
Inactivation of Bacillus subtilis by Curcumin-Mediated Photodynamic Technology through Inducing Oxidative Stress Response


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strain and Culture Conditions
2.2. Preparation of Photosensitizer
2.3. Light-Emitting Diode (LED) System
2.4. PDT Inactivation
2.5. ROS Measurement
2.6. Oxidative Enzyme Detection
2.7. Propidium Iodide (PI) Staining
2.8. Scanning Electron Microscopy (SEM)
2.9. Transmission Electron Microscope (TEM)
2.10. RNA Extraction, Reverse Transcription, and RT-qPCR
2.11. Statistics
3. Results
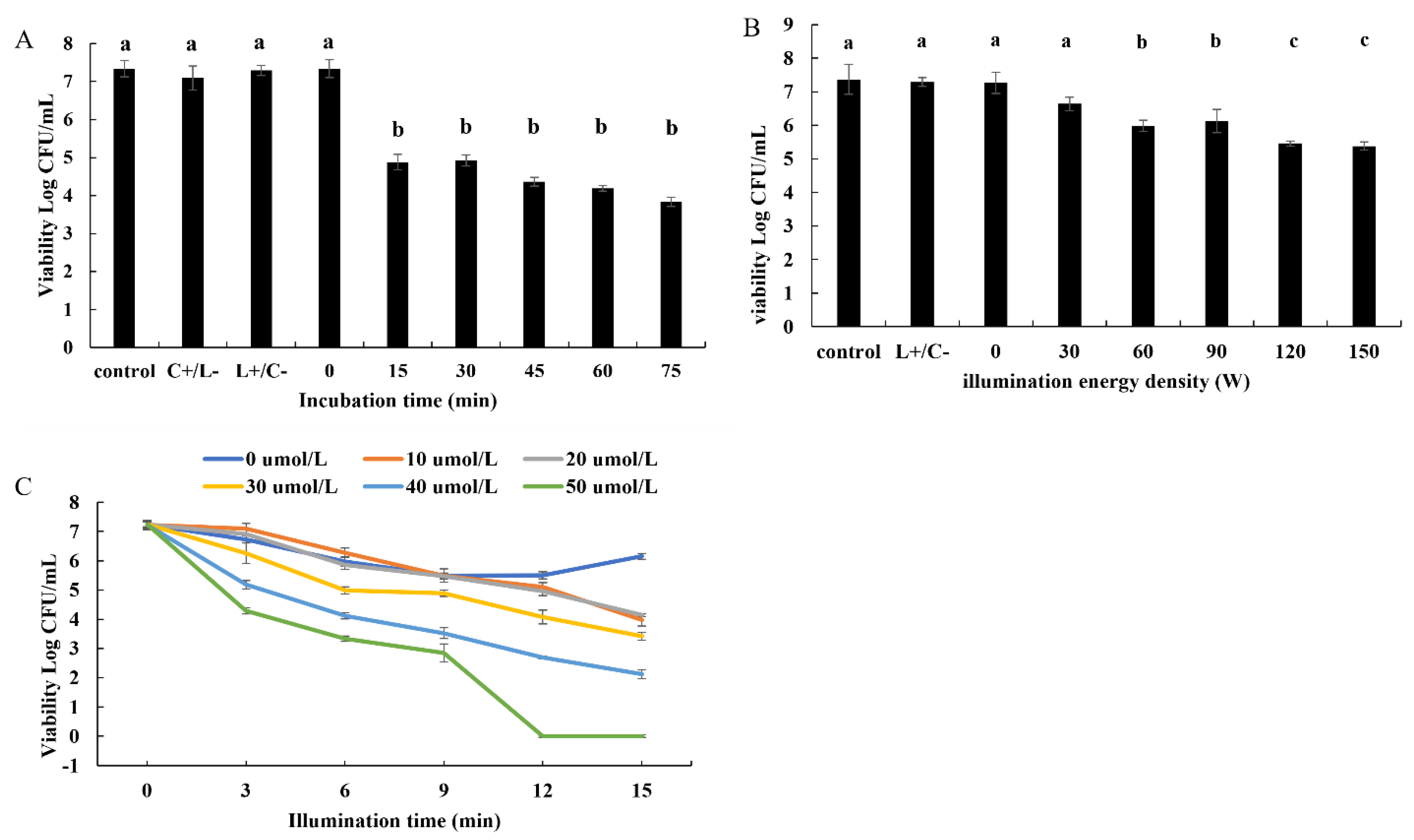
3.1. Antibacterial Efficacy of Photosensitizer-Mediated PDT against B. subtilis
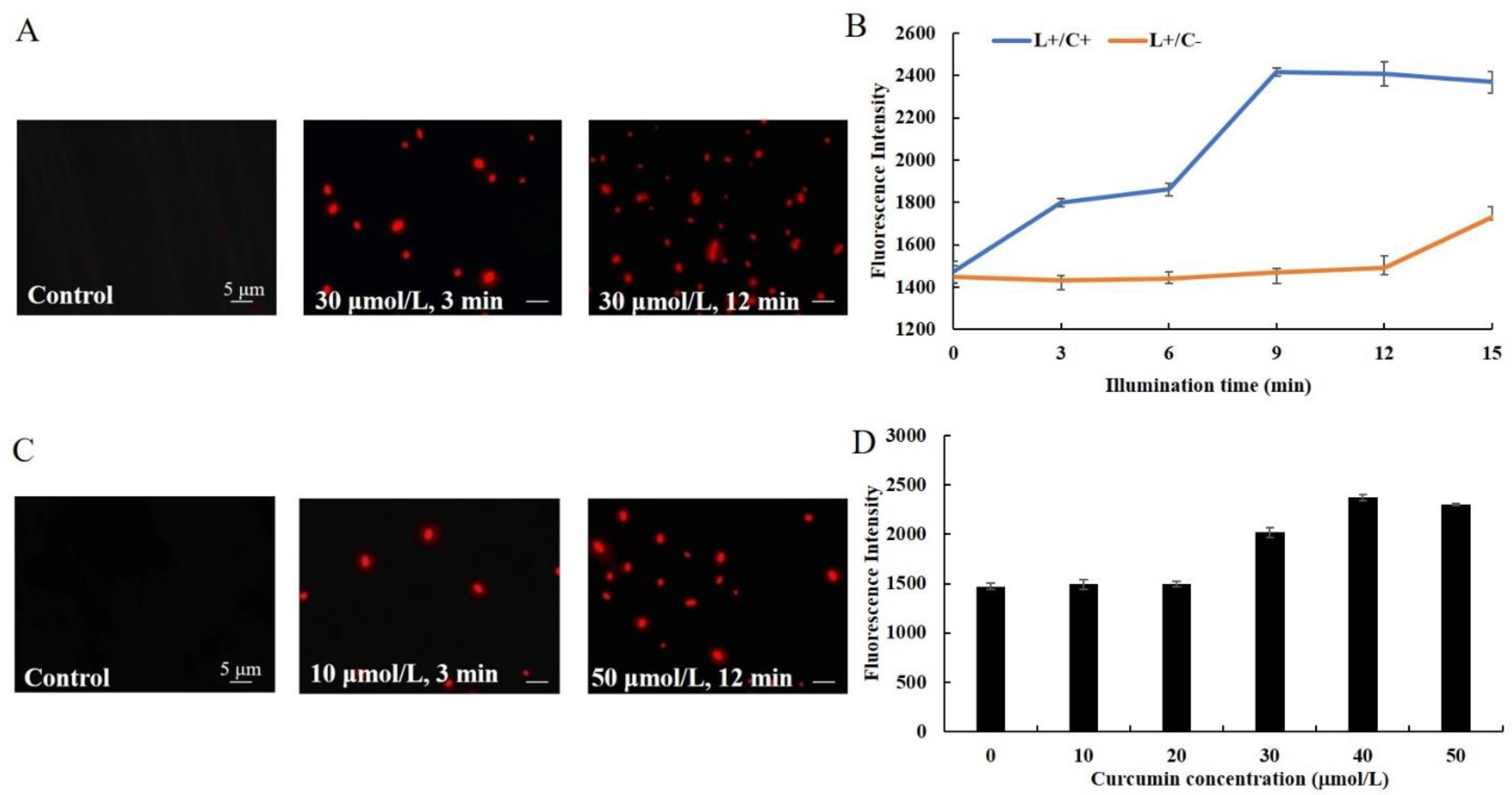
3.2. Effect of Cur-PDT on Cell Membrane Permeability
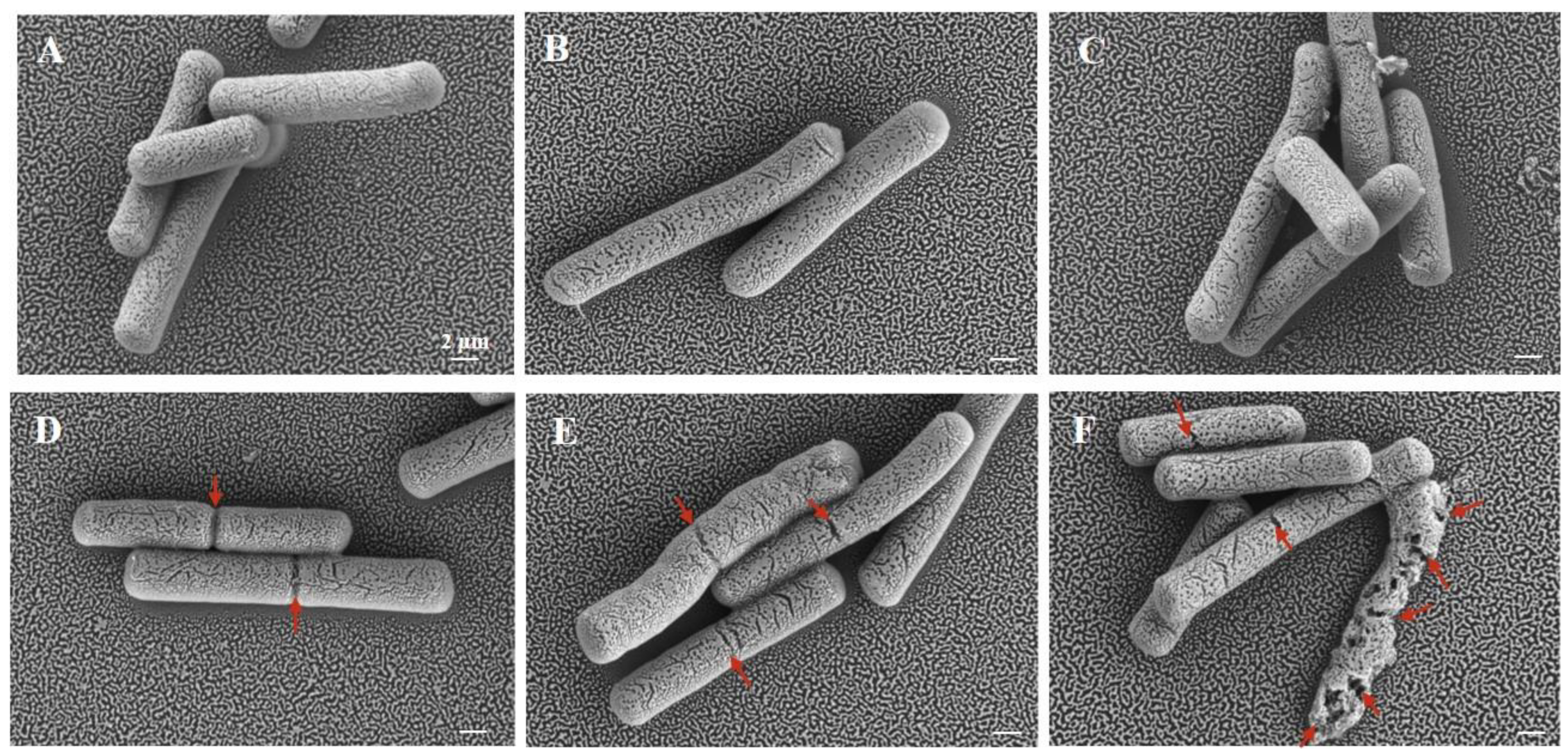
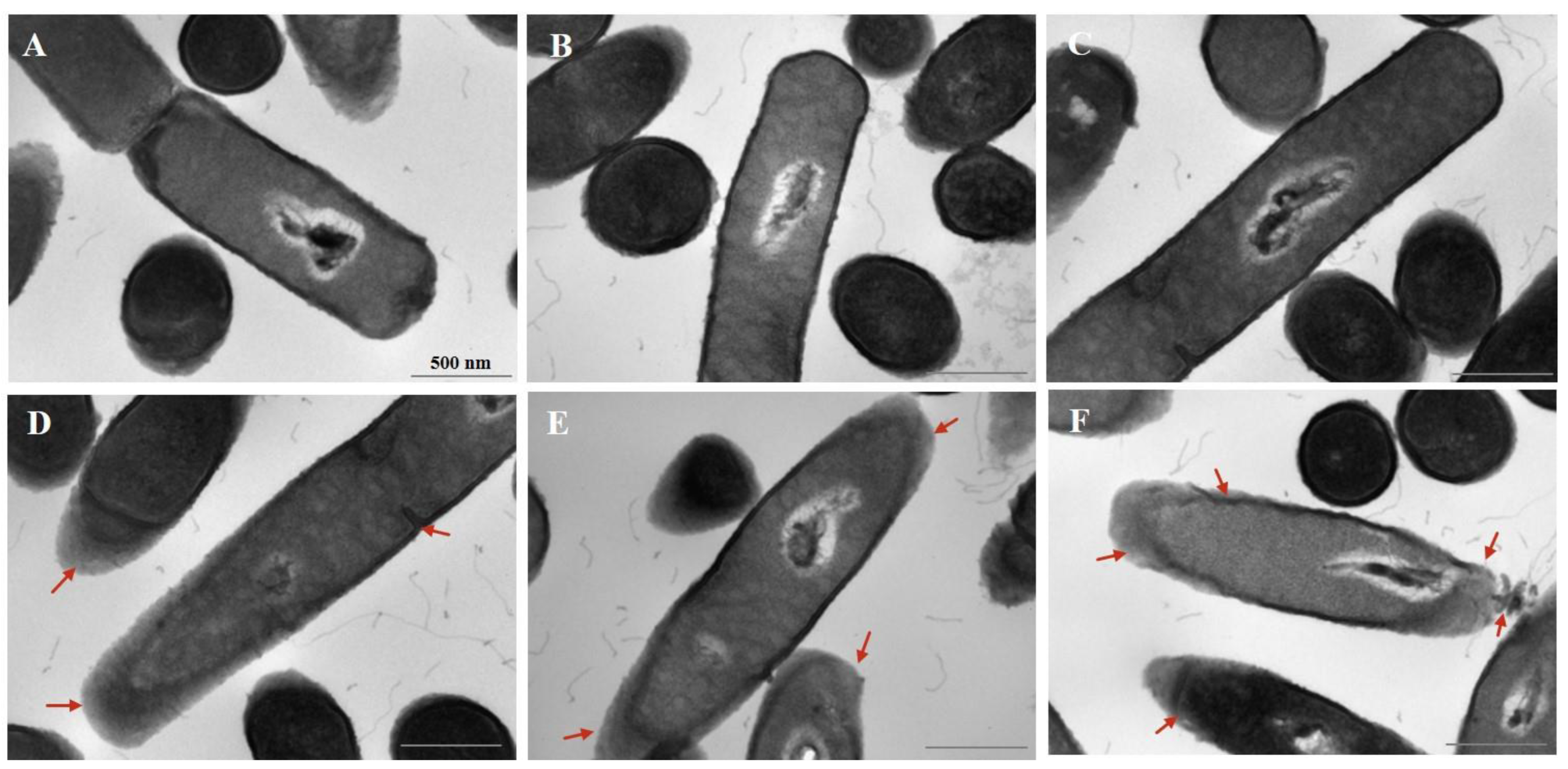
3.3. Effect of Cur-PDT on Cell Morphology
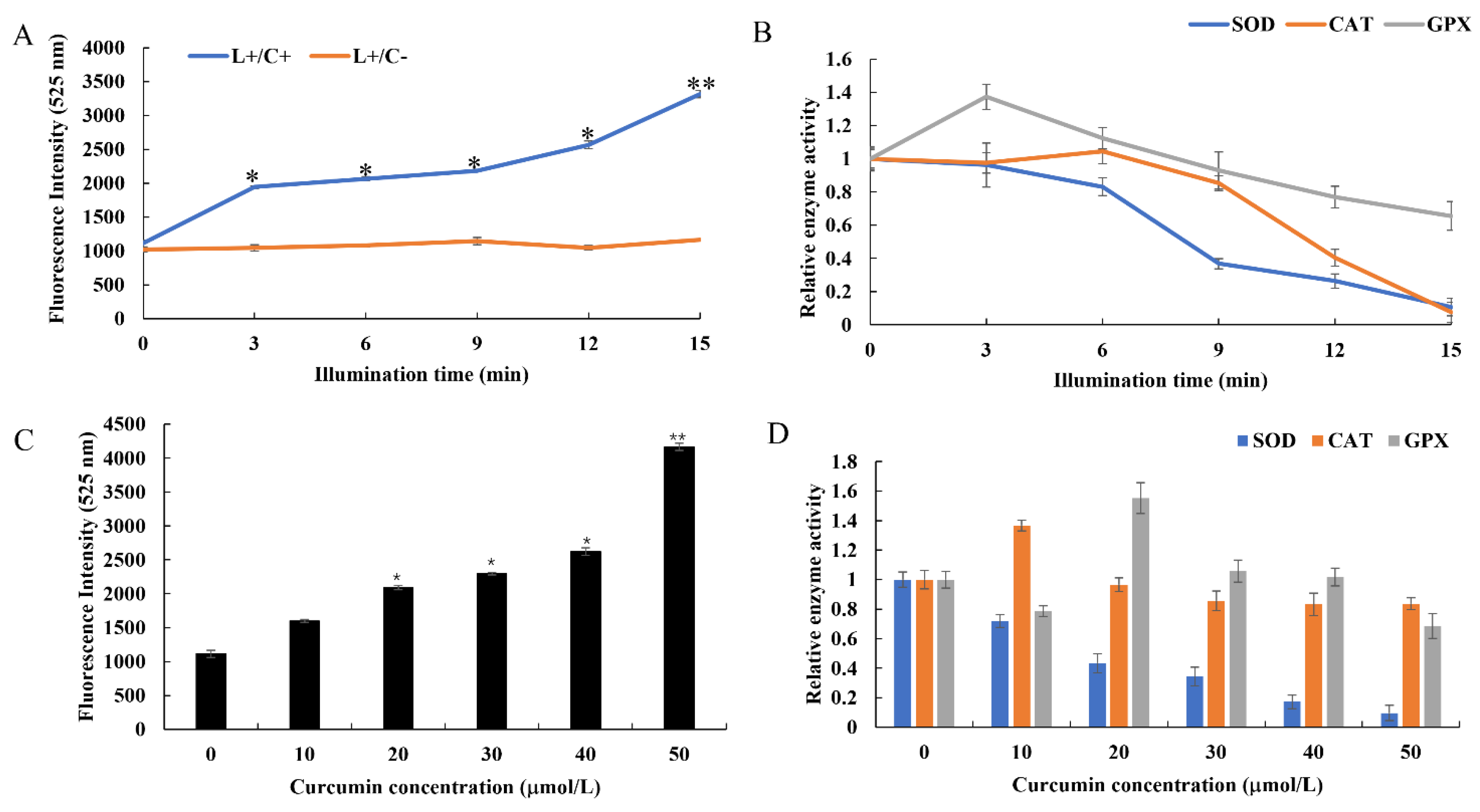
3.4. Changes of Redox State in Cells after Curcumin-PDT Treatment
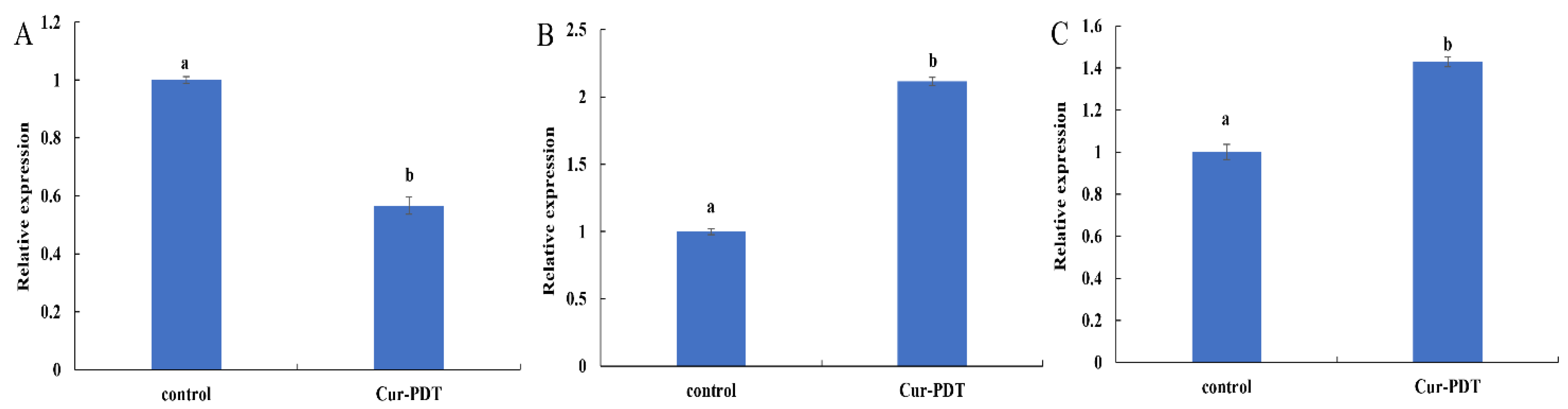
3.5. Regulatory Effects of Cur-PDT on Membrane Structure and DNA Repair System at the Transcriptional Level
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Picart-Palmade, L.; Cunault, C.; Chevalier-Lucia, D.; Belleville, M.-P.; Marchesseau, S. Potentialities and Limits of Some Non-thermal Technologies to Improve Sustainability of Food Processing. Front. Nutr. 2019, 5, 130. [Google Scholar] [CrossRef] [PubMed]
- Morata, A.; Escott, C.; Loira, I.; López, C.; Palomero, F.; González, C. Emerging Non-Thermal Technologies for the Extraction of Grape Anthocyanins. Antioxidants 2021, 10, 1863. [Google Scholar] [CrossRef] [PubMed]
- Chacha, J.S.; Zhang, L.; Ofoedu, C.E.; Suleiman, R.A.; Dotto, J.M.; Roobab, U.; Agunbiade, A.O.; Duguma, H.T.; Mkojera, B.T.; Hossaini, S.M.; et al. Revisiting Non-Thermal Food Processing and Preservation Methods-Action Mechanisms, Pros and Cons: A Technological Update (2016–2021). Foods 2021, 10, 1430. [Google Scholar] [CrossRef] [PubMed]
- Assadi, I.; Guesmi, A.; Baaloudj, O.; Zeghioud, H.; Elfalleh, W.; Benhammadi, N.; Khezami, L.; Assadi, A.A. Review on inactivation of airborne viruses using non-thermal plasma technologies: From MS2 to coronavirus. Environ. Sci. Pollut. Res. Int. 2022, 29, 4880–4892. [Google Scholar] [CrossRef] [PubMed]
- Sakudo, A.; Yagyu, Y.; Onodera, T. Disinfection and Sterilization Using Plasma Technology: Fundamentals and Future Perspectives for Biological Applications. Int. J. Mol. Sci. 2019, 20, 5216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Youf, R.; Müller, M.; Balasini, A.; Thétiot, F.; Müller, M.; Hascoët, A.; Jonas, U.; Schönherr, H.; Lemercier, G.; Montier, T.; et al. Antimicrobial Photodynamic Therapy: Latest Developments with a Focus on Combinatory Strategies. Pharmaceutics 2021, 13, 1995. [Google Scholar] [CrossRef]
- Benov, L. Photodynamic therapy: Current status and future directions. Med. Princ. Pract. 2015, 24 (Suppl. S1), 14–28. [Google Scholar] [CrossRef] [PubMed]
- Dai, T.; Huang, Y.-Y.; Hamblin, M.R. Photodynamic therapy for localized infections—State of the art. Photodiagn. Photodyn. Ther. 2009, 6, 170–188. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.; Zhou, F.; Lai, D.; Zhang, Y.; Hu, J.; Lin, S. Curcumin-mediated sono/photodynamic treatment preserved the quality of shrimp surimi and influenced its microbial community changes during refrigerated storage. Ultrason. Sonochem. 2021, 78, 105715. [Google Scholar] [CrossRef]
- Aponiene, K.; Paskeviciute, E.; Reklaitis, I.; Luksiene, Z. Reduction of microbial contamination of fruits and vegetables by hypericin-based photosensitization: Comparison with other emerging antimicrobial treatments. J. Food Eng. 2015, 144, 29–35. [Google Scholar] [CrossRef]
- Yu, J.; Zhang, F.; Zhang, J.; Han, Q.; Song, L.; Meng, X. Effect of photodynamic treatments on quality and antioxidant properties of fresh-cut potatoes. Food Chem. 2021, 362, 130224. [Google Scholar] [CrossRef] [PubMed]
- Hyun, J.E.; Lee, S.Y.; Hyun, J.E.; Lee, S.Y. Antibacterial effect and mechanisms of action of 460–470 nm light-emitting diode against Listeria monocytogenes and Pseudomonas fluorescens on the surface of packaged sliced cheese. Food Microbiol. 2020, 86, 103314. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Matthews, K.R. Effects of the photosensitizer curcumin in inactivating foodborne pathogens on chicken skin. Food Control 2020, 109, 106959. [Google Scholar] [CrossRef]
- Lin, Y.A.-O.; Hu, J.A.-O.; Li, S.; Hamzah, S.S.; Jiang, H.; Zhou, A.; Zeng, S.; Lin, S. Curcumin-Based Photodynamic Sterilization for Preservation of Fresh-Cut Hami Melon. Molecules 2019, 24, 2374. [Google Scholar] [CrossRef] [Green Version]
- Setlow, P. Spore Resistance Properties. Microbiol. Spectr. 2014, 2, 4099–4100. [Google Scholar] [CrossRef] [Green Version]
- Setlow, P. Spores of Bacillus subtilis: Their resistance to and killing by radiation, heat and chemicals. J. Appl. Microbiol. 2006, 101, 514–525. [Google Scholar] [CrossRef]
- Liu, X.-Y.; Hu, Q.; Xu, F.; Ding, S.-Y.; Zhu, K. Characterization of Bacillus cereus in Dairy Products in China. Toxins 2020, 12, 454. [Google Scholar] [CrossRef]
- Kang, J.-W.; Hong, H.-N.; Kang, D.-H. Application of a Krypton-Chlorine Excilamp To Control Alicyclobacillus acidoterrestris Spores in Apple Juice and Identification of Its Sporicidal Mechanism. Appl. Environ. Microbiol. 2020, 86, e00159-20. [Google Scholar] [CrossRef]
- Wood, J.P.; Adrion, A.C. Review of Decontamination Techniques for the Inactivation of Bacillus anthracis and Other Spore-Forming Bacteria Associated with Building or Outdoor Materials. Environ. Sci. Technol. 2019, 53, 4045–4062. [Google Scholar] [CrossRef]
- Priyadarsini, K.I. The chemistry of curcumin: From extraction to therapeutic agent. Molecules 2014, 19, 20091–20112. [Google Scholar] [CrossRef] [Green Version]
- Chen, B.; Huang, J.; Li, H.; Zeng, Q.-H.; Wang, J.J.; Liu, H.; Pan, Y.; Zhao, Y. Eradication of planktonic Vibrio parahaemolyticus and its sessile biofilm by curcumin-mediated photodynamic inactivation. Food Control 2020, 113, 107181. [Google Scholar] [CrossRef]
- Temba, B.A.; Fletcher, M.T.; Fox, G.P.; Harvey, J.J.W.; Sultanbawa, Y. Inactivation of Aspergillus flavus spores by curcumin-mediated photosensitization. Food Control 2016, 59, 708–713. [Google Scholar] [CrossRef]
- Chen, B.; Huang, J.; Liu, Y.; Liu, H.; Zhao, Y.; Wang, J.J. Effects of the curcumin-mediated photodynamic inactivation on the quality of cooked oysters with Vibrio parahaemolyticus during storage at different temperature. Int. J. Food Microbiol. 2021, 345, 109152. [Google Scholar] [CrossRef] [PubMed]
- Qi, S.; Guo, L.; Yan, S.; Lee, R.J.; Yu, S.; Chen, S. Hypocrellin A-based photodynamic action induces apoptosis in A549 cells through ROS-mediated mitochondrial signaling pathway. Acta Pharm. Sin. B 2019, 9, 279–293. [Google Scholar] [CrossRef]
- Kawano, A.; Yamasaki, R.; Sakakura, T.; Takatsuji, Y.; Haruyama, T.; Yoshioka, Y.; Ariyoshi, W. Reactive Oxygen Species Penetrate Persister Cell Membranes of Escherichia coli for Effective Cell Killing. Front. Cell Infect. Microbiol. 2020, 10, 496. [Google Scholar] [CrossRef]
- St Denis, T.G.; Huang, L.; Dai, T.; Hamblin, M.R. Analysis of the bacterial heat shock response to photodynamic therapy-mediated oxidative stress. Photochem. Photobiol. 2011, 87, 707–713. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Hu, Y.; Jingwen, C. Impact of Dimethyl Phthalate on the Activities of Oxidative Stress Enzymes and Growth of Typical Bacteria. Asian J. Ecotoxicol. 2015, 3, 298–303. [Google Scholar]
- Ghate, V.; Kumar, A.; Zhou, W.; Yuk, H.-G. Irradiance and Temperature Influence the Bactericidal Effect of 460-Nanometer Light-Emitting Diodes on Salmonella in Orange Juice. J. Food Prot. 2016, 79, 553–560. [Google Scholar] [CrossRef]
- Verma, N.; Srivastava, S.; Malik, R.; Goyal, P.; Pandey, J. Inhibition and disintegration of Bacillus subtilis biofilm with small molecule inhibitors identified through virtual screening for targeting TasA (28-261), the major protein component of ECM. J. Biomol. Struct. Dyn. 2022, 31, 1–17. [Google Scholar] [CrossRef]
- Romero, D.; Aguilar, C.; Losick, R.; Kolter, R. Amyloid fibers provide structural integrity to Bacillus subtilis biofilms. Proc. Natl. Acad. Sci. USA 2010, 107, 2230–2234. [Google Scholar] [CrossRef] [Green Version]
- de Melo, W.C.M.A.; Avci, P.; de Oliveira, M.N.; Gupta, A.; Vecchio, D.; Sadasivam, M.; Chandran, R.; Huang, Y.-Y.; Yin, R.; Perussi, L.R.; et al. Photodynamic inactivation of biofilm: Taking a lightly colored approach to stubborn infection. Expert Rev. Anti-Infect. Ther. 2013, 11, 669–693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, Y.; Cheng, Q.; Yang, H.; Li, H.; Gong, N.; Liu, D.; Wu, J.; Lei, X. Effects of ALA-PDT on biofilm structure, virulence factor secretion, and QS in Pseudomonas aeruginosa. Photodiagn. Photodyn. Ther. 2018, 24, 88–94. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zeng, R.; Duan, Z.; Xu, H.; Wu, Q.; Chen, Q.; Lin, T.; Li, M. Effect of 5-aminolevulinic Acid Photodynamics Therapy on Biofilm of Propionibacterium Acnes. Zhongguo Yi Xue Ke Xue Yuan Xue Bao 2020, 42, 283–288. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, I.P.; Pinto, J.G.; Souza, B.M.N.; Minan, A.G.; Ferreira-Strixino, J. Antimicrobial photodynamic therapy with curcumin on methicillin-resistant Staphylococcus aureus biofilm. Photodiagn. Photodyn. Ther. 2022, 37, 102729. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Jiang, C.; Figueiro Longo, J.P.; Azevedo, R.B.; Zhang, H.; Muehlmann, L.A. An updated overview on the development of new photosensitizers for anticancer photodynamic therapy. Acta Pharm. Sin. B 2018, 8, 137–146. [Google Scholar] [CrossRef]
- Choi, S.S.; Oh, H.Y.; Kim, E.J.; Lee, H.K.; Kim, H.K.; Choi, H.H.; Kim, S.W.; Chae, H.S. In Vitro Bactericidal Effects of Photodynamic Therapy Combined with Four Tetracyclines against Clostridioides difficile KCTC5009 in Planktonic Cultures. Pathogens 2020, 9, 279. [Google Scholar] [CrossRef]
- Chen, R.; Song, Y.; Zhen, Y.; Yao, L.A.-O.; Shi, Y.; Cui, Y.A.-O.; Li, S.A.-O. 5-Aminolevulinic acid-mediated photodynamic therapy has effective antifungal activity against Sporothrix globosa in vitro. Mycoses 2020, 63, 1311–1320. [Google Scholar] [CrossRef]
- Rangasamy, S.; Ju, H.; Um, S.; Oh, D.C.; Song, J.M. Mitochondria and DNA Targeting of 5,10,15,20-Tetrakis(7-sulfonatobenzo[b]thiophene) Porphyrin-Induced Photodynamic Therapy via Intrinsic and Extrinsic Apoptotic Cell Death. J. Med. Chem. 2015, 58, 6864–6874. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dong, L.; Qin, J.; Tai, L.; Mou, K.; Liao, X.; Chen, F.; Hu, X. Inactivation of Bacillus subtilis by Curcumin-Mediated Photodynamic Technology through Inducing Oxidative Stress Response. Microorganisms 2022, 10, 802. https://doi.org/10.3390/microorganisms10040802
Dong L, Qin J, Tai L, Mou K, Liao X, Chen F, Hu X. Inactivation of Bacillus subtilis by Curcumin-Mediated Photodynamic Technology through Inducing Oxidative Stress Response. Microorganisms. 2022; 10(4):802. https://doi.org/10.3390/microorganisms10040802
Chicago/Turabian StyleDong, Li, Jianran Qin, Luyang Tai, Kangyi Mou, Xiaojun Liao, Fang Chen, and Xiaosong Hu. 2022. "Inactivation of Bacillus subtilis by Curcumin-Mediated Photodynamic Technology through Inducing Oxidative Stress Response" Microorganisms 10, no. 4: 802. https://doi.org/10.3390/microorganisms10040802