
Adaptation of Cyanobacteria to the Endolithic Light Spectrum in Hyper-Arid Deserts

 , , ,
, , , 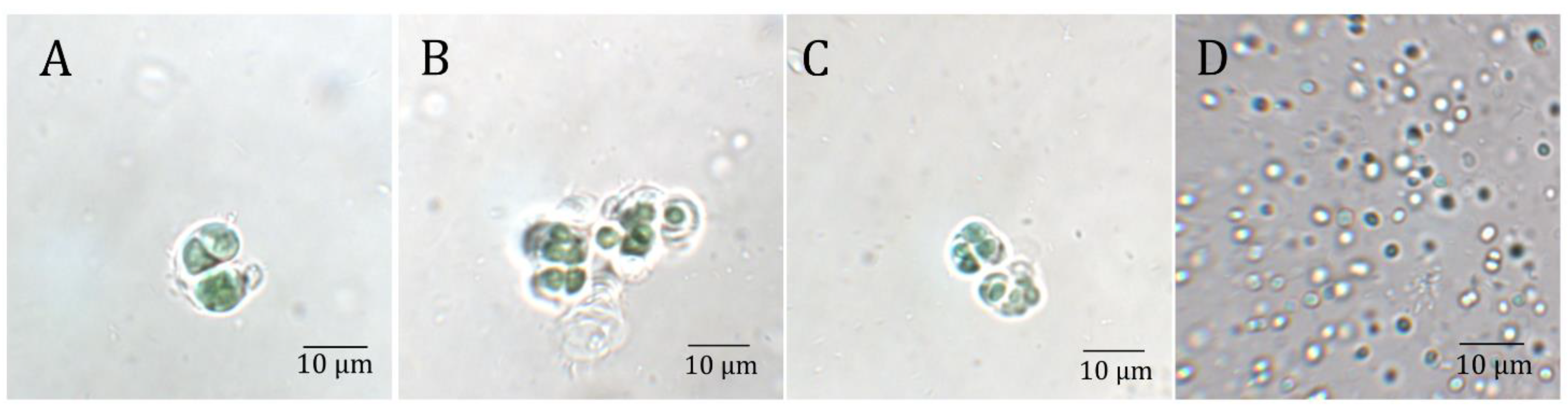{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
3. Results and Discussion
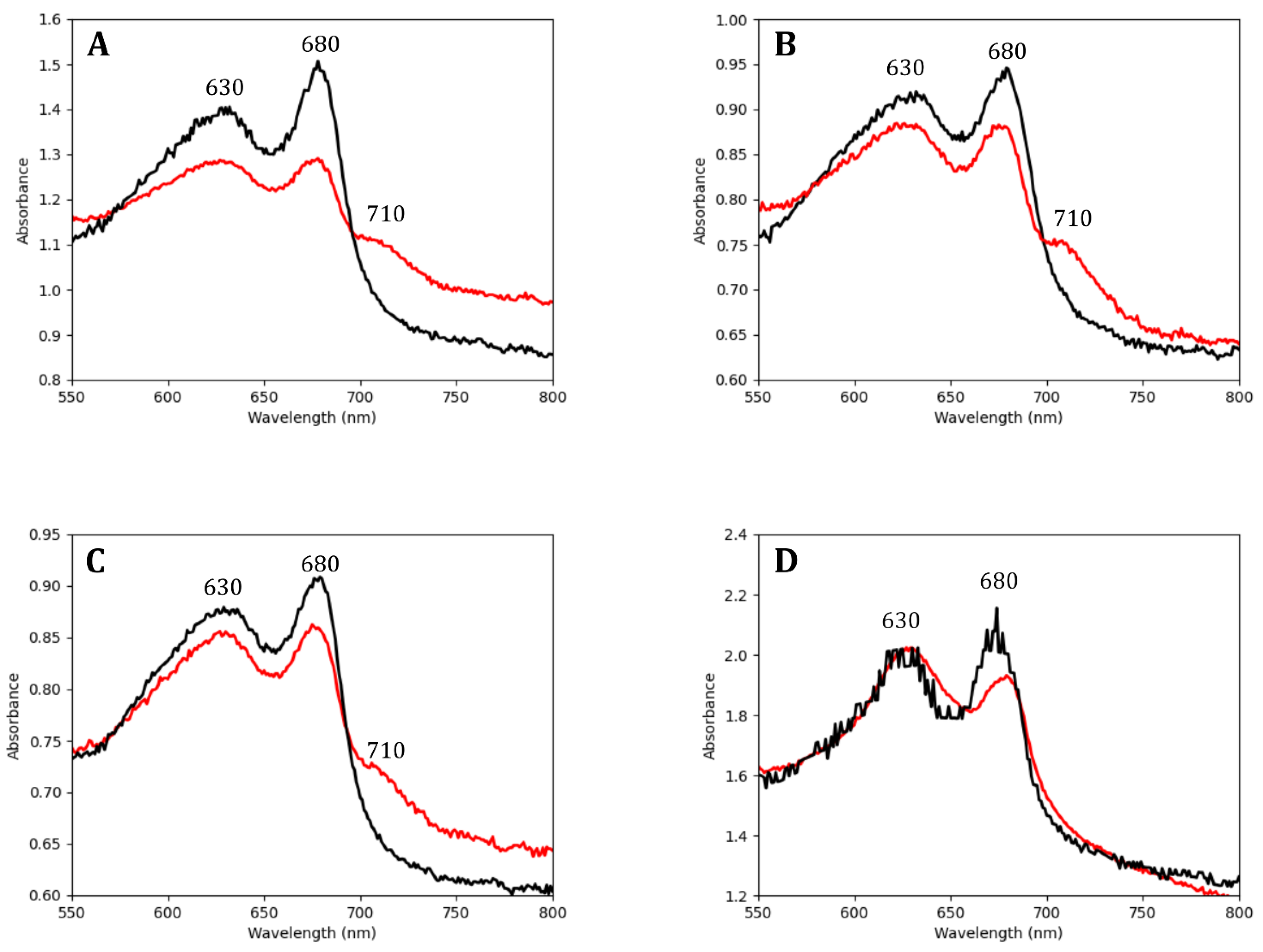
3.1. Endolithic Cyanobacteria Absorbed FRL Photons
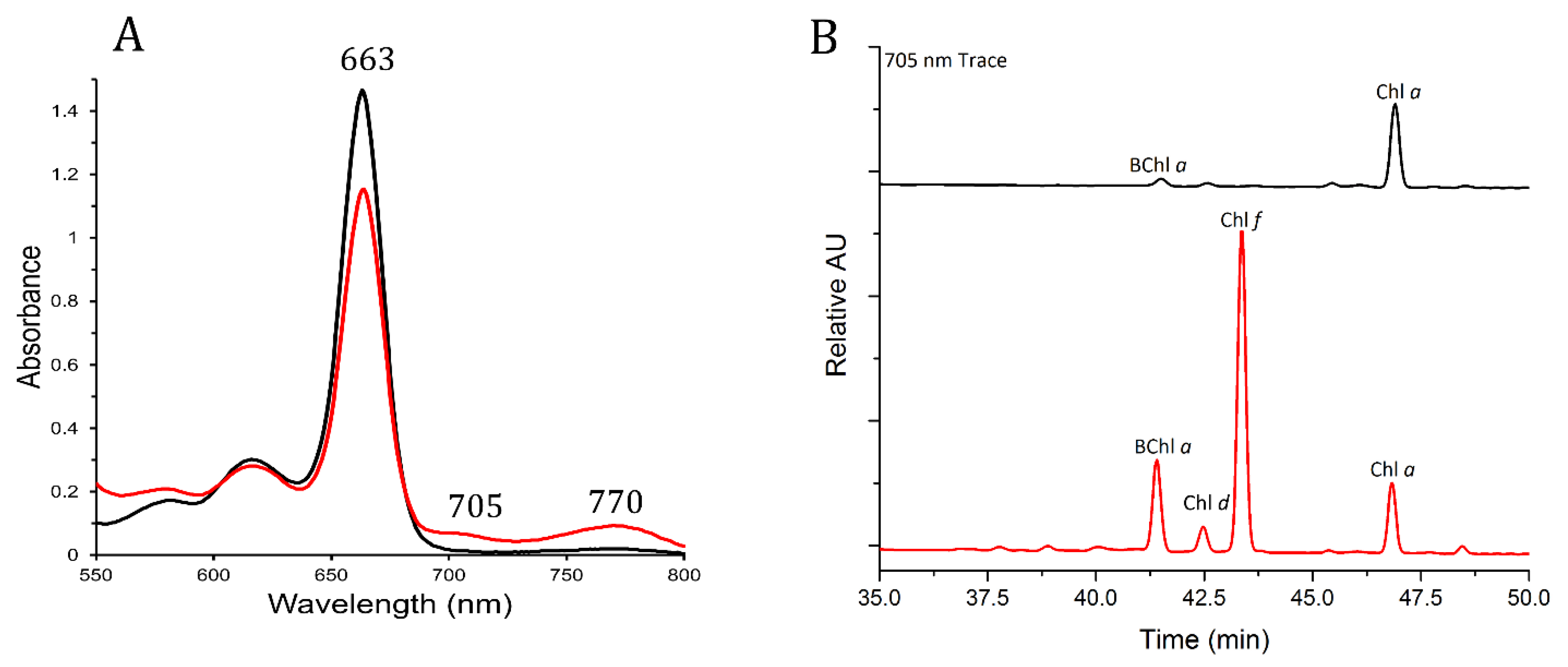
3.2. Pigments from FRL-Grown Chroococcidiopsis Contained Chl f and Chl d
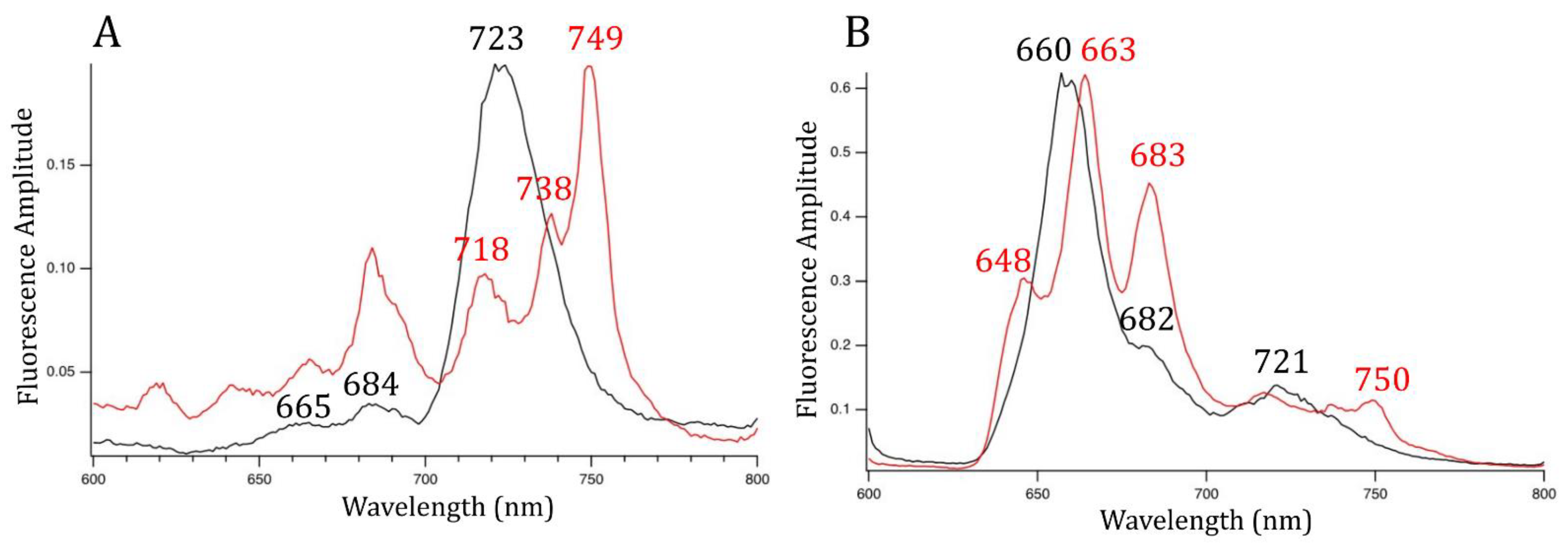
3.3. PSI, PSII, and PBS Have Different Spectral Properties after FRL Exposure
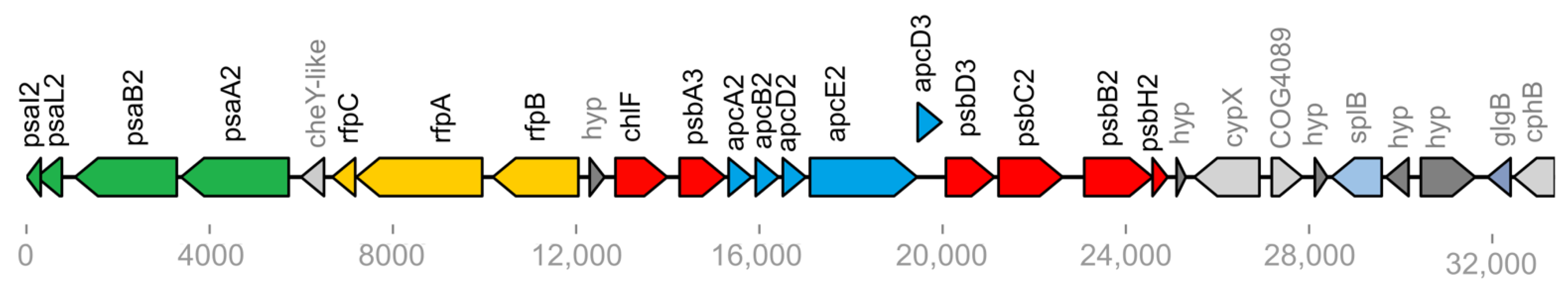
3.4. FaRLiP Gene Clusters Identified in Endolithic Chroococcidiopsis Metagenomes
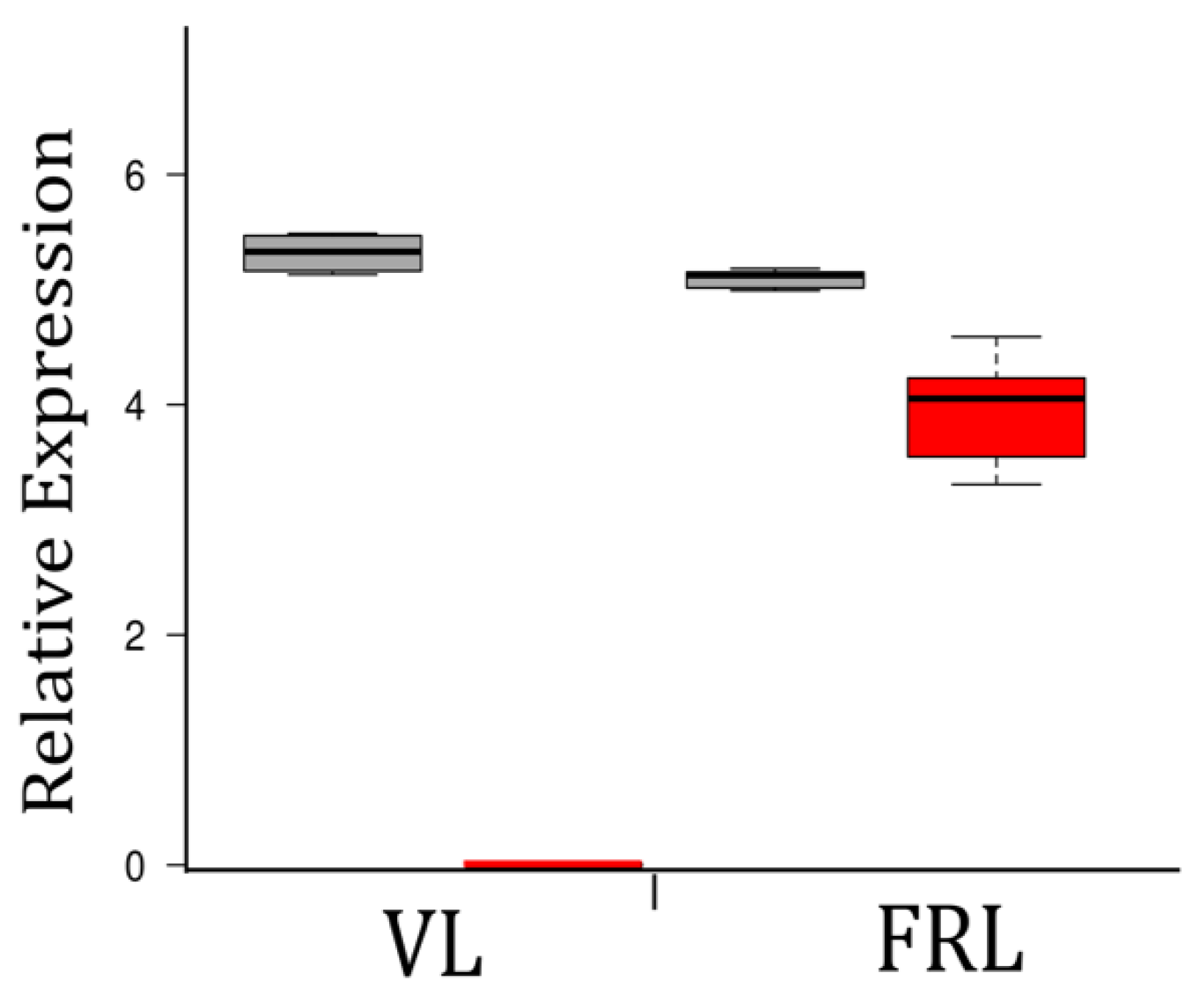
3.5. The chlF Gene Was Only Expressed in FRL-Grown Cultures
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pointing, S.B.; Belnap, J. Microbial Colonization and Controls in Dryland Systems. Nat. Rev. Microbiol. 2012, 10, 551–562. [Google Scholar] [CrossRef] [PubMed]
- Meslier, V.; DiRuggiero, J. Endolithic Microbial Communities as Model Systems for Ecology and Astrobiology. In Model Ecosystems in Extreme Environments; Seckbach, B., Rampelotto, P., Eds.; Elsevier: Amsterdam, The Netherlands, 2019; Volume 2, pp. 145–168. [Google Scholar]
- Walker, J.J.; Pace, N.R. Endolithic Microbial Ecosystems. Annu. Rev. Microbiol. 2007, 61, 331–347. [Google Scholar] [CrossRef] [PubMed]
- Meslier, V.; Casero, M.C.; Dailey, M.; Wierzchos, J.; Ascaso, C.; Artieda, O.; McCullough, P.R.; DiRuggiero, J. Fundamental Drivers for Endolithic Microbial Community Assemblies in the Hyperarid Atacama Desert. Environ. Microbiol. 2018, 20, 1765–1781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crits-Christoph, A.; Gelsinger, D.R.; Ma, B.; Wierzchos, J.; Ravel, J.; Davila, A.; Casero, M.C.; DiRuggiero, J. Functional Interactions of Archaea, Bacteria and Viruses in a Hypersaline Endolithic Community. Environ. Microbiol. 2016, 18, 2064–2077. [Google Scholar] [CrossRef]
- Uritskiy, G.; Munn, A.; Dailey, M.; Gelsinger, D.R.; Getsin, S.; Davila, A.; McCullough, P.R.; Taylor, J.; DiRuggiero, J. Environmental Factors Driving Spatial Heterogeneity in Desert Halophile Microbial Communities. Front. Microbiol. 2020, 11, 578669. [Google Scholar] [CrossRef]
- Ertekin, E.; Meslier, V.; Browning, A.; Treadgold, J.; DiRuggiero, J. Rock Structure Drives the Taxonomic and Functional Diversity of Endolithic Microbial Communities in Extreme Environments. Environ. Microbiol. 2021, 23, 3937–3956. [Google Scholar] [CrossRef]
- Goordial, J.; Altshuler, I.; Hindson, K.; Chan-Yam, K.; Marcolefas, E.; Whyte, L.G. In Situ Field Sequencing and Life Detection in Remote (79°26′ N) Canadian High Arctic Permafrost Ice Wedge Microbial Communities. Front. Microbiol. 2017, 8, 2594. [Google Scholar] [CrossRef]
- Archer, S.D.J.; de los Ríos, A.; Lee, K.C.; Niederberger, T.S.; Cary, S.C.; Coyne, K.J.; Douglas, S.; Lacap-Bugler, D.C.; Pointing, S.B. Endolithic Microbial Diversity in Sandstone and Granite from the McMurdo Dry Valleys, Antarctica. Polar Biol. 2017, 40, 997–1006. [Google Scholar] [CrossRef]
- Qu, E.B.; Omelon, C.R.; Oren, A.; Meslier, V.; Cowan, D.A.; Maggs-Kölling, G.; DiRuggiero, J. Trophic Selective Pressures Organize the Composition of Endolithic Microbial Communities From Global Deserts. Front. Microbiol. 2020, 10, 2952. [Google Scholar] [CrossRef] [Green Version]
- Wierzchos, J.; DiRuggiero, J.; Vítek, P.; Artieda, O.; Souza-Egipsy, V.; Škaloud, P.; Tisza, M.J.; Davila, A.F.; Vílchez, C.; Garbayo, I.; et al. Adaptation Strategies of Endolithic Chlorophototrophs to Survive the Hyperarid and Extreme Solar Radiation Environment of the Atacama Desert. Front. Microbiol. 2015, 6, 934. [Google Scholar] [CrossRef] [Green Version]
- Singh, N.K.; Sonani, R.R.; Prasad Rastogi, R.; Madamwar, D. The Phycobilisomes: An Early Requisite for Efficient Photosynthesis in Cyanobacteria. EXCLI J. 2015, 14, 268–289. [Google Scholar] [CrossRef] [PubMed]
- Cordero, R.R.; Seckmeyer, G.; Damiani, A.; Riechelmann, S.; Rayas, J.; Labbe, F.; Laroze, D. The World’s Highest Levels of Surface UV. Photochem. Photobiol. Sci. 2014, 13, 70–81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Warren-Rhodes, K.A.; McKay, C.P.; Boyle, L.N.; Wing, M.R.; Kiekebusch, E.M.; Cowan, D.A.; Stomeo, F.; Pointing, S.B.; Kaseke, K.F.; Eckardt, F.; et al. Physical Ecology of Hypolithic Communities in the Central Namib Desert: The Role of Fog, Rain, Rock Habitat, and Light. J. Geophys. Res. BioGeosci. 2013, 118, 1451–1460. [Google Scholar] [CrossRef] [Green Version]
- McKay, C.P. Full Solar Spectrum Measurements of Absorption of Light in a Sample of the Beacon Sandstone Containing the Antarctic Cryptoendolithic Microbial Community. Antarct. Sci. 2012, 24, 243–248. [Google Scholar] [CrossRef]
- Croce, R.; Van Amerongen, H. Natural Strategies for Photosynthetic Light Harvesting. Nat. Chem. Biol. 2014, 10, 492–501. [Google Scholar] [CrossRef] [PubMed]
- Ho, M.Y.; Shen, G.; Canniffe, D.P.; Zhao, C.; Bryant, D.A. Light-Dependent Chlorophyll f Synthase Is a Highly Divergent Paralog of PsbA of Photosystem II. Science 2016, 353, aaf9178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ho, M.Y.; Soulier, N.T.; Canniffe, D.P.; Shen, G.; Bryant, D.A. Light Regulation of Pigment and Photosystem Biosynthesis in Cyanobacteria. Curr. Opin. Plant Biol. 2017, 37, 24–33. [Google Scholar] [CrossRef] [Green Version]
- Gan, F.; Shen, G.; Bryant, D.A. Occurrence of Far-Red Light Photoacclimation (FaRLiP) in Diverse Cyanobacteria. Life 2015, 5, 4–24. [Google Scholar] [CrossRef]
- Ho, M.Y.; Gan, F.; Shen, G.; Zhao, C.; Bryant, D.A. Far-Red Light Photoacclimation (FaRLiP) in Synechococcus sp. PCC 7335: I. Regulation of FaRLiP Gene Expression. Photosynth. Res. 2017, 131, 173–186. [Google Scholar] [CrossRef]
- Ho, M.Y.; Gan, F.; Shen, G.; Bryant, D.A. Far-Red Light Photoacclimation (FaRLiP) in Synechococcus sp. PCC 7335. II.Characterization of Phycobiliproteins Produced during Acclimation to Far-Red Light. Photosynth. Res. 2017, 131, 187–202. [Google Scholar] [CrossRef]
- Zhao, C.; Gan, F.; Shen, G.; Bryant, D.A. RfpA, RfpB, and RfpC Are the Master Control Elements of Far-Red Light Photoacclimation (FaRLiP). Front. Microbiol. 2015, 6, 1303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, G.; Canniffe, D.P.; Ho, M.Y.; Kurashov, V.; van der Est, A.; Golbeck, J.H.; Bryant, D.A. Characterization of Chlorophyll f Synthase Heterologously Produced in Synechococcus sp. PCC 7002. Photosynth. Res. 2019, 140, 77–92. [Google Scholar] [CrossRef] [PubMed]
- Murray, J.W. Sequence Variation at the Oxygen-Evolving Centre of Photosystem II: A New Class of “rogue” Cyanobacterial D1 Proteins. Photosynth. Res. 2012, 110, 177–184. [Google Scholar] [CrossRef] [PubMed]
- Cardona, T.; Murray, J.W.; Rutherford, A.W. Origin and Evolution of Water Oxidation before the Last Common Ancestor of the Cyanobacteria. Mol. Biol. Evol. 2015, 32, 1310–1328. [Google Scholar] [CrossRef] [PubMed]
- Antonaru, L.A.; Cardona, T.; Larkum, A.W.D.; Nürnberg, D.J. Global Distribution of a Chlorophyll f Cyanobacterial Marker. ISME J. 2020, 14, 2275–2287. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.C.; Li, Z.K.; Yin, Y.C.; Li, Y.; Jia, Y.; Chen, M.; Qiu, B.S. Widespread Occurrence and Unexpected Diversity of Red-Shifted Chlorophyll Producing Cyanobacteria in Humid Subtropical Forest Ecosystems. Environ. Microbiol. 2019, 21, 1497–1510. [Google Scholar] [CrossRef] [PubMed]
- Ohkubo, S.; Miyashita, H. A Niche for Cyanobacteria Producing Chlorophyll f within a Microbial Mat. ISME J. 2017, 11, 2368–2378. [Google Scholar] [CrossRef]
- Trampe, E.; Kühl, M. Chlorophyll f Distribution and Dynamics in Cyanobacterial Beachrock Biofilms. J. Phycol. 2016, 52, 990–996. [Google Scholar] [CrossRef]
- Mascoli, V.; Bersanini, L.; Croce, R. Far-Red Absorption and Light-Use Efficiency Trade-Offs in Chlorophyll f Photosynthesis. Nat. Plants 2020, 6, 1044–1053. [Google Scholar] [CrossRef]
- Murray, B.; Dailey, M.; Ertekin, E.; DiRuggiero, J. Draft Metagenomes of Endolithic Cyanobacteria and Cohabitants from Hyper-Arid Deserts. Microbiol. Resour. Announc. 2021, 10, e0020621. [Google Scholar] [CrossRef]
- Rippka, R.; Deruelles, J.; Waterbury, J.B. Generic Assignments, Strain Histories and Properties of Pure Cultures of Cyanobacteria. J. Gen. Microbiol. 1979, 111, 1–61. [Google Scholar] [CrossRef] [Green Version]
- Friedmann, I. Chroococcidiopsis kashaii sp. n. and the Genus Chroococcidiopsis (Studies on Cave Algae from Israel III). Osterr. Bot. Z. 1961, 108, 354–367. [Google Scholar] [CrossRef]
- Lebre, P.H.; De Maayer, P.; Cowan, D.A. Xerotolerant Bacteria: Surviving through a Dry Spell. Nat. Rev. Microbiol. 2017, 15, 285–296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knowles, E.J.; Castenholz, R.W. Effect of Exogenous Extracellular Polysaccharides on the Desiccation and Freezing Tolerance of Rock-Inhabiting Phototrophic Microorganisms. FEMS Microbiol. Ecol. 2008, 66, 261–270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, M.; Schliep, M.; Willows, R.D.; Cai, Z.-L.; Neilan, B.A.; Scheer, H. A Red-Shifted Chlorophyll. Science 2010, 329, 1318–1320. [Google Scholar] [CrossRef]
- Canniffe, D.P.; Hunter, C.N. Engineered Biosynthesis of Bacteriochlorophyll b in Rhodobacter sphaeroides. Biochim. Biophys. Acta Bioenerg. 2014, 1837, 1611–1616. [Google Scholar] [CrossRef] [Green Version]
- Bryant, D.A.; Shen, G.; Turner, G.M.; Soulier, N.; Laremore, T.N.; Ho, M.Y. Far-Red Light Allophycocyanin Subunits Play a Role in Chlorophyll d Accumulation in Far-Red Light. Photosynth. Res. 2020, 143, 81–95. [Google Scholar] [CrossRef]
- Ho, M.Y.; Bryant, D.A. Global Transcriptional Profiling of the Cyanobacterium Chlorogloeopsis fritschii PCC 9212 in Far-Red Light: Insights into the Regulation of Chlorophyll d Synthesis. Front. Microbiol. 2019, 10, 465. [Google Scholar] [CrossRef]
- Kurashov, V.; Ho, M.Y.; Shen, G.; Piedl, K.; Laremore, T.N.; Bryant, D.A.; Golbeck, J.H. Energy Transfer from Chlorophyll f to the Trapping Center in Naturally Occurring and Engineered Photosystem I Complexes. Photosynth. Res. 2019, 141, 151–163. [Google Scholar] [CrossRef]
- Nürnberg, D.J.; Morton, J.; Santabarbara, S.; Telfer, A.; Joliot, P.; Antonaru, L.A.; Ruban, A.V.; Cardona, T.; Krausz, E.; Boussac, A.; et al. Photochemistry beyond the Red Limit in Chlorophyll f–Containing Photosystems. Science 2018, 360, 1210–1213. [Google Scholar] [CrossRef] [Green Version]
- Gisriel, C.J.; Shen, G.; Ho, M.Y.; Kurashov, V.; Flesher, D.A.; Wang, J.; Armstrong, W.H.; Golbeck, J.H.; Gunner, M.R.; Vinyard, D.J.; et al. Structure of a Monomeric Photosystem II Core Complex from a Cyanobacterium Acclimated to Far-Red Light Reveals the Functions of Chlorophylls d and f. J. Biol. Chem. 2022, 298, 101424. [Google Scholar] [CrossRef] [PubMed]
- Gisriel, C.J.; Flesher, D.A.; Shen, G.; Wang, J.; Ho, M.Y.; Brudvig, G.W.; Bryant, D.A. Structure of a Photosystem I-Ferredoxin Complex from a Marine Cyanobacterium Provides Insights into Far-Red Light Photoacclimation. J. Biol. Chem. 2022, 298, 101408. [Google Scholar] [CrossRef] [PubMed]
- Airs, R.L.; Temperton, B.; Sambles, C.; Farnham, G.; Skill, S.C.; Llewellyn, C.A. Chlorophyll f and Chlorophyll d Are Produced in the Cyanobacterium Chlorogloeopsis fritschii When Cultured under Natural Light and Near-Infrared Radiation. FEBS Lett. 2014, 588, 3770–3777. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hunter, C.N.; Daldal, F.; Thurnauer, M.C.; Beatty, J.T. (Eds.) The Purple Phototrophic Bacteria. In Advances in Photosynthesis and Respiration; Springer: Berlin, Germany, 2008; Volume 28. [Google Scholar]
- Soulier, N.; Laremore, T.N.; Bryant, D.A. Characterization of Cyanobacterial Allophycocyanins Absorbing Far-Red Light. Photosynth. Res. 2020, 145, 189–207. [Google Scholar] [CrossRef]
- Gwizdala, M.; Lebre, P.H.; Maggs-Kölling, G.; Marais, E.; Cowan, D.A.; Krüger, T.P.J. Sub-Lithic Photosynthesis in Hot Desert Habitats. Environ. Microbiol. 2021, 23, 3867–3880. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Murray, B.; Ertekin, E.; Dailey, M.; Soulier, N.T.; Shen, G.; Bryant, D.A.; Perez-Fernandez, C.; DiRuggiero, J. Adaptation of Cyanobacteria to the Endolithic Light Spectrum in Hyper-Arid Deserts. Microorganisms 2022, 10, 1198. https://doi.org/10.3390/microorganisms10061198
Murray B, Ertekin E, Dailey M, Soulier NT, Shen G, Bryant DA, Perez-Fernandez C, DiRuggiero J. Adaptation of Cyanobacteria to the Endolithic Light Spectrum in Hyper-Arid Deserts. Microorganisms. 2022; 10(6):1198. https://doi.org/10.3390/microorganisms10061198
Chicago/Turabian StyleMurray, Bayleigh, Emine Ertekin, Micah Dailey, Nathan T. Soulier, Gaozhong Shen, Donald A. Bryant, Cesar Perez-Fernandez, and Jocelyne DiRuggiero. 2022. "Adaptation of Cyanobacteria to the Endolithic Light Spectrum in Hyper-Arid Deserts" Microorganisms 10, no. 6: 1198. https://doi.org/10.3390/microorganisms10061198