
Screening for Biofilm-Stimulating Factors in the Freshwater Planctomycete Planctopirus limnophila to Improve Sessile Growth in a Chemically Defined Medium



Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strain and Media
2.2. Cultivation of P. limnophila in Microtiter Plates
2.3. Cultivation of P. limnophila in the Bioreactor
2.4. MTT Assay
2.5. Crystal Violet Staining
2.6. Measurement of OD600
3. Results
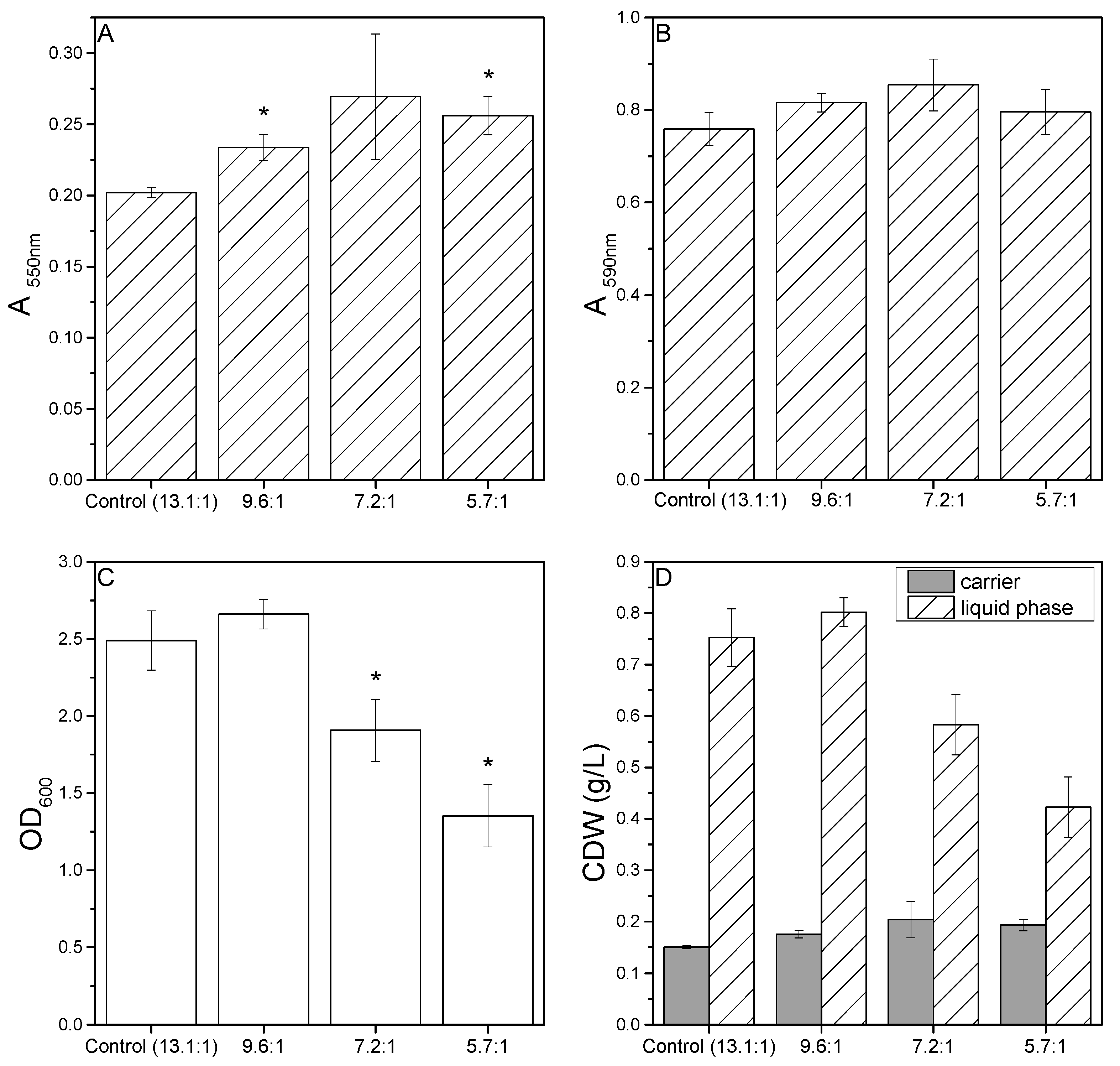
3.1. Effect of C:N Ratio on Biofilm Formation in Microtiter Plates
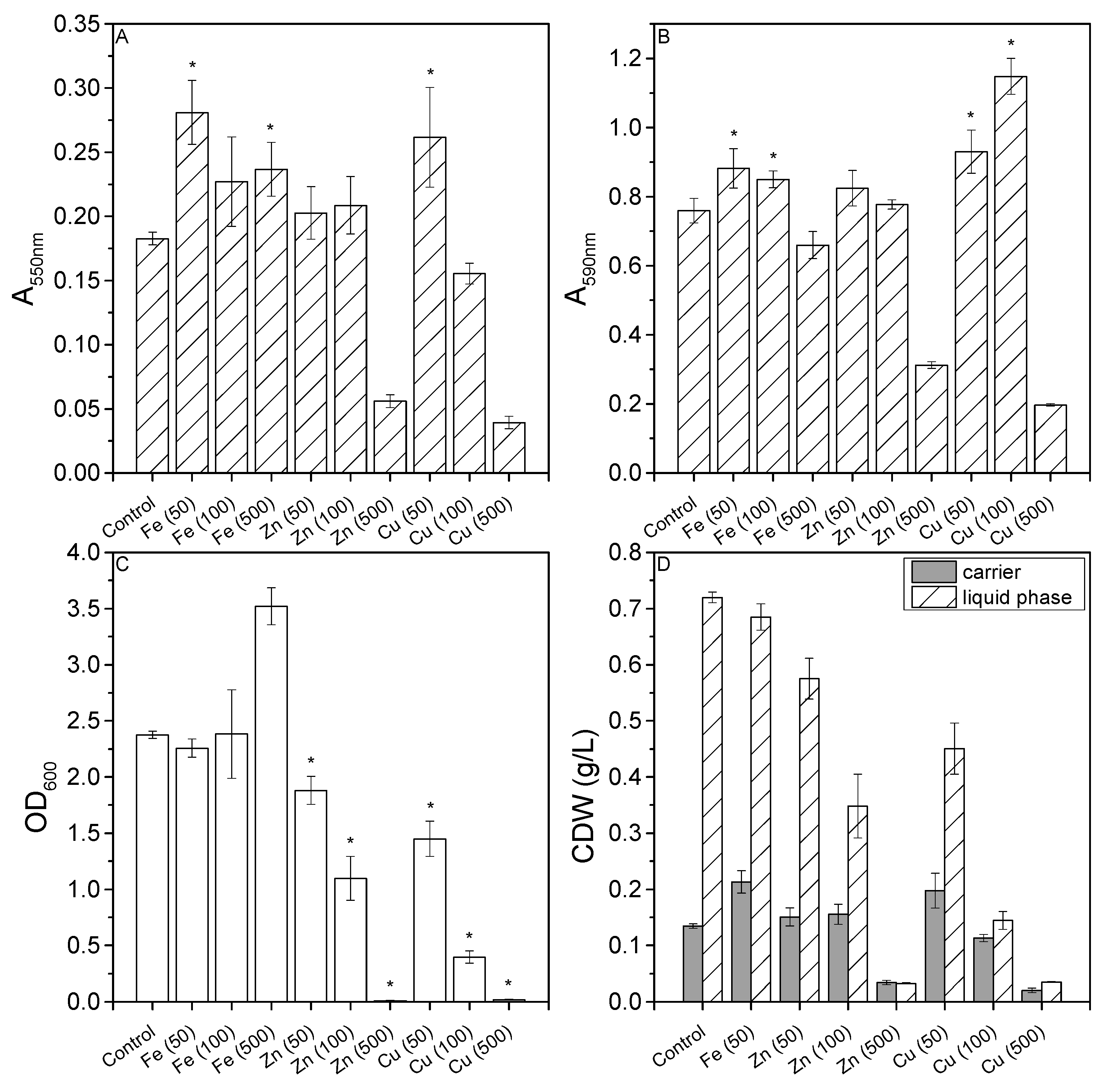
3.2. Effect of Metal Ions on Biofilm Formation in Microtiter Plates
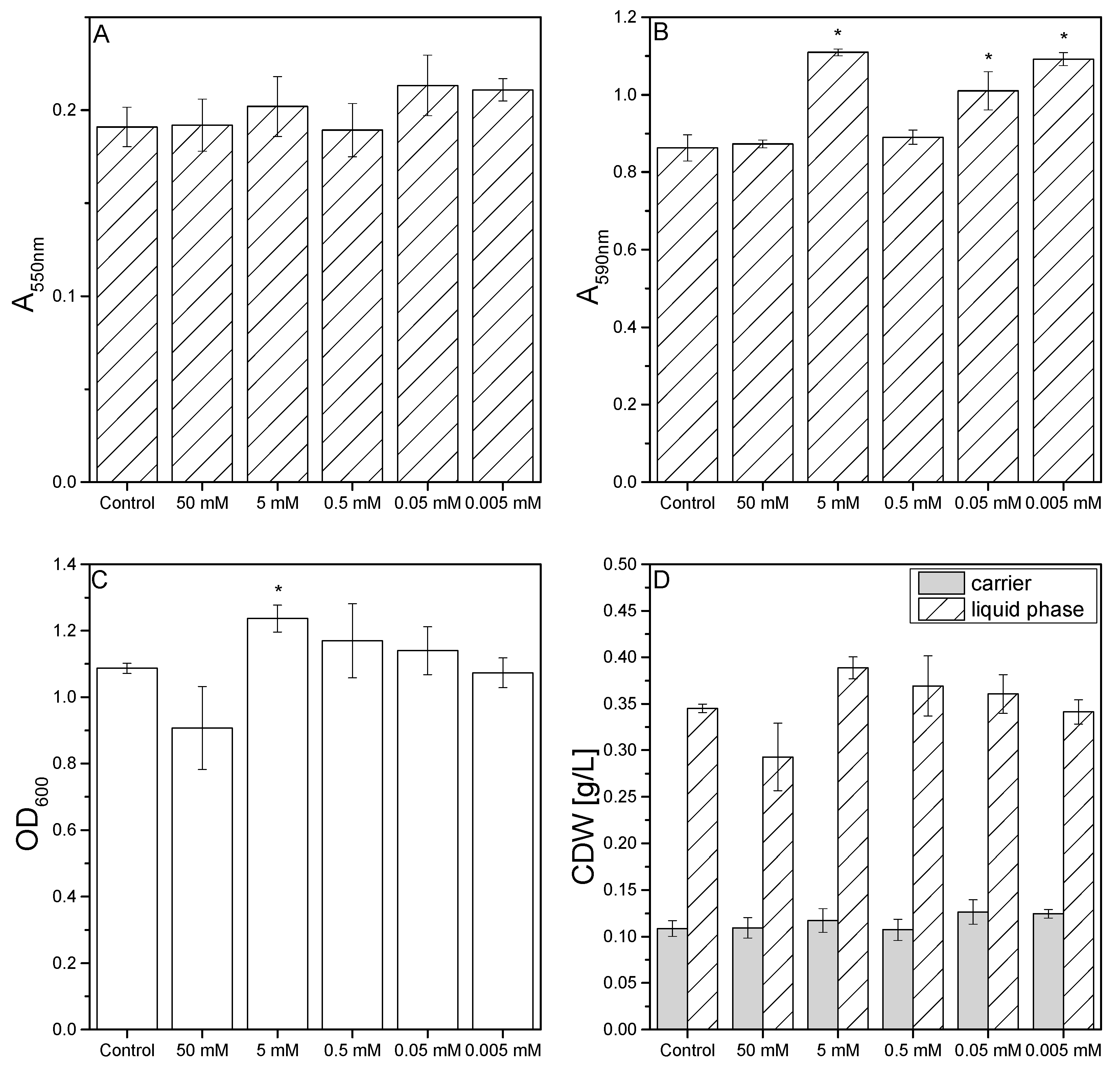
3.3. Effect of Oxidative Stress on Biofilm Formation in Microtiter Plates
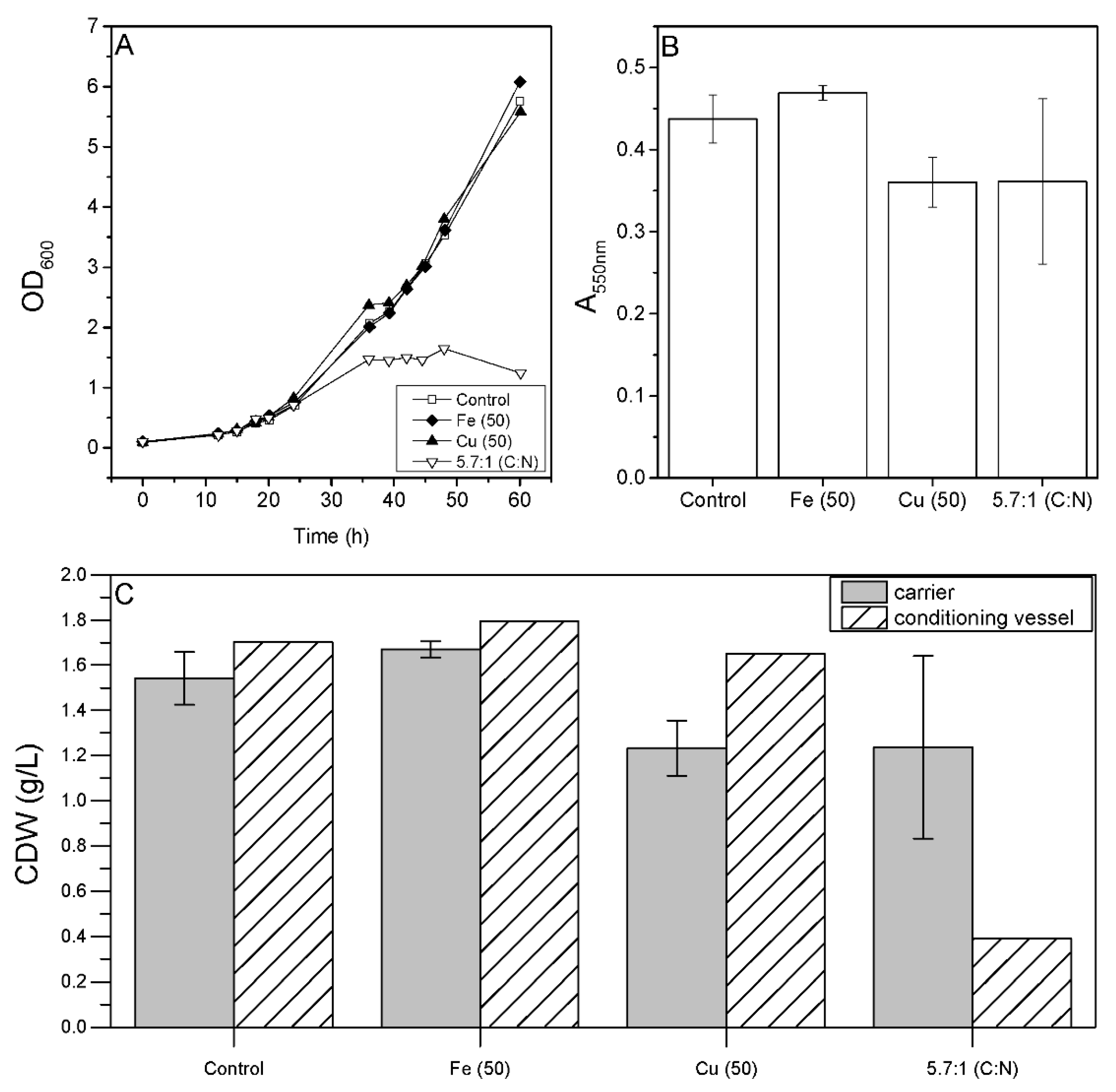
3.4. Effect of Cultivation Parameters on Biofilm Formation in Bioreactors
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jeske, O.; Jogler, M.; Petersen, J.; Sikorski, J.; Jogler, C. From genome mining to phenotypic microarrays: Planctomycetes as source for novel bioactive molecules. Antonie Leeuwenhoek 2013, 104, 551–567. [Google Scholar] [CrossRef] [PubMed]
- Kohn, T.; Heuer, A.; Jogler, M.; Vollmers, J.; Boedeker, C.; Bunk, B.; Rast, P.; Borchert, D.; Glöckner, I.; Freese, H.M.; et al. Fuerstia marisgermanicae gen. nov., sp. nov., an Unusual Member of the Phylum Planctomycetes from the German Wadden Sea. Front. Microbiol. 2016, 7, 2079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Graça, A.P.; Calisto, R.; Lage, O.M. Planctomycetes as Novel Source of Bioactive Molecules. Front. Microbiol. 2016, 7, 1241. [Google Scholar] [CrossRef]
- Peeters, S.H.; Wiegand, S.; Kallscheuer, N.; Jogler, M.; Heuer, A.; Jetten, M.S.M.; Rast, P.; Boedeker, C.; Rohde, M.; Jogler, C. Three marine strains constitute the novel genus and species Crateriforma conspicua in the phylum Planctomycetes. Antonie Leeuwenhoek 2020, 113, 1797–1809. [Google Scholar] [CrossRef] [PubMed]
- Kallscheuer, N.; Jeske, O.; Sandargo, B.; Boedeker, C.; Wiegand, S.; Bartling, P.; Jogler, M.; Rohde, M.; Petersen, J.; Medema, M.H.; et al. The planctomycete Stieleria maiorica Mal15T employs stieleriacines to alter the species composition in marine biofilms. Commun. Biol. 2020, 3, 303. [Google Scholar] [CrossRef] [PubMed]
- Wagner, M.; Horn, M. The Planctomycetes, Verrucomicrobia, Chlamydiae and sister phyla comprise a superphylum with biotechnological and medical relevance. Curr. Opin. Biotechnol. 2006, 17, 241–249. [Google Scholar] [CrossRef]
- Vigneron, A.; Alsop, E.B.; Cruaud, P.; Philibert, G.; King, B.; Baksmaty, L.; Lavallée, D.; Lomans, B.P.; Kyrpides, N.C.; Head, I.M.; et al. Comparative metagenomics of hydrocarbon and methane seeps of the Gulf of Mexico. Sci. Rep. 2017, 7, 16015. [Google Scholar] [CrossRef] [Green Version]
- Peeters, S.H.; van Niftrik, L. Trending topics and open questions in anaerobic ammonium oxidation. Curr. Opin. Chem. Biol. 2019, 49, 45–52. [Google Scholar] [CrossRef]
- Jogler, C.; Waldmann, J.; Huang, X.; Jogler, M.; Glöckner, F.O.; Mascher, T.; Kolter, R. Identification of proteins likely to be involved in morphogenesis, cell division, and signal transduction in Planctomycetes by comparative genomics. J. Bacteriol. 2012, 194, 6419–6430. [Google Scholar] [CrossRef] [Green Version]
- Wiegand, S.; Jogler, M.; Boedeker, C.; Pinto, D.; Vollmers, J.; Rivas-Marín, E.; Kohn, T.; Peeters, S.H.; Heuer, A.; Rast, P.; et al. Cultivation and functional characterization of 79 planctomycetes uncovers their unique biology. Nat. Microbiol. 2020, 5, 126–140. [Google Scholar] [CrossRef]
- Jeske, O.; Schüler, M.; Schumann, P.; Schneider, A.; Boedeker, C.; Jogler, M.; Bollschweiler, D.; Rohde, M.; Mayer, C.; Engelhardt, H.; et al. Planctomycetes do possess a peptidoglycan cell wall. Nat. Commun. 2015, 6, 7116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jogler, C.; Glöckner, F.O.; Kolter, R. Characterization of Planctomyces limnophilus and development of genetic tools for its manipulation establish it as a model species for the phylum Planctomycetes. Appl. Environ. Microbiol. 2011, 77, 5826–5829. [Google Scholar] [CrossRef] [Green Version]
- Buckley, D.H.; Huangyutitham, V.; Nelson, T.A.; Rumberger, A.; Thies, J.E. Diversity of Planctomycetes in soil in relation to soil history and environmental heterogeneity. Appl. Environ. Microbiol. 2006, 72, 4522–4531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiegand, S.; Jogler, M.; Jogler, C. On the maverick Planctomycetes. FEMS Microbiol. Rev. 2018, 42, 739–760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bondoso, J.; Balagué, V.; Gasol, J.M.; Lage, O.M. Community composition of the Planctomycetes associated with different macroalgae. FEMS Microbiol. Ecol. 2014, 88, 445–456. [Google Scholar] [CrossRef] [Green Version]
- Lage, O.M.; Bondoso, J. Planctomycetes diversity associated with macroalgae. FEMS Microbiol. Ecol. 2011, 78, 366–375. [Google Scholar] [CrossRef] [Green Version]
- Lage, O.M.; Bondoso, J. Planctomycetes and macroalgae, a striking association. Front. Microbiol. 2014, 5, 267. [Google Scholar] [CrossRef] [Green Version]
- Bengtsson, M.M.; Øvreås, L. Planctomycetes dominate biofilms on surfaces of the kelp Laminaria hyperborea. BMC Microbiol. 2010, 10, 261. [Google Scholar] [CrossRef] [Green Version]
- Fuerst, J.A.; Webb, R.I. Membrane-bounded nucleoid in the eubacterium Gemmata obscuriglobus. Proc. Natl. Acad. Sci. USA 1991, 88, 8184–8188. [Google Scholar] [CrossRef] [Green Version]
- Lonhienne, T.G.A.; Sagulenko, E.; Webb, R.I.; Lee, K.-C.; Franke, J.; Devos, D.P.; Nouwens, A.; Carroll, B.J.; Fuerst, J.A. Endocytosis-like protein uptake in the bacterium Gemmata obscuriglobus. Proc. Natl. Acad. Sci. USA 2010, 107, 12883–12888. [Google Scholar] [CrossRef] [Green Version]
- Lindsay, M.R.; Webb, R.I.; Fuerst, J.A. Pirellulosomes: A new type of membrane-bounded cell compartment in planctomycete bacteria of the genus Pirellula. Microbiology 1997, 143, 739–748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- König, E.; Schlesner, H.; Hirsch, P. Cell wall studies on budding bacteria of the Planctomyces/Pasteuria group and on a Prosthecomicrobium sp. Arch. Microbiol. 1984, 138, 200–205. [Google Scholar] [CrossRef]
- Boedeker, C.; Schüler, M.; Reintjes, G.; Jeske, O.; van Teeseling, M.C.F.; Jogler, M.; Rast, P.; Borchert, D.; Devos, D.P.; Kucklick, M.; et al. Determining the bacterial cell biology of Planctomycetes. Nat. Commun. 2017, 8, 14853. [Google Scholar] [CrossRef]
- Lage, O.M.; Bondoso, J.; Lobo-da-Cunha, A. Insights into the ultrastructural morphology of novel Planctomycetes. Antonie Leeuwenhoek 2013, 104, 467–476. [Google Scholar] [CrossRef] [PubMed]
- Panter, F.; Garcia, R.; Thewes, A.; Zaburannyi, N.; Bunk, B.; Overmann, J.; Gutierrez, M.V.; Krug, D.; Müller, R. Production of a Dibrominated Aromatic Secondary Metabolite by a Planctomycete Implies Complex Interaction with a Macroalgal Host. ACS Chem. Biol. 2019, 14, 2713–2719. [Google Scholar] [CrossRef]
- Jeske, O.; Surup, F.; Ketteniß, M.; Rast, P.; Förster, B.; Jogler, M.; Wink, J.; Jogler, C. Developing Techniques for the Utilization of Planctomycetes As Producers of Bioactive Molecules. Front. Microbiol. 2016, 7, 1242. [Google Scholar] [CrossRef] [Green Version]
- Rumbo-Feal, S.; Gómez, M.J.; Gayoso, C.; Álvarez-Fraga, L.; Cabral, M.P.; Aransay, A.M.; Rodríguez-Ezpeleta, N.; Fullaondo, A.; Valle, J.; Tomás, M.; et al. Whole transcriptome analysis of Acinetobacter baumannii assessed by RNA-sequencing reveals different mRNA expression profiles in biofilm compared to planktonic cells. PLoS ONE 2013, 8, e72968. [Google Scholar] [CrossRef] [Green Version]
- Shemesh, M.; Tam, A.; Steinberg, D. Differential gene expression profiling of Streptococcus mutans cultured under biofilm and planktonic conditions. Microbiology 2007, 153, 1307–1317. [Google Scholar] [CrossRef] [Green Version]
- Beloin, C.; Valle, J.; Latour-Lambert, P.; Faure, P.; Kzreminski, M.; Balestrino, D.; Haagensen, J.A.J.; Molin, S.; Prensier, G.; Arbeille, B.; et al. Global impact of mature biofilm lifestyle on Escherichia coli K-12 gene expression. Mol. Microbiol. 2004, 51, 659–674. [Google Scholar] [CrossRef]
- Landini, P. Cross-talk mechanisms in biofilm formation and responses to environmental and physiological stress in Escherichia coli. Res. Microbiol. 2009, 160, 259–266. [Google Scholar] [CrossRef]
- Wilson, G.S.; Raftos, D.A.; Nair, S.V. Antimicrobial activity of surface attached marine bacteria in biofilms. Microbiol. Res. 2011, 166, 437–448. [Google Scholar] [CrossRef]
- Rieusset, L.; Rey, M.; Muller, D.; Vacheron, J.; Gerin, F.; Dubost, A.; Comte, G.; Prigent-Combaret, C. Secondary metabolites from plant-associated Pseudomonas are overproduced in biofilm. Microb. Biotechnol. 2020, 13, 1562–1580. [Google Scholar] [CrossRef] [PubMed]
- Bengtsson, M.M.; Sjøtun, K.; Øvreås, L. Seasonal dynamics of bacterial biofilms on the kelp Laminaria hyperborea. Aquat. Microb. Ecol. 2010, 60, 71–83. [Google Scholar] [CrossRef] [Green Version]
- Pizzetti, I.; Gobet, A.; Fuchs, B.M.; Amann, R.; Fazi, S. Abundance and diversity of Planctomycetes in a Tyrrhenian coastal system of central Italy. Aquat. Microb. Ecol. 2011, 65, 129–141. [Google Scholar] [CrossRef] [Green Version]
- Flemming, H.-C.; Wingender, J. The biofilm matrix. Nat. Rev. Microbiol. 2010, 8, 623–633. [Google Scholar] [CrossRef] [PubMed]
- Donlan, R.M. Biofilms: Microbial life on surfaces. Emerging Infect. Dis. 2002, 8, 881–890. [Google Scholar] [CrossRef] [PubMed]
- De La Fuente-Núñez, C.; Reffuveille, F.; Fernández, L.; Hancock, R.E.W. Bacterial biofilm development as a multicellular adaptation: Antibiotic resistance and new therapeutic strategies. Curr. Opin. Microbiol. 2013, 16, 580–589. [Google Scholar] [CrossRef]
- Aguilera, A.; Souza-Egipsy, V.; Martín-Uriz, P.S.; Amils, R. Extracellular matrix assembly in extreme acidic eukaryotic biofilms and their possible implications in heavy metal adsorption. Aquat. Toxicol. 2008, 88, 257–266. [Google Scholar] [CrossRef]
- Roberson, E.B.; Firestone, M.K. Relationship between Desiccation and Exopolysaccharide Production in a Soil Pseudomonas sp. AEM 1992, 58, 1284–1291. [Google Scholar] [CrossRef] [Green Version]
- Sivadon, P.; Barnier, C.; Urios, L.; Grimaud, R. Biofilm formation as a microbial strategy to assimilate particulate substrates. Environ. Microbiol. Rep. 2019, 11, 749–764. [Google Scholar] [CrossRef]
- Whiteley, M.; Diggle, S.P.; Greenberg, E.P. Progress in and promise of bacterial quorum sensing research. Nature 2017, 551, 313–320. [Google Scholar] [CrossRef] [PubMed]
- Shrout, J.D.; Chopp, D.L.; Just, C.L.; Hentzer, M.; Givskov, M.; Parsek, M.R. The impact of quorum sensing and swarming motility on Pseudomonas aeruginosa biofilm formation is nutritionally conditional. Mol. Microbiol. 2006, 62, 1264–1277. [Google Scholar] [CrossRef] [PubMed]
- O’Toole, G.; Kaplan, H.B.; Kolter, R. Biofilm formation as microbial development. Annu. Rev. Microbiol. 2000, 54, 49–79. [Google Scholar] [CrossRef] [PubMed]
- de Kievit, T.R. Quorum sensing in Pseudomonas aeruginosa biofilms. Environ. Microbiol. 2009, 11, 279–288. [Google Scholar] [CrossRef]
- Kruppa, O.C.; Gerlach, D.; Fan, R.; Czermak, P. Development of a chemically defined medium for Planctopirus limnophila to increase biomass production. Electron. J. Biotechnol. 2021. [Google Scholar] [CrossRef]
- Riss, T.L.; Moravec, R.A.; Niles, A.L.; Duellman, S.; Benink, H.A.; Worzella, T.J.; Minor, L. Assay Guidance Manual: Cell Viability Assays; Eli Lilly & Company and The National Center for Advancing Translational Sciences: Bethesda, MD, USA, 2004. [PubMed]
- O’Toole, G.A.; Pratt, L.A.; Watnick, P.I.; Newman, D.K.; Weaver, V.B.; Kolter, R. Genetic approaches to study of biofilms. Methods Enzymol. 1999, 310, 91–109. [Google Scholar] [CrossRef]
- Frank, C.S.; Langhammer, P.; Fuchs, B.M.; Harder, J. Ammonium and attachment of Rhodopirellula baltica. Arch. Microbiol. 2011, 193, 365–372. [Google Scholar] [CrossRef]
- Geier, H.; Mostowy, S.; Cangelosi, G.A.; Behr, M.A.; Ford, T.E. Autoinducer-2 triggers the oxidative stress response in Mycobacterium avium, leading to biofilm formation. Appl. Environ. Microbiol. 2008, 74, 1798–1804. [Google Scholar] [CrossRef] [Green Version]
- Kaboré, O.D.; Aghnatios, R.; Godreuil, S.; Drancourt, M. Escherichia coli Culture Filtrate Enhances the Growth of Gemmata spp. Front. Microbiol. 2019, 10, 2552. [Google Scholar] [CrossRef]
- Storesund, J.E.; Øvreås, L. Diversity of Planctomycetes in iron-hydroxide deposits from the Arctic Mid Ocean Ridge (AMOR) and description of Bythopirellula goksoyri gen. nov., sp. nov., a novel Planctomycete from deep sea iron-hydroxide deposits. Antonie Leeuwenhoek 2013, 104, 569–584. [Google Scholar] [CrossRef]
- Banin, E.; Vasil, M.L.; Greenberg, E.P. Iron and Pseudomonas aeruginosa biofilm formation. Proc. Natl. Acad. Sci. USA 2005, 102, 11076–11081. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banin, E.; Brady, K.M.; Greenberg, E.P. Chelator-induced dispersal and killing of Pseudomonas aeruginosa cells in a biofilm. AEM 2006, 72, 2064–2069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Y.; Outten, F.W. IscR controls iron-dependent biofilm formation in Escherichia coli by regulating type I fimbria expression. J. Bacteriol. 2009, 191, 1248–1257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grumbein, S.; Opitz, M.; Lieleg, O. Selected metal ions protect Bacillus subtilis biofilms from erosion. Metallomics 2014, 6, 1441–1450. [Google Scholar] [CrossRef] [Green Version]
- Lin, M.-H.; Shu, J.-C.; Huang, H.-Y.; Cheng, Y.-C. Involvement of iron in biofilm formation by Staphylococcus aureus. PLoS ONE 2012, 7, e34388. [Google Scholar] [CrossRef]
- Mey, A.R.; Craig, S.A.; Payne, S.M. Characterization of Vibrio cholerae RyhB: The RyhB regulon and role of ryhB in biofilm formation. Infect. Immun. 2005, 73, 5706–5719. [Google Scholar] [CrossRef] [Green Version]
- Oh, E.; Andrews, K.J.; Jeon, B. Enhanced Biofilm Formation by Ferrous and Ferric Iron Through Oxidative Stress in Campylobacter jejuni. Front. Microbiol. 2018, 9, 1204. [Google Scholar] [CrossRef] [Green Version]
- Lage, O.M.; Bondoso, J.; Catita, J.A.M. Determination of zeta potential in Planctomycetes and its application in heavy metals toxicity assessment. Arch. Microbiol. 2012, 194, 847–855. [Google Scholar] [CrossRef]
- Murphy, T.F.; Kirkham, C.; Sethi, S.; Lesse, A.J. Expression of a peroxiredoxin-glutaredoxin by Haemophilus influenzae in biofilms and during human respiratory tract infection. FEMS Immunol. Med. Microbiol. 2005, 44, 81–89. [Google Scholar] [CrossRef] [Green Version]
- Wen, Z.T.; Suntharaligham, P.; Cvitkovitch, D.G.; Burne, R.A. Trigger factor in Streptococcus mutans is involved in stress tolerance, competence development, and biofilm formation. Infect. Immun. 2005, 73, 219–225. [Google Scholar] [CrossRef] [Green Version]
- Schembri, M.A.; Hjerrild, L.; Gjermansen, M.; Klemm, P. Differential expression of the Escherichia coli autoaggregation factor antigen 43. J. Bacteriol. 2003, 185, 2236–2242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaboré, O.D.; Godreuil, S.; Drancourt, M. Planctomycetes as Host-Associated Bacteria: A Perspective That Holds Promise for Their Future Isolations, by Mimicking Their Native Environmental Niches in Clinical Microbiology Laboratories. Front. Cell. Infect. Microbiol. 2020, 10, 519301. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Medium | C:N | KNO3 (mM) | NH4Cl (mM) |
|---|---|---|---|
| D1ASO (control) | 13.1:1 | 23.18 | 2.32 |
| Medium 2 | 9.6:1 | 23.18 | 11.6 |
| Medium 3 | 7.2:1 | 23.18 | 23.2 |
| Medium 4 | 5.7:1 | 23.18 | 34.8 |
| ZnSO4 (µM) | CuSO4 (µM) | FeSO4 (µM) | |
|---|---|---|---|
| D1ASO | 6.5 | 0.3 | 15.2 |
| Zn (50) | 56.5 | 0.3 | 15.2 |
| Zn (100) | 106.5 | 0.3 | 15.2 |
| Zn (500) | 506.5 | 0.3 | 15.2 |
| Cu (50) | 6.5 | 50.3 | 15.2 |
| Cu (100) | 6.5 | 100.3 | 15.2 |
| Cu (500) | 6.5 | 500.3 | 15.2 |
| Fe (50) | 6.5 | 0.3 | 65.2 |
| Fe (100) | 6.5 | 0.3 | 115.2 |
| Fe (500) | 6.5 | 0.3 | 515.2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kruppa, O.; Czermak, P. Screening for Biofilm-Stimulating Factors in the Freshwater Planctomycete Planctopirus limnophila to Improve Sessile Growth in a Chemically Defined Medium. Microorganisms 2022, 10, 801. https://doi.org/10.3390/microorganisms10040801
Kruppa O, Czermak P. Screening for Biofilm-Stimulating Factors in the Freshwater Planctomycete Planctopirus limnophila to Improve Sessile Growth in a Chemically Defined Medium. Microorganisms. 2022; 10(4):801. https://doi.org/10.3390/microorganisms10040801
Chicago/Turabian StyleKruppa, Oscar, and Peter Czermak. 2022. "Screening for Biofilm-Stimulating Factors in the Freshwater Planctomycete Planctopirus limnophila to Improve Sessile Growth in a Chemically Defined Medium" Microorganisms 10, no. 4: 801. https://doi.org/10.3390/microorganisms10040801