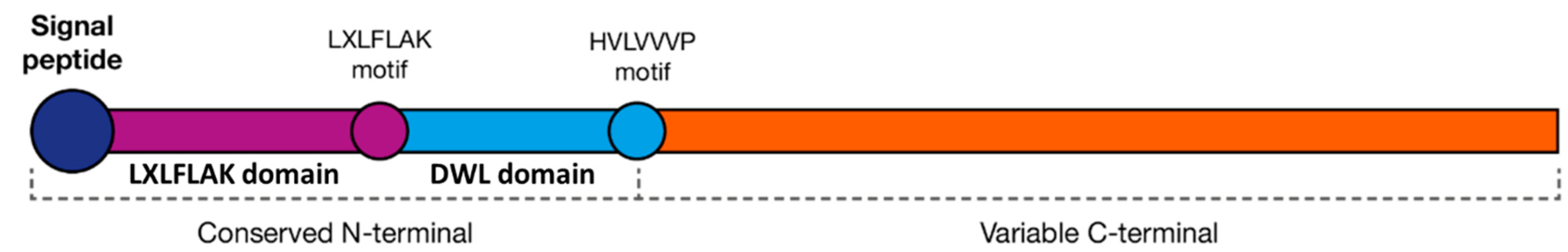
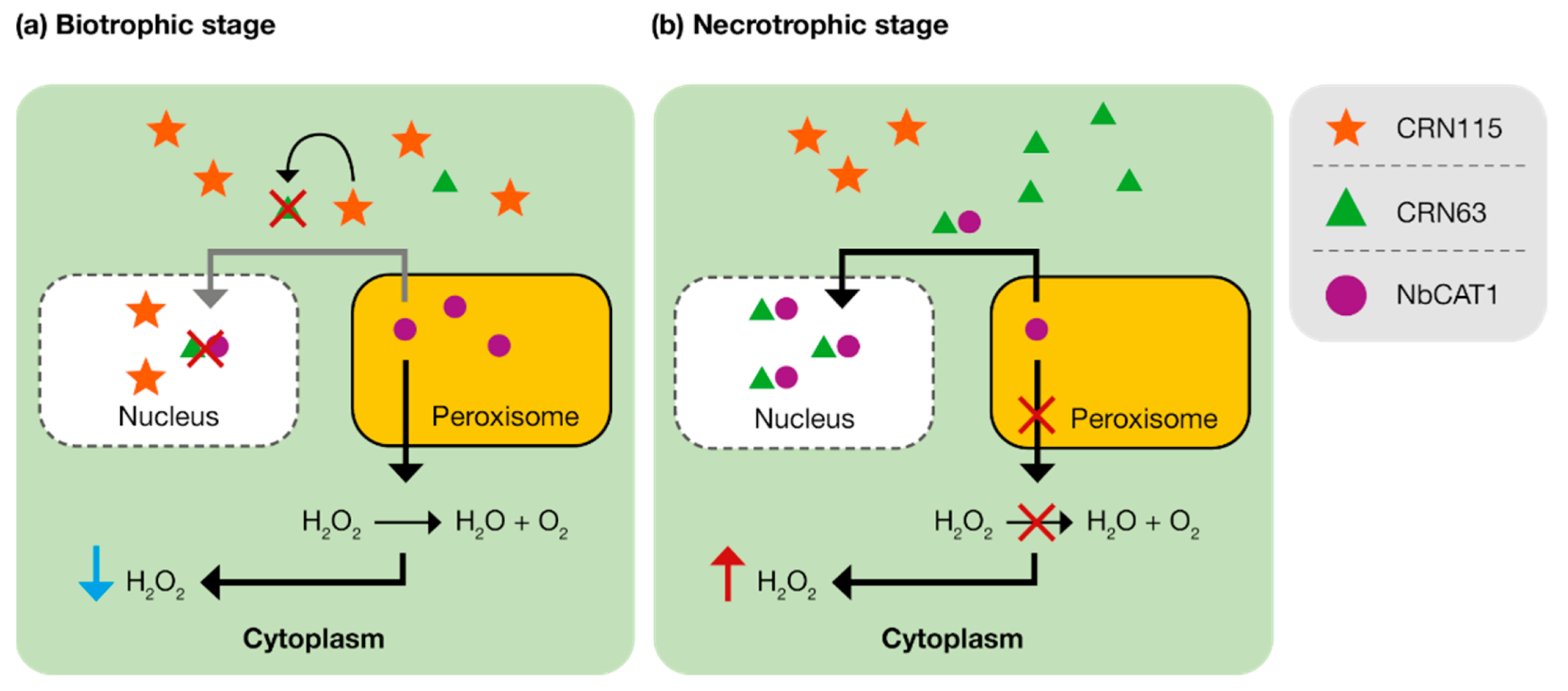
4.2.2. Crinklers
5. Techniques Used in the Functional Characterization of Phytophthora Effectors
6. Conclusions
Funding
Conflicts of Interest
References
- Kroon, L.P.N.M.; Brouwer, H.; De Cock, A.W.A.M.; Govers, F. The Phytophthora genus anno 2012. Phytopathology 2012, 102, 348–364. [Google Scholar] [CrossRef] [Green Version]
- Lamour, K.H.; Stam, R.; Jupe, J.; Huitema, E. The oomycete broad-host-range pathogen Phytophthora capsici. Mol. Plant Pathol. 2012, 13, 329–337. [Google Scholar] [CrossRef] [PubMed]
- Kamoun, S.; Furzer, O.; Jones, J.D.G.; Judelson, H.S.; Ali, G.S.; Dalio, R.J.D.; Roy, S.G.; Schena, L.; Zambounis, A.; Panabières, F.; et al. The Top 10 oomycete pathogens in molecular plant pathology. Mol. Plant Pathol. 2015, 16, 413–434. [Google Scholar] [CrossRef] [PubMed]
- Boevink, P.C.; Birch, P.R.J.; Turnbull, D.; Whisson, S.C. Devastating intimacy: The cell biology of plant—Phytophthora interactions. New Phytol. 2020, 228, 445–458. [Google Scholar] [CrossRef] [PubMed]
- Kinealy, C. This Great Calamity: The Irish Famine 1845-52; Gill & Macmillan Ltd.: Dublin, Ireland, 2006; pp. 10–98. [Google Scholar]
- Batini, F.E.; Hopkins, E.R. Phytophthora cinnamomi Rands—a root pathogen of the Jarrah Forest. Aust. For. 1972, 36, 57–68. [Google Scholar] [CrossRef]
- Rani, G.D. Advances in Soil Borne Plant Diseases; New India Publishing: New Dehli, India, 2008; pp. 371–413. [Google Scholar]
- Mafurah, J.J.; Ma, H.; Zhang, M.; Xu, J.; He, F.; Ye, T.; Shen, D.; Chen, Y.; Rajput, N.A.; Dou, D. A virulence essential CRN effector of Phytophthora capsici suppresses host defense and induces cell death in plant nucleus. PLoS ONE 2015, 10, e0127965. [Google Scholar] [CrossRef]
- Huysmans, M.; Coll, N.S.; Nowack, M.K. Dying two deaths—programmed cell death regulation in development and disease. Curr. Opin. Plant Biol. 2017, 35, 37–44. [Google Scholar] [CrossRef] [Green Version]
- Tyler, B.M.; Tripathy, S.; Zhang, X.; Dehal, P.; Jiang, R.H.Y.; Aerts, A.; Arredondo, F.D.; Baxter, L.; Bensasson, D.; Beynon, J.L.; et al. Phytophthora genome sequences uncover evolutionary origins and mechanisms of pathogenesis. Science 2006, 313, 1261–1266. [Google Scholar] [CrossRef] [Green Version]
- Engelbrecht, J.; Duong, T.A.; Prabhu, S.A.; Seedat, M.; Berg, N.V.D. Genome of the destructive oomycete Phytophthora cinnamomi provides insights into its pathogenicity and adaptive potential. BMC Genom. 2021, 22, 302. [Google Scholar] [CrossRef]
- Hardham, A.R. Phytophthora cinnamomi. Mol. Plant Pathol. 2005, 6, 589–604. [Google Scholar] [CrossRef]
- Koeck, M.; Hardham, A.R.; Dodds, P.N. The role of effectors of biotrophic and hemibiotrophic fungi in infection. Cell. Microbiol. 2011, 13, 1849–1857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Doorn, W.G.; Beers, E.P.; Dangl, J.L.; E Franklin-Tong, V.; Gallois, P.; Hara-Nishimura, I.; Jones, A.M.; Kawai-Yamada, M.; Lam, E.; Mundy, J.; et al. Morphological classification of plant cell deaths. Cell Death Differ. 2011, 18, 1241–1246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petrov, V.; Hille, J.; Mueller-Roeber, B.; Gechev, T.S. ROS-mediated abiotic stress-induced programmed cell death in plants. Front. Plant Sci. 2015, 6, 69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mittler, R.; Blumwald, E. Genetic engineering for modern agriculture: Challenges and perspectives. Annu. Rev. Plant Biol. 2010, 61, 443–462. [Google Scholar] [CrossRef] [Green Version]
- Burke, R.; Schwarze, J.; Sherwood, O.L.; Jnaid, Y.; McCabe, P.F.; Kacprzyk, J. Stressed to death: The role of transcription factors in plant programmed cell death induced by abiotic and biotic stimuli. Front. Plant Sci. 2020, 11, 1235. [Google Scholar] [CrossRef] [PubMed]
- Kiraly, Z.; Barna, B.; Ersek, T. Hypersensitivity as a consequence, not the cause, of plant resistance to infection. Nature 1972, 239, 456–458. [Google Scholar] [CrossRef]
- Hatsugai, N.; Kuroyanagi, M.; Yamada, K.; Meshi, T.; Tsuda, S.; Kondo, M.; Nishimura, M.; Hara-Nishimura, I. A plant vacuolar protease, VPE, mediates virus-induced hypersensitive cell death. Science 2004, 305, 855–858. [Google Scholar] [CrossRef]
- Rojo, E.; Martín, R.; Carter, C.; Zouhar, J.; Pan, S.; Plotnikova, J.; Jin, H.; Paneque, M.; Serrano, J.J.S.; Baker, B.; et al. VPEγ exhibits a caspase-like activity that contributes to defense against pathogens. Curr. Biol. 2004, 14, 1897–1906. [Google Scholar] [CrossRef] [Green Version]
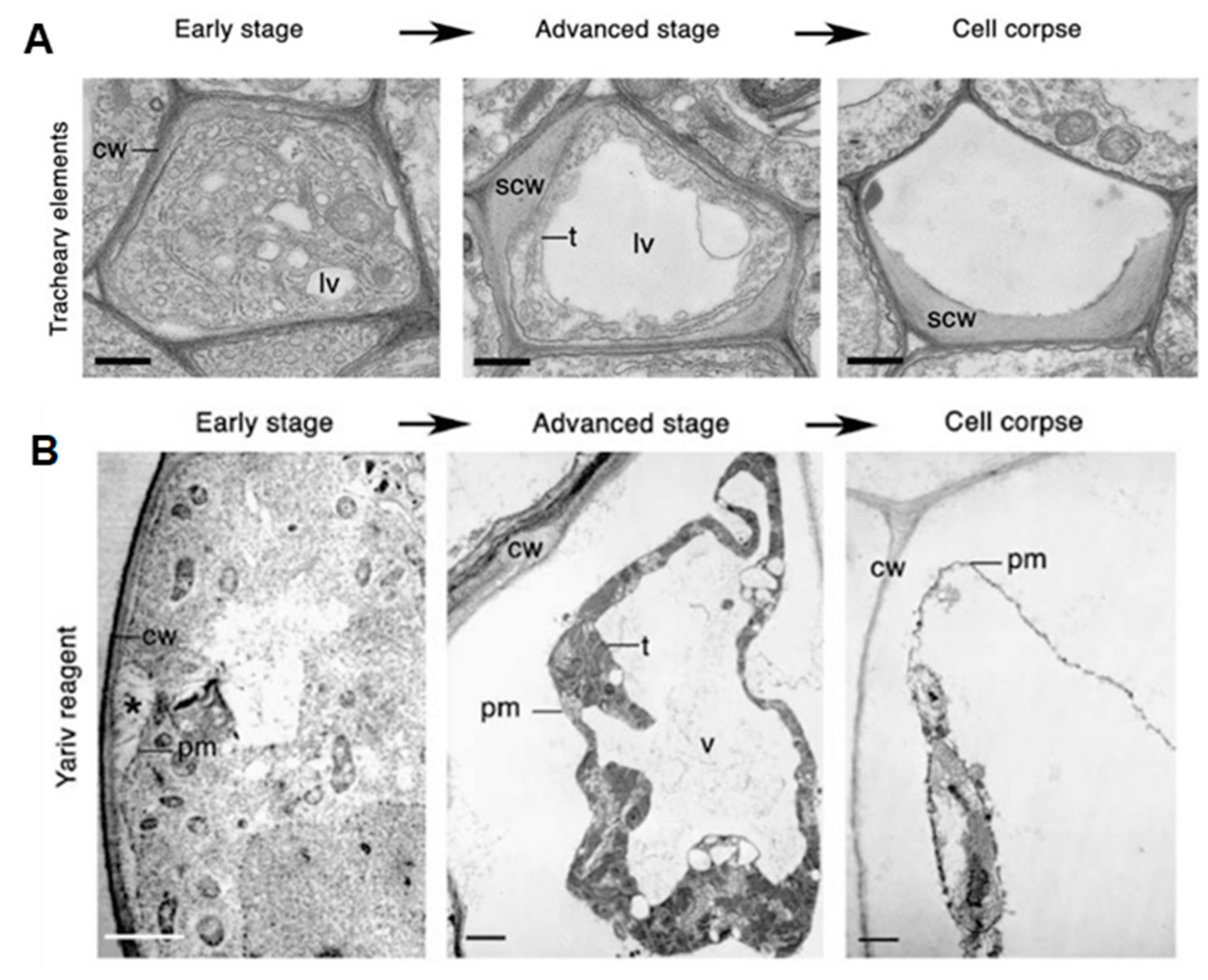
- Avci, U.; Petzold, H.E.; Ismail, I.O.; Beers, E.P.; Haigler, C.H. Cysteine proteases XCP1 and XCP2 aid micro-autolysis within the intact central vacuole during xylogenesis in Arabidopsis roots. Plant J. 2008, 56, 303–315. [Google Scholar] [CrossRef]
- Gao, M.; Showalter, A.M. Yariv reagent treatment induces programmed cell death in Arabidopsis cell cultures and implicates arabinogalactan protein involvement. Plant J. 1999, 19, 321–331. [Google Scholar] [CrossRef] [Green Version]
- Mur, L.A.J.; Kenton, P.; Lloyd, A.J.; Ougham, H.; Prats, E. The hypersensitive response; the centenary is upon us but how much do we know? J. Exp. Bot. 2008, 59, 501–520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, M.; Li, Q.; Liu, T.; Liu, L.; Shen, D.; Zhu, Y.; Liu, P.; Zhou, J.-M.; Dou, D. Two cytoplasmic effectors of Phytophthora sojae regulate plant cell death via interactions with plant catalases. Plant Physiol. 2015, 167, 164–175. [Google Scholar] [CrossRef] [Green Version]
- Daneva, A.; Gao, Z.; Van Durme, M.; Nowack, M.K. Functions and regulation of programmed cell death in plant development. Annu. Rev. Cell Dev. Biol. 2016, 32, 441–468. [Google Scholar] [CrossRef] [PubMed]
- Mukhtar, M.S.; McCormack, M.E.; Argueso, C.T.; Pajerowska-Mukhtar, K.M. Pathogen tactics to manipulate plant cell death. Curr. Biol. 2016, 26, 608–619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fuchs, Y.; Steller, H. Programmed cell death in animal development and disease. Cell 2011, 147, 742–758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhai, Z.; Ha, N.; Papagiannouli, F.; Hamacher-Brady, A.; Brady, N.; Sorge, S.; Bezdan, D.; Lohmann, I. Antagonistic regulation of apoptosis and differentiation by the Cut transcription factor represents a tumor-suppressing mechanism in Drosophila. PLoS Genet. 2012, 8, e1002582. [Google Scholar] [CrossRef] [Green Version]
- Aubrey, B.J.; Kelly, G.L.; Janic, A.; Herold, M.J.; Strasser, A. How does p53 induce apoptosis and how does this relate to p53-mediated tumour suppression? Cell Death Differ. 2018, 25, 104–113. [Google Scholar] [CrossRef] [Green Version]
- Bin Nasir, K.H.; Takahashi, Y.; Ito, A.; Saitoh, H.; Matsumura, H.; Kanzaki, H.; Shimizu, T.; Ito, M.; Fujisawa, S.; Sharma, P.C.; et al. High-throughput in planta expression screening identifies a class II ethylene-responsive element binding factor-like protein that regulates plant cell death and non-host resistance. Plant J. 2005, 43, 491–505. [Google Scholar] [CrossRef]
- Xie, H.-T.; Wan, Z.-Y.; Li, S.; Zhang, Y. Spatiotemporal production of reactive oxygen species by NADPH oxidase is critical for tapetal programmed cell death and pollen development in Arabidopsis. Plant Cell 2014, 26, 2007–2023. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.H.; Jeon, H.S.; Kim, H.G.; Park, O.K. An Arabidopsis NAC transcription factor NAC4 promotes pathogen-induced cell death under negative regulation by microRNA164. New Phytol. 2017, 214, 343–360. [Google Scholar] [CrossRef]
- Awwad, F.; Bertrand, G.; Grandbois, M.; Beaudoin, N. Auxin protects Arabidopsis thaliana cell suspension cultures from programmed cell death induced by the cellulose biosynthesis inhibitors thaxtomin A and isoxaben. BMC Plant Biol. 2019, 19, 512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, X.; Wang, H.; Cai, J.; Li, D.; Song, F. NAC transcription factors in plant immunity. Phytopathol. Res. 2019, 1, 3. [Google Scholar] [CrossRef]
- Kaneda, T.; Taga, Y.; Takai, R.; Iwano, M.; Matsui, H.; Takayama, S.; Isogai, A.; Che, F.-S. The transcription factor OsNAC4 is a key positive regulator of plant hypersensitive cell death. EMBO J. 2009, 28, 926–936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, X.; Chen, C.; Fan, B.; Chen, Z. Physical and functional interactions between pathogen-induced Arabidopsis WRKY18, WRKY40, and WRKY60 transcription factors. Plant Cell 2006, 18, 1310–1326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, L.-L.; Xue, H.-W. The MADS29 transcription factor regulates the degradation of the nucellus and the nucellar projection during rice seed development. Plant Cell 2012, 24, 1049–1065. [Google Scholar] [CrossRef] [Green Version]
- Qi, T.; Wang, J.; Huang, H.; Liu, B.; Gao, H.; Liu, Y.; Song, S.; Xie, D. Regulation of jasmonate-induced leaf senescence by antagonism between bHLH subgroup IIIe and IIId factors in Arabidopsis. Plant Cell 2015, 27, 1634–1649. [Google Scholar] [CrossRef] [Green Version]
- Ueda, H.; Kusaba, M. Strigolactone regulates leaf senescence in concert with ethylene in Arabidopsis. Plant Physiol. 2015, 169, 138–147. [Google Scholar] [CrossRef] [Green Version]
- Glazebrook, J. Contrasting mechanisms of defense against biotrophic and necrotrophic pathogens. Annu. Rev. Phytopathol. 2005, 43, 205–227. [Google Scholar] [CrossRef]
- Huang, S.; Zhang, X.; Fernando, W.G.D. Directing trophic divergence in plant-pathogen interactions: Antagonistic phytohormones with no doubt? Front. Plant Sci. 2020, 11, 600063. [Google Scholar] [CrossRef]
- Pieterse, C.M.J.; Leon-Reyes, A.; Van der Ent, S.; Van Wees, S.C.M. Networking by small-molecule hormones in plant immunity. Nat. Chem. Biol. 2009, 5, 308–316. [Google Scholar] [CrossRef] [Green Version]
- Birkenbihl, R.P.; Somssich, I.E. Transcriptional plant responses critical for resistance towards necrotrophic pathogens. Front. Plant Sci. 2011, 2, 76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kazan, K.; Lyons, R. Intervention of phytohormone pathways by pathogen effectors. Plant Cell 2014, 26, 2285–2309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Durme, M.; Nowack, M.K. Mechanisms of developmentally controlled cell death in plants. Curr. Opin. Plant Biol. 2016, 29, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Wilkins, K.A.; Bosch, M.; Haque, T.; Teng, N.; Poulter, N.S.; Franklin-Tong, V.E. Self-incompatibility-induced programmed cell death in field poppy pollen involves dramatic acidification of the incompatible pollen tube cytosol. Plant Physiol. 2015, 167, 766–779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coll, N.S.; Epple, P.; Dangl, J.L. Programmed cell death in the plant immune system. Cell Death Differ. 2011, 18, 1247–1256. [Google Scholar] [CrossRef] [Green Version]
- Herrera-Vãsquez, A.; Salinas, P.; Holuigue, L. Salicylic acid and reactive oxygen species interplay in the transcriptional control of defense genes expression. Front. Plant Sci. 2015, 6, 171. [Google Scholar] [CrossRef] [Green Version]
- Lorang, J.; Kidarsa, T.; Bradford, C.S.; Gilbert, B.; Curtis, M.; Tzeng, S.-C.; Maier, C.S.; Wolpert, T.J. Tricking the guard: Exploiting plant defense for disease susceptibility. Science 2012, 338, 659–662. [Google Scholar] [CrossRef] [Green Version]
- Lorang, J. Necrotrophic exploitation and subversion of plant defense: A lifestyle or just a phase, and implications in breeding resistance. Phytopathology 2019, 109, 332–346. [Google Scholar] [CrossRef] [Green Version]
- Stam, R.; Jupe, J.; Howden, A.J.M.; Morris, J.A.; Boevink, P.C.; Hedley, P.E.; Huitema, E. Identification and characterisation CRN effectors in Phytophthora capsici shows modularity and functional diversity. PLoS ONE 2013, 8, e59517. [Google Scholar] [CrossRef]
- Münch, S.; Lingner, U.; Floss, D.S.; Ludwig, N.; Sauer, N.; Deising, H.B. The hemibiotrophic lifestyle of Colletotrichum species. J. Plant Physiol. 2008, 165, 41–51. [Google Scholar] [CrossRef] [PubMed]
- Jupe, J.; Stam, R.; Howden, A.J.; Morris, J.A.; Zhang, R.; Hedley, P.E.; Huitema, E. Phytophthora capsici-tomato interaction features dramatic shifts in gene expression associated with a hemi-biotrophic lifestyle. Genome Biol. 2013, 14, R63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, T.; Ye, W.; Ru, Y.; Yang, X.; Gu, B.; Tao, K.; Lu, S.; Dong, S.; Zheng, X.; Shan, W.; et al. Two host cytoplasmic effectors are required for pathogenesis of Phytophthora sojae by suppression of host defenses. Plant Physiol. 2011, 155, 490–501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maximo, H.J.; Dalio, R.O.; Litholdo, C.G.; Felizatti, H.L.; Machado, M.A. PpCRN7 and PpCRN20 of Phythophthora parasitica regulate plant cell death leading to enhancement of host susceptibility. BMC Plant Biol. 2019, 19, 544. [Google Scholar] [CrossRef]
- Wang, S.; Welsh, L.; Thorpe, P.; Whisson, S.C.; Boevink, P.C.; Birch, P.R.J. The Phytophthora infestans haustorium is a site for secretion of diverse classes of infection-associated proteins. MBio 2018, 9, e01216-18. [Google Scholar] [CrossRef] [Green Version]
- Judelson, H.S.; Ah-Fong, A.M.V. Exchanges at the plant-oomycete interface that influence disease. Plant Physiol. 2019, 179, 1198–1211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, S.; Kong, G.; Qutob, D.; Yu, X.; Tang, J.; Kang, J.; Dai, T.; Wang, H.; Gijzen, M.; Wang, Y. The NLP toxin family in Phytophthora sojae includes rapidly evolving groups that lack necrosis-inducing activity. Mol. Plant Microbe Interact. 2012, 25, 896–909. [Google Scholar] [CrossRef] [Green Version]
- Ah-Fong, A.M.V.; Shrivastava, J.; Judelson, H.S. Lifestyle, gene gain and loss, and transcriptional remodeling cause divergence in the transcriptomes of Phytophthora infestans and Pythium ultimum during potato tuber colonization. BMC Genom. 2017, 18, 764. [Google Scholar] [CrossRef]
- Van Damme, M.; Bozkurt, T.O.; Cakir, C.; Schornack, S.; Sklenář, J.; Jones, A.M.E.; Kamoun, S. The Irish potato famine pathogen Phytophthora infestans translocates the CRN8 kinase into host plant cells. PLoS Pathog. 2012, 8, e1002875. [Google Scholar] [CrossRef]
- Li, Q.; Ai, G.; Shen, D.; Zou, F.; Wang, J.; Bai, T.; Chen, Y.; Li, S.; Zhang, M.; Jing, M.; et al. A Phytophthora capsici effector targets ACD11 binding partners that regulate ROS-mediated defense response in Arabidopsis. Mol. Plant 2019, 12, 565–581. [Google Scholar] [CrossRef] [Green Version]
- Toljamo, A.; Blande, D.; Munawar, M.; Kärenlampi, S.O.; Kokko, H. Expression of the GAF sensor, carbohydrate-active Enzymes, elicitins, and RXLRs differs markedly between two Phytophthora cactorum isolates. Phytopathology 2019, 109, 726–735. [Google Scholar] [CrossRef]
- Hardham, A.R.; Blackman, L.M. Phytophthora cinnamomi. Mol. Plant Pathol. 2018, 19, 260–285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamoun, S. A catalogue of the effector secretome of plant pathogenic oomycetes. Annu. Rev. Phytopathol. 2006, 44, 41–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gijzen, M.; Nürnberger, T. Nep1-like proteins from plant pathogens: Recruitment and diversification of the NPP1 domain across taxa. Phytochemistry 2006, 67, 1800–1807. [Google Scholar] [CrossRef]
- Lamour, K.; Kamoun, S. Oomycete Genetics and Genomics: Diversity, Interactions and Research Tools; John Wiley & Sons: Hoboken, NJ, USA, 2009; pp. 1–387. [Google Scholar]
- Wang, Y.; Hu, D.; Zhang, Z.; Ma, Z.; Zheng, X.; Li, D. Purification and immune cytolocalization of a novel Phytophthora boehmeriae protein inducing the hypersensitive response and systemic acquired resistance in tobacco and Chinese cabbage. Physiol. Mol. Plant Pathol. 2003, 63, 223–232. [Google Scholar] [CrossRef]
- Zhang, Z.-G.; Wang, Y.-C.; Li, J.; Ji, R.; Shen, G.; Wang, S.-C.; Zhou, X.; Zheng, X.-B. The role of SA in the hypersensitive response and systemic acquired resistance induced by elicitor PB90 from Phytophthora boehmeriae. Physiol. Mol. Plant Pathol. 2004, 65, 31–38. [Google Scholar] [CrossRef]
- Chen, Q.; Chen, Z.; Lu, L.; Jin, H.; Sun, L.; Yu, Q.; Xu, H.; Yang, F.; Fu, M.; Li, S.; et al. Interaction between abscisic acid and nitric oxide in PB90-induced catharanthine biosynthesis of Catharanthus roseus cell suspension cultures. Biotechnol. Prog. 2013, 29, 994–1001. [Google Scholar] [CrossRef]
- Huet, J.-C.; Pernollet, M.M.J.-C. Amino acid sequence of the α-elicitin secreted by Phytophthora cactorum. Phytochemistry 1993, 34, 1261–1264. [Google Scholar] [CrossRef]
- Chen, X.-R.; Zhang, B.-Y.; Xing, Y.-P.; Li, Q.-Y.; Li, Y.-P.; Tong, Y.-H.; Xu, J.-Y. Transcriptomic analysis of the phytopathogenic oomycete Phytophthora cactorum provides insights into infection-related effectors. BMC Genom. 2014, 15, 980. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.R.; Huang, S.X.; Zhang, Y.; Sheng, G.L.; Zhang, B.Y.; Li, Q.Y.; Zhu, F.; Xu, J.Y. Transcription profiling and identification of infection-related genes in Phytophthora cactorum. Mol. Genet. Genom. 2017, 293, 541–555. [Google Scholar] [CrossRef]
- Ricci, P.; Bonnet, P.; Huet, J.-C.; Sallantin, M.; Beauvais-Cante, F.; Bruneteau, M.; Billard, V.; Michel, G.; Pernollet, J.-C. Structure and activity of proteins from pathogenic fungi Phytophthora eliciting necrosis and acquired resistance in tobacco. Eur. J. Biochem. 1989, 183, 555–563. [Google Scholar] [CrossRef]
- Liu, Z.-Q.; Qiu, A.-L.; Shi, L.-P.; Cai, J.-S.; Huang, X.-Y.; Yang, S.; Wang, B.; Shen, L.; Huang, M.-K.; Mou, S.-L.; et al. SRC2-1 is required in PcINF1-induced pepper immunity by acting as an interacting partner of PcINF1. J. Exp. Bot. 2015, 66, 3683–3698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Z.-Q.; Liu, Y.-Y.; Shi, L.-P.; Yang, S.; Shen, L.; Yu, H.-X.; Wang, R.-Z.; Wen, J.-Y.; Tang, Q.; Hussain, A.; et al. SGT1 is required in PcINF1/SRC2-1 induced pepper defense response by interacting with SRC2-1. Sci. Rep. 2016, 6, 21651. [Google Scholar] [CrossRef]
- Billard, V.; Bruneteau, M.; Bonnet, P.; Ricci, P.; Pernollet, J.; Huet, J.; Vergne, A.; Richard, G.; Michel, G. Chromatographic purification and characterization of elicitors of necrosis on tobacco produced by incompatible Phytophthora species. J. Chromatogr. 1988, 440, 87–94. [Google Scholar] [CrossRef]
- Huet, J.-C.; Pernollet, J.-C. Amino acid sequence of cinnamomin, a new member of the elicitin family, and its comparison to cryptogein and capsicein. FEBS Lett. 1989, 257, 302–306. [Google Scholar] [CrossRef] [Green Version]
- Mishra, A.K.; Sharma, K.; Misra, R.S. Purification and characterization of elicitor protein from Phytophthora colocasiae and basic resistance in Colocasia esculenta. Microbiol. Res. 2009, 164, 688–693. [Google Scholar] [CrossRef]
- Galiana, E.; Bonnet, P.; Conrod, S.; Keller, H.; Panabieres, F.; Ponchet, M.; Poupet, A.; Ricci, P. RNase activity prevents the growth of a fungal pathogen in tobacco leaves and increases upon induction of systemic acquired resistance with elicitin. Plant Physiol. 1997, 115, 1557–1567. [Google Scholar] [CrossRef] [Green Version]
- Mikes, V.; Milat, M.-L.; Ponchet, M.; Ricci, P.; Blein, J.-P. The fungal elicitor cryptogein is a sterol carrier protein. FEBS Lett. 1997, 416, 190–192. [Google Scholar] [CrossRef] [Green Version]
- Leborgne-Castel, N.; Lherminier, J.; Der, C.; Fromentin, J.; Houot, V.; Simon-Plas, F. The plant defense elicitor cryptogein stimulates clathrin mediated endocytosis correlated with reactive oxygen species production in bright yellow-2 tobacco cells. Plant Physiol. 2008, 146, 1255–1266. [Google Scholar] [CrossRef] [Green Version]
- Coursol, S.; Fromentin, J.; Noirot, E.; Brière, C.; Robert, F.; Morel, J.; Liang, Y.; Lherminier, J.; Simon-Plas, F. Long-chain bases and their phosphorylated derivatives differentially regulate cryptogein-induced production of reactive oxygen species in tobacco (Nicotiana tabacum) BY-2 cells. New Phytol. 2015, 205, 1239–1249. [Google Scholar] [CrossRef]
- Kulik, A.; Noirot, E.; Grandperret, V.; Bourque, S.; Fromentin, J.; Salloignon, P.; Truntzer, C.; Dobrowolska, G.; Simon-Plas, F.; Wendehenne, D. Interplays between nitric oxide and reactive oxygen species in cryptogein signalling. Plant Cell Environ. 2015, 38, 331–348. [Google Scholar] [CrossRef]
- Ptáčková, N.; Klempová, J.; Obořil, M.; Nedělová, S.; Lochman, J.; Kašparovský, T. The effect of cryptogein with changed abilities to transfer sterols and altered charge distribution on extracellular alkalinization, ROS and NO generation, lipid peroxidation and LOX gene transcription in Nicotiana tabacum. Plant Physiol. Biochem. 2015, 97, 82–95. [Google Scholar] [CrossRef]
- Starý, T.; Satková, P.; Piterková, J.; Mieslerová, B.; Luhová, L.; Mikulík, J.; Kašparovský, T.; Petřivalský, M.; Lochman, J. The elicitin β-cryptogein’s activity in tomato is mediated by jasmonic acid and ethylene signalling pathways independently of elicitin-sterol interactions. Planta 2019, 249, 739–749. [Google Scholar] [CrossRef] [PubMed]
- Huet, J.-C.; Nespoulous, C.; Pernollet, J.-C. Structures of elicitin isoforms secreted by Phytophthora drechsleri. Phytochemistry 1992, 31, 1471–1476. [Google Scholar] [CrossRef]
- Capasso, R.; Di Maro, A.; Cristinzio, G.; De Martino, A.; Chambery, A.; Daniele, A.; Sannino, F.; Testa, A.; Parente, A. Isolation, characterization and structure-elicitor activity relationships of hibernalin and its two oxidized forms from Phytophthora hibernalis. Carne 1925. J. Biochem. 2008, 143, 131–141. [Google Scholar] [CrossRef]
- Huet, J.; Sallé-Tourne, M.; Pernollet, J. Amino acid sequence and toxicity of the alpha elicitin secreted with ubiquitin by Phytophthora infestans. Mol. Plant Microbe Interact. 1994, 7, 302–304. [Google Scholar] [CrossRef] [PubMed]
- Kamoun, S.; van West, P.; de Jong, A.J.; de Groot, K.E.; Vleeshouwers, V.G.A.A.; Govers, F. A gene encoding a protein elicitor of Phytophthora infestans is down-regulated during infection of potato. Mol. Plant Microbe Interact. 1997, 10, 13–20. [Google Scholar] [CrossRef] [Green Version]
- Kamoun, S.; Van West, P.; Vleeshouwers, V.G.A.A.; De Groot, K.E.; Govers, F. Resistance of Nicotiana benthamiana to Phytophthora infestans is mediated by the recognition of the elicitor protein INF1. Plant Cell 1998, 10, 1413–1425. [Google Scholar] [CrossRef] [Green Version]
- Kanzaki, H.; Saitoh, H.; Ito, A.; Fujisawa, S.; Kamoun, S.; Katou, S.; Yoshioka, H.; Terauchi, R. Cytosolic HSP90 and HSP70 are essential components of INF1-mediated hypersensitive response and non-host resistance to Pseudomonas cichorii in Nicotiana benthamiana. Mol. Plant Pathol. 2003, 4, 383–391. [Google Scholar] [CrossRef] [Green Version]
- Huitema, E.; Vleeshouwers, V.G.; Cakir, C.; Kamoun, S.; Govers, F. Differences in intensity and specificity of hypersensitive response induction in Nicotiana spp. by INF1, INF2A, and INF2B of Phytophthora infestans. Mol. Plant Microbe Interact. 2005, 18, 183–193. [Google Scholar] [CrossRef] [Green Version]
- Du, J.; Verzaux, E.; Chaparro-Garcia, A.; Bijsterbosch, G.; Keizer, L.C.P.; Zhou, J.; Liebrand, T.W.H.; Xie, C.; Govers, F.; Robatzek, S.; et al. Elicitin recognition confers enhanced resistance to Phytophthora infestans in potato. Nat. Plants 2015, 1, 15034. [Google Scholar] [CrossRef]
- Huet, J.-C.; Pernollet, J.-C. Sequences of acidic and basic elicitin isoforms secreted by Phytophthora megasperma. Phytochemistry 1993, 33, 797–805. [Google Scholar] [CrossRef]
- Baillieul, F.; de Ruffray, P.; Kauffmann, S. Molecular cloning and biological activity of α-, β-, and γ-megaspermin, three elicitins secreted by Phytophthora megasperma H20. Plant Physiol. 2003, 131, 155–166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baillieul, F.; Genetet, I.; Kopp, M.; Saindrenan, P.; Fritig, B.; Kauffmann, S. A new elicitor of the hypersensitive response in tobacco: A fungal glycoprotein elicits cell death, expression of defence genes, production of salicylic acid, and induction of systemic acquired resistance. Plant J. 1995, 8, 551–560. [Google Scholar] [CrossRef] [PubMed]
- Churngchow, N.; Rattarasarn, M. The elicitin secreted by Phytophthora palmivora, a rubber tree pathogen. Phytochemistry 2000, 54, 33–38. [Google Scholar] [CrossRef]
- Nespoulous, C.; Huet, J.-C.; Pernollet, J.-C. Structure-function relationships of α and β elicitins, signal proteins involved in the plant-Phytophthora interaction. Planta 1992, 186, 551–557. [Google Scholar] [CrossRef]
- Kamoun, S.; Klucher, K.M.; Coffey, M.D.; Tyler, B.M. A gene encoding a host-specific elicitor protein of Phytophthora parasitica. Mol. Plant Microbe Interact. 1993, 6, 573. [Google Scholar] [CrossRef]
- Mouton-Perronnet, F.; Bruneteau, M.; Denoroy, L.; Bouliteau, P.; Ricci, P.; Bonnet, P.; Michel, G. Elicitin produced by an isolate of Phytophthora parasitica pathogenic to tobacco. Phytochemistry 1995, 38, 41–44. [Google Scholar] [CrossRef]
- Capasso, R.; Cristinzio, G.; Evidente, A.; Visca, C.; Ferranti, P.; Blanco, F.D.V.; Parente, A. Elicitin 172 from an isolate of Phytophthora nicotianae pathogenic to tomato. Phytochemistry 1999, 50, 703–709. [Google Scholar] [CrossRef]
- Capasso, R.; Cristinzio, G.; Di Maro, A.; Ferranti, P.; Parente, A. Syringicin, a new α-elicitin from an isolate of Phytophthora syringae, pathogenic to citrus fruit. Phytochemistry 2001, 58, 257–262. [Google Scholar] [CrossRef]
- Chen, X.-R.; Huang, S.-X.; Zhang, Y.; Sheng, G.-L.; Li, Y.-P.; Zhu, F. Identification and functional analysis of the NLP-encoding genes from the phytopathogenic oomycete Phytophthora capsici. Mol. Genet. Genom. 2018, 293, 931–943. [Google Scholar] [CrossRef]
- Feng, B.-Z.; Zhu, X.-P.; Fu, L.; Lv, R.-F.; Storey, D.; Tooley, P.; Zhang, X.-G. Characterization of necrosis-inducing NLP proteins in Phytophthora capsici. BMC Plant Biol. 2014, 14, 126. [Google Scholar] [CrossRef] [Green Version]
- Kanneganti, T.-D.; Huitema, E.; Cakir, C.; Kamoun, S. Synergistic interactions of the plant cell death pathways induced by Phytophthora infestans Nep1-like protein PiNPP1.1 and INF1 elicitin. Mol. Plant Microbe Interact. 2006, 19, 854–863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fellbrich, G.; Romanski, A.; Varet, A.; Blume, B.; Brunner, F.; Engelhardt, S.; Felix, G.; Kemmerling, B.; Krzymowska, M.; Nürnberger, T. NPP1, a Phytophthora-associated trigger of plant defense in parsley and Arabidopsis. Plant J. 2002, 32, 375–390. [Google Scholar] [CrossRef]
- Qutob, D.; Kemmerling, B.; Brunner, F.; Kufner, I.; Engelhardt, S.; Gust, A.A.; Luberacki, B.; Seitz, H.U.; Stahl, D.; Rauhut, T.; et al. Phytotoxicity and innate immune responses induced by Nep1-like proteins. Plant Cell 2006, 18, 3721–3744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ottmann, C.; Luberacki, B.; Küfner, I.; Koch, W.; Brunner, F.; Weyand, M.; Mattinen, L.; Pirhonen, M.; Anderluh, G.; Seitz, H.U.; et al. A common toxin fold mediates microbial attack and plant defense. Proc. Natl. Acad. Sci. USA 2009, 106, 10359–10364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Böhm, H.; Albert, I.; Oome, S.; Raaymakers, T.M.; Van Den Ackerveken, G.; Nürnberger, T. A conserved peptide pattern from a widespread microbial virulence factor triggers pattern-induced immunity in Arabidopsis. PLoS Pathog. 2014, 10, e1004491. [Google Scholar] [CrossRef] [PubMed]
- Albert, I.; Böhm, H.; Albert, M.; Feiler, C.E.; Imkampe, J.; Wallmeroth, N.; Brancato, C.; Raaymakers, T.M.; Oome, S.; Zhang, H.; et al. An RLP23-SOBIR1-BAK1 complex mediates NLP triggered immunity. Nat. Plants. 2015, 1, 15140. [Google Scholar] [CrossRef]
- Qutob, D.; Kamoun, S.; Gijzen, M. Expression of a Phytophthora sojae necrosis-inducing protein occurs during transition from biotrophy to necrotrophy. Plant J. 2002, 32, 361–373. [Google Scholar] [CrossRef] [Green Version]
- Veit, S.; Wörle, J.M.; Nürnberger, T.; Koch, W.; Seitz, H.U. A novel protein elicitor (PaNie) from Pythium aphanidermatum induces multiple defense responses in carrot, Arabidopsis, and tobacco. Plant Physiol. 2001, 127, 832–841. [Google Scholar] [CrossRef]
- Mateos, F.V.; Rickauer, M.; Esquerré-Tugayé, M.-T. Cloning and characterization of a cDNA encoding an elicitor of Phytophthora parasitica var. nicotianae that shows cellulose-binding and lectin-like activities. Mol. Plant Microbe Interact. 1997, 10, 1045–1053. [Google Scholar] [CrossRef] [Green Version]
- Khatib, M.; Lafitte, C.; Esquerré-Tugayé, M.; Bottin, A.; Rickauer, M. The CBEL elicitor of Phytophthora parasitica var. nicotianae activates defence in Arabidopsis thaliana via three different signalling pathways. New Phytol. 2004, 162, 501–510. [Google Scholar] [CrossRef]
- Fu, L.; Zhu, C.; Ding, X.; Yang, X.; Morris, P.F.; Tyler, B.M.; Zhang, X. Characterization of cell-death-inducing members of the pectate lyase gene family in Phytophthora capsici and their contributions to infection of pepper. Mol. Plant Microbe Interact. 2015, 28, 766–775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, Z.; Song, T.; Zhu, L.; Ye, W.; Wang, Y.; Shao, Y.; Dong, S.; Zhang, Z.; Dou, D.; Zheng, X.; et al. A Phytophthora sojae glycoside hydrolase 12 protein is a major virulence factor during soybean infection and is recognized as a PAMP. Plant Cell 2015, 27, 2057–2072. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Xu, Y.; Sun, Y.; Wang, H.; Qi, J.; Wan, B.; Ye, W.; Lin, Y.; Shao, Y.; Dong, S.; et al. Leucine rich repeat receptor-like gene screen reveals that Nicotiana RXEG1 regulates glycoside hydrolase 12 MAMP detection. Nat. Commun. 2018, 9, 594. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.-H.; Yan, H.-Z.; Liou, R.-F. A novel elicitor protein from Phytophthora parasitica induces plant basal immunity and systemic acquired resistance. Mol. Plant Pathol. 2015, 16, 123–136. [Google Scholar] [CrossRef] [PubMed]
- Orsomando, G.; Lorenzi, M.; Raffaelli, N.; Rizza, M.D.; Mezzetti, B.; Ruggieri, S. Phytotoxic protein PcF, purification, characterization, and cDNA sequencing of a novel hydroxyproline-containing factor secreted by the strawberry pathogen Phytophthora cactorum. J. Biol. Chem. 2001, 276, 21578–21584. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.-R.; Li, Y.-P.; Li, Q.-Y.; Xing, Y.-P.; Liu, B.-B.; Tong, Y.-H.; Xu, J.-Y. SCR96, a small cysteine-rich secretory protein of Phytophthora cactorum, can trigger cell death in the Solanaceae and is important for pathogenicity and oxidative stress tolerance. Mol. Plant Pathol. 2016, 17, 577–587. [Google Scholar] [CrossRef]
- Duclos, J.; Fauconnier, A.; Coelho, A.-C.; Bollen, A.; Cravador, A.; Godfroid, E. Identification of an elicitin gene cluster in Phytophthora cinnamomi. DNA Seq. 1998, 9, 231–237. [Google Scholar] [CrossRef]
- Jiang, R.H.Y.; Tyler, B.M.; Whisson, S.C.; Hardham, A.R.; Govers, F. Ancient origin of elicitin gene clusters in Phytophthora genomes. Mol. Biol. Evol. 2006, 23, 338–351. [Google Scholar] [CrossRef] [Green Version]
- Osman, H.; Vauthrin, S.; Mikes, V.; Milat, M.-L.; Panabières, F.; Marais, A.; Brunie, S.; Maume, B.; Ponchet, M.; Blein, J.-P. Mediation of elicitin activity on tobacco is assumed by elicitin-sterol complexes. Mol. Biol. Cell 2001, 12, 2825–2834. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues, M.L.; Archer, M.; Martel, P.; Miranda, S.; Thomaz, M.; Enguita, F.J.; Baptista, R.P.; e Melo, E.P.; Sousa, N.; Cravador, A.; et al. Crystal structures of the free and sterol-bound forms of β-cinnamomin. Biochim. Biophys. Acta Proteins Proteom. 2006, 1764, 110–121. [Google Scholar] [CrossRef] [PubMed]
- Boissy, G.; de La Fortelle, E.; Kahn, R.; Huet, J.-C.; Bricogne, G.; Pernollet, J.-C.; Brunie, S. Crystal structure of a fungal elicitor secreted by Phytophthora cryptogea, a member of a novel class of plant necrotic proteins. Structure 1996, 4, 1429–1439. [Google Scholar] [CrossRef] [Green Version]
- Derevnina, L.; Dagdas, Y.F.; De la Concepcion, J.C.; Białas, A.; Kellner, R.; Petre, B.; Domazakis, E.; Du, J.; Wu, C.-H.; Lin, X.; et al. Nine things to know about elicitins. New Phytol. 2016, 212, 888–895. [Google Scholar] [CrossRef] [Green Version]
- Nie, J.; Yin, Z.; Li, Z.; Wu, Y.; Huang, L. A small cysteine-rich protein from two kingdoms of microbes is recognized as a novel pathogen-associated molecular pattern. New Phytol. 2019, 222, 995–1011. [Google Scholar] [CrossRef]
- Dokládal, L.; Obořil, M.; Stejskal, K.; Zdráhal, Z.; Ptáčková, N.; Chaloupková, R.; Damborský, J.; Kašparovský, T.; Jeandroz, S.; Žd’árská, M.; et al. Physiological and proteomic approaches to evaluate the role of sterol binding in elicitin-induced resistance. J. Exp. Bot. 2012, 63, 2203–2215. [Google Scholar] [CrossRef] [PubMed]
- Adachi, H.; Nakano, T.; Miyagawa, N.; Ishihama, N.; Yoshioka, M.; Katou, Y.; Yaeno, T.; Shirasu, K.; Yoshioka, H. WRKY transcription factors phosphorylated by MAPK regulate a plant immune NADPH oxidase in Nicotiana benthamiana. Plant Cell 2015, 27, 2645–2663. [Google Scholar] [CrossRef] [Green Version]
- Bos, J.I.B.; Kanneganti, T.-D.; Young, C.; Cakir, C.; Huitema, E.; Win, J.; Armstrong, M.R.; Birch, P.R.J.; Kamoun, S. The C-terminal half of Phytophthora infestans RXLR effector AVR3a is sufficient to trigger R3a-mediated hypersensitivity and suppress INF1-induced cell death in Nicotiana benthamiana. Plant J. 2006, 48, 165–176. [Google Scholar] [CrossRef]
- Haas, B.J.; Kamoun, S.; Zody, M.C.; Jiang, R.H.Y.; Handsaker, R.E.; Cano, L.M.; Grabherr, M.; Kodira, C.D.; Raffaele, S.; Torto-Alalibo, T.; et al. Genome sequence and analysis of the Irish potato famine pathogen Phytophthora infestans. Nature 2009, 461, 393–398. [Google Scholar] [CrossRef]
- Masago, H. Selective inhibition of Pythium spp. on a medium for direct isolation of Phytophthora spp. from soils and plants. Phytopathology 1977, 67, 425–428. [Google Scholar] [CrossRef]
- Cabral, A.; Oome, S.; Sander, N.; Küfner, I.; Nürnberger, T.; Van den Ackerveken, G. Nontoxic Nep1-like proteins of the downy mildew pathogen Hyaloperonospora arabidopsidis: Repression of necrosis-inducing activity by a surface-exposed region. Mol. Plant Microbe Interact. 2012, 25, 697–708. [Google Scholar] [CrossRef] [Green Version]
- Oome, S.; Van den Ackerveken, G. Comparative and functional analysis of the widely occurring family of Nep1-like proteins. Mol. Plant Microbe Interact. 2014, 27, 1081–1094. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lenarčič, T.; Albert, I.; Böhm, H.; Hodnik, V.; Pirc, K.; Zavec, A.B.; Podobnik, M.; Pahovnik, D.; Žagar, E.; Pruitt, R.; et al. Eudicot plant-specific sphingolipids determine host selectivity of microbial NLP cytolysins. Science 2017, 358, 1431–1434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lenarčič, T.; Pric, K.; Hodnik, V.; Albert, I.; Borišek, J.; Magistrato, A.; Nürnberger, T.; Podobnik, M.; Anderluh, G. Molecular basis for functional diversity among microbial Nep1-like proteins. PLoS Pathog. 2019, 15, e1007951. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bae, H.; Bowers, J.H.; Tooley, P.W.; Bailey, B.A. NEP1 orthologs encoding necrosis and ethylene inducing proteins exist as a multigene family in Phytophthora megakarya, causal agent of black pod disease on cacao. Mycol. Res. 2005, 109, 1373–1385. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.-R.; Xing, Y.-P.; Li, Y.-P.; Tong, Y.-H.; Xu, J.-Y. RNA-Seq reveals infection-related gene expression changes in Phytophthora capsici. PLoS ONE 2013, 8, e74588. [Google Scholar] [CrossRef]
- Martins, I.M.; Meirinho, S.; Costa, R.; Cravador, A.; Choupina, A. Cloning, characterization, in vitro and in planta expression of a necrosis-inducing Phytophthora protein 1 gene npp1 from Phytophthora cinnamomi. Mol. Biol. Rep. 2019, 46, 6453–6462. [Google Scholar] [CrossRef] [Green Version]
- Win, J.; Morgan, W.; Bos, J.; Krasileva, K.; Cano, L.M.; Chaparro-Garcia, A.; Ammar, R.; Staskawicz, B.J.; Kamoun, S. Adaptive evolution has targeted the C-terminal domain of the RXLR effectors of plant pathogenic oomycetes. Plant Cell 2007, 19, 2349–2369. [Google Scholar] [CrossRef] [Green Version]
- Dou, D.; Kale, S.D.; Wang, X.; Jiang, R.H.; Bruce, N.A.; Arredondo, F.D.; Zhang, X.; Tyler, B.M. RXLR-mediated entry of Phytophthora sojae effector Avr1b into soybean cells does not require pathogen-encoded machinery. Plant Cell 2008, 20, 1930–1947. [Google Scholar] [CrossRef] [Green Version]
- Kale, S.D.; Gu, B.; Capelluto, D.G.; Dou, D.; Feldman, E.; Rumore, A.; Arredondo, F.D.; Hanlon, R.; Fudal, I.; Rouxel, T.; et al. External lipid PI3P mediates entry of eukaryotic pathogen effectors into plant and animal host cells. Cell 2010, 142, 284–295. [Google Scholar] [CrossRef] [Green Version]
- Joubert, M.; Backer, R.; Engelbrecht, J.; Berg, N.V.D. Expression of several Phytophthora cinnamomi putative RxLRs provides evidence for virulence roles in avocado. PLoS ONE 2021, 16, e0254645. [Google Scholar] [CrossRef]
- Wang, S.; McLellan, H.; Bukharova, T.; He, Q.; Murphy, F.; Shi, J.; Sun, S.; van Weymers, P.; Ren, Y.; Thilliez, G.; et al. Phytophthora infestans RXLR effectors act in concert at diverse subcellular locations to enhance host colonization. J. Exp. Bot. 2019, 70, 343–356. [Google Scholar] [CrossRef] [Green Version]
- Yang, B.; Wang, Q.; Jing, M.; Guo, B.; Wu, J.; Wang, H.; Wang, Y.; Lin, L.; Ye, W.; Dong, S.; et al. Distinct regions of the Phytophthora essential effector Avh238 determine its function in cell death activation and plant immunity suppression. New Phytol. 2017, 214, 361–375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Wang, H.; Jing, M.; Zhu, J.; Guo, B.; Wang, Y.; Lin, Y.; Chen, H.; Kong, L.; Ma, Z.; et al. A Phytophthora effector recruits a host cytoplasmic transacetylase into nuclear speckles to enhance plant susceptibility. eLife 2018, 7, e40039. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.-R.; Zhang, Y.; Li, H.-Y.; Zhang, Z.-H.; Sheng, G.-L.; Li, Y.-P.; Xing, Y.-P.; Huang, S.-X.; Tao, H.; Kuan, T.; et al. The RXLR effector PcAvh1 is required for full virulence of Phytophthora capsici. Mol. Plant Microbe Interact. 2019, 32, 986–1000. [Google Scholar] [CrossRef]
- Branco, I.; Choupina, A. In Silico characterization of the Phytopathogenic efector, Avr3a, from Phytophthora cinnamomi. J. Basic Appl. Sci. 2020, 16, 20–30. [Google Scholar] [CrossRef]
- Torto, T.A.; Li, S.; Styler, A.; Huitema, E.; Testa, A.; Gow, N.A.R.; van West, P.; Kamoun, S. EST mining and functional expression assays identify extracellular effector proteins from the plant pathogen Phytophthora. Genome Res. 2003, 13, 1675–1685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schornack, S.; van Damme, M.; Bozkurt, T.O.; Cano, L.M.; Smoker, M.; Thines, M.; Gaulin, E.; Kamoun, S.; Huitema, E. Ancient class of translocated oomycete effectors targets the host nucleus. Proc. Natl. Acad. Sci. USA 2010, 107, 17421–17426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stam, R.; Howden, A.J.M.; Delgado-Cerezo, M.; Amaro, T.M.M.M.; Motion, G.B.; Pham, J.; Huitema, E. Characterization of cell death inducing Phytophthora capsici CRN effectors suggests diverse activities in the host nucleus. Front. Plant Sci. 2013, 4, 387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amaro, T.M.M.M.; Thilliez, G.J.A.; Motion, G.B.; Huitema, E. A perspective on CRN Proteins in the genomics age: Evolution, classification, delivery and function revisited. Front. Plant Sci. 2017, 8, 99. [Google Scholar] [CrossRef] [Green Version]
- Cheung, F.; Win, J.; Lang, J.M.; Hamilton, J.; Vuong, H.; Leach, J.E.; Kamoun, S.; Lévesque, C.A.; Tisserat, N.; Buell, C.R. Analysis of the Pythium ultimum transcriptome using Sanger and pyrosequencing approaches. BMC Genom. 2008, 9, 542. [Google Scholar] [CrossRef] [Green Version]
- Gaulin, E.; Madoui, M.-A.; Bottin, A.; Jacquet, C.; Mathé, C.; Couloux, A.; Wincker, P.; Dumas, B. Transcriptome of Aphanomyces euteiches: New oomycete putative pathogenicity factors and metabolic pathways. PLoS ONE 2008, 3, e1723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lévesque, C.A.; Brouwer, H.; Cano, L.; Hamilton, J.P.; Holt, C.; Huitrma, E.; Raffaele, S.; Robideau, G.P.; Thines, M.; Win, J.; et al. Genome sequence of the necrotrophic plant pathogen Pythium ultimum reveals original pathogenicity mechanisms and effector repertoire. Genome Biol. 2010, 11, R73. [Google Scholar] [CrossRef] [PubMed]
- Meyer, F.E.; Shuey, L.S.; Naidoo, S.; Mamni, T.; Berger, D.K.; Myburg, A.A.; Berg, N.V.D.; Naidoo, S. Dual RNA-sequencing of Eucalyptus nitens during Phytophthora cinnamomi challenge reveals pathogen and host factors influencing compatibility. Front. Plant Sci. 2016, 7, 191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, D.; Burroughs, A.M.; Vidal, N.D.; Iyer, L.M.; Aravind, L. Transposons to toxins: The provenance, architecture and diversification of a widespread class of eukaryotic effectors. Nucleic Acids Res. 2016, 44, 3513–3533. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Zhang, M.; Shen, D.; Liu, T.; Chen, Y.; Zhou, J.M.; Dou, D. A Phytophthora sojae effector PsCRN63 forms homo-/hetero-dimers to suppress plant immunity via an inverted association manner. Sci. Rep. 2016, 6, 26951. [Google Scholar] [CrossRef] [Green Version]
- Bendahmane, A.; Querci, M.; Kanyuka, K.; Baulcombe, D.C. Agrobacterium transient expression system as a tool for the isolation of disease resistance genes: Application to the Rx2 locus in potato. Plant J. 2000, 21, 73–81. [Google Scholar] [CrossRef]
- Van der Hoorn, R.A.L.; Laurent, F.; Roth, R.; De Wit, P.J.G.M. Agroinfiltration is a versatile tool that facilitates comparative analyses of Avr 9/Cf-9-induced and Avr4/Cf-4-induced necrosis. Mol. Plant Microbe Interact. 2000, 13, 439–446. [Google Scholar] [CrossRef] [Green Version]
- Wroblewski, T.; Tomczak, A.; Michelmore, R. Optimization of Agrobacterium-mediated transient assays of gene expression in lettuce, tomato and Arabidopsis. Plant Biotechnol. J. 2005, 3, 259–273. [Google Scholar] [CrossRef]
- Lee, M.W.; Yang, Y. Transient expression assay by agroinfiltration of leaves. In Arabidopsis Protocols; Springer: New York, NY, USA, 2006; pp. 225–229. [Google Scholar]
- Ramirez-Garcés, D.; Camborde, L.; Pel, M.J.C.; Jauneau, A.; Martinez, Y.; Néant, I.; Leclerc, C.; Moreau, M.; Dumas, B.; Gaulin, E. CRN 13 candidate effectors from plant and animal eukaryotic pathogens are DNA-binding proteins which trigger host DNA damage response. New Phytol. 2016, 210, 602–617. [Google Scholar] [CrossRef] [Green Version]
- Goodin, M.M.; Zaitlin, D.; Naidu, R.A.; Lommel, S.A. Nicotiana benthamiana: Its history and future as a model for plant–pathogen interactions. Mol. Plant Microbe Interact. 2008, 21, 1015–1026. [Google Scholar] [CrossRef] [Green Version]
- Kamoun, S.; Goodwin, S.B. Fungal and oomycete genes galore. New Phytol. 2007, 174, 713–717. [Google Scholar] [CrossRef] [PubMed]
- Studholme, D.; McDougal, R.; Sambles, C.; Hansen, E.; Hardy, G.; Grant, M.; Ganley, R.J.; Williams, N.M. Genome sequences of six Phytophthora species associated with forests in New Zealand. Genom. Data 2016, 7, 54–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langmuir, A.L.; Beech, P.L.; Richardson, M.F. Draft genomes of two Australian strains of the plant pathogen, Phytophthora cinnamomi. F1000Research 2017, 6, 1972. [Google Scholar] [CrossRef]
- Kunjeti, S.G.; Evans, T.A.; Marsh, A.G.; Gregory, N.F.; Kunjeti, S.; Meyers, B.C.; Kalavacharla, V.S.; Donofrio, N.M. RNA-Seq reveals infection-related global gene changes in Phytophthora phaseoli, the causal agent of lima bean downy mildew. Mol. Plant Pathol. 2012, 13, 454–466. [Google Scholar] [CrossRef]
- Westermann, A.J.; Gorski, S.A.; Vogel, J. Dual RNA-seq of pathogen and host. Nat. Rev. Microbiol. 2012, 10, 618–630. [Google Scholar] [CrossRef]
- Hayden, K.J.; Garbelotto, J.M.; Knaus, B.J.; Cronn, R.C.; Wright, J.W. Dual RNA-seq of the plant pathogen Phytophthora ramorum and its tanoak host. Tree Genet. Genomes 2014, 10, 489–502. [Google Scholar] [CrossRef] [Green Version]
- Reitmann, A.; Berger, D.K.; Berg, N.V.D. Putative pathogenicity genes of Phytophthora cinnamomi identified via RNA-Seq analysis of pre-infection structures. Eur. J. Plant Pathol. 2017, 147, 211–228. [Google Scholar] [CrossRef] [Green Version]
- Judelson, H.S. Recent advances in the genetics of oomycete plant pathogens. Mol. Plant Microbe Interact. 1996, 9, 443–449. [Google Scholar] [CrossRef]
- Dai, T.; Xu, Y.; Yang, X.; Jiao, B.; Qiu, M.; Xue, J.; Arredondo, F.; Tyler, B.M. An improved transformation system for Phytophthora cinnamomi using green fluorescent protein. Front. Microbiol. 2021, 12, 682754. [Google Scholar] [CrossRef]
- Bailey, A.M.; Mena, G.L.; Herrera-Estrella, L.R. Genetic transformation of the plant pathogens Phytophthora capsici and Phytophthora parasitica. Nucleic Acids Res. 1991, 19, 4273–4278. [Google Scholar] [CrossRef] [Green Version]
- Judelson, H.S.; Tyler, B.M.; Michelmore, R.W. Transformation of the oomycete pathogen, Phytophthora infestans. Mol. Plant-Microbe Interact. 1991, 4, 602–607. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein Family | Plant Cell Surface Receptor | Co-Receptor | Protein | Phytophthora spp. | Function | References |
|---|---|---|---|---|---|---|
| ND | - | - | PB90 | Phytophthora boehmeriae | Induces cell death | [67,68,69] |
| Elicitin | ELR | BAK1, HSP70, HSP90, NbLRK1, SGT1, SRC2-1 | Cacto | Phytophthora cactorum | Induces cell death | [70] |
| PcELL1 | Induces cell death | [71] | ||||
| PcINF1 | Induces cell death | [72] | ||||
| Capsicein | Phytophthora capsici | Induces cell death and increases defense against P. nicotianae in Nicotiana benthamiana | [73] | |||
| PcINF1 | Induces cell death and pepper defense response | [74,75] | ||||
| Cinnamomin | Phytophthora cinnamomi | Induces cell death and protects N. benthamiana against pathogens | [76,77] | |||
| 15-kDa glycoprotein | Phytophthora colocasiae | Induces cell death and SAR | [78] | |||
| Cryptogein | Phytophthora cryptogea | Induces cell death, SAR and defense of N. benthamiana against P. nicotianae | [73,79,80,81,82,83,84,85] | |||
| Dreα, Dreβ | Phytophthora drechsleri | Induces cell death | [86] | |||
| Hibernalin1 | Phytophthora hibernalis | Induces cell death | [87] | |||
| INF1 | Phytophthora infestans | Triggers HR dependent on HSP70, HSP90 and SGT1 | [88,89,90,91,92,93] | |||
| INF2A, INF2B | INF2A-induced necrosis dependent on SGT1 | [92] | ||||
| MgMα, MgMβ | Phytophthora megasperma | Induces cell death | [94] | |||
| α-megaspermin, β-megaspermin, γ-megaspermin/32 kDa glycoprotein | Induces cell death, PR gene expression and SAR | [95,96] | ||||
| Palmivorein | Phytophthora palmivora | Induces cell death | [97] | |||
| Parasiticein/parA1/elicitin 310/elicitin 172 | Phytophthora parasitica | Induces cell death | [98,99,100,101] | |||
| Syringicin | Phytophthora syringae | Induces HR and electrolyte leakage in N. benthamiana | [102] | |||
| NLP | RLP23 | BAK1, COI1, HSP90, MEK2, NPR1, SGT1, SOBIR1 and TGA2.2 | PcNLP1 | P. cactorum | Induces cell death | [71] |
| Pc11951, Pc107869, Pc109174, Pc118548 | P. capsici | Induces cell death | [103] | |||
| PcNLP1 to 3, 6 to 10, 13 to 15 | Induces cell death | [104] | ||||
| PiNPP1.1 | P. infestans | Induces HR dependent on SGT1 and HSP90 | [105] | |||
| PpNLP/NLPPp | P. parasitica | Induces cell death | [106,107,108,109,110] | |||
| PsojNIP | Phytophthora sojae | Induces cell death dependent on SGT1 and HSP90 | [105,111] | |||
| PaNie213/NLPPya | Phytophthora aphanidermatum | Induces cell death | [107,108,112] | |||
| CBM | - | - | CBEL | P. parasitica | Induces cell death; activates defense responses via SA, JA and ET signaling pathways | [113,114] |
| PL | - | - | PcPL1, PcPL15, PcPL16, PcPL20 | P. capsici | Induces cell death | [115] |
| GH12 | RXEG1 | BAK1, SOBIR1 | XEG1 | P. sojae | Induces cell death; associates with SOBIR1 and BAK1 complex to trigger immune responses | [116,117] |
| GH16 | - | - | OPEL | P. parasitica | Induces cell death | [118] |
| PcF toxin | - | - | PcF | P. cactorum | Induces cell death and PR gene expression in N. benthamiana | [119] |
| SCR96, SCR99, SCR121 | Induces cell death | [120] | ||||
| SCR113 | Induces cell death | [72] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Midgley, K.A.; van den Berg, N.; Swart, V. Unraveling Plant Cell Death during Phytophthora Infection. Microorganisms 2022, 10, 1139. https://doi.org/10.3390/microorganisms10061139
Midgley KA, van den Berg N, Swart V. Unraveling Plant Cell Death during Phytophthora Infection. Microorganisms. 2022; 10(6):1139. https://doi.org/10.3390/microorganisms10061139
Chicago/Turabian StyleMidgley, Kayla A., Noëlani van den Berg, and Velushka Swart. 2022. "Unraveling Plant Cell Death during Phytophthora Infection" Microorganisms 10, no. 6: 1139. https://doi.org/10.3390/microorganisms10061139