
The Transcription of Flagella of Enteropathogenic Escherichia coli O127:H6 Is Activated in Response to Environmental and Nutritional Signals

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Construction of the fliC::lacZ Transcriptional Fusion
2.2. Culture Conditions and β-Galactosidase Assays
2.3. Adherence to Epithelial Cells and Detection of Flagella by Immunofluorescence
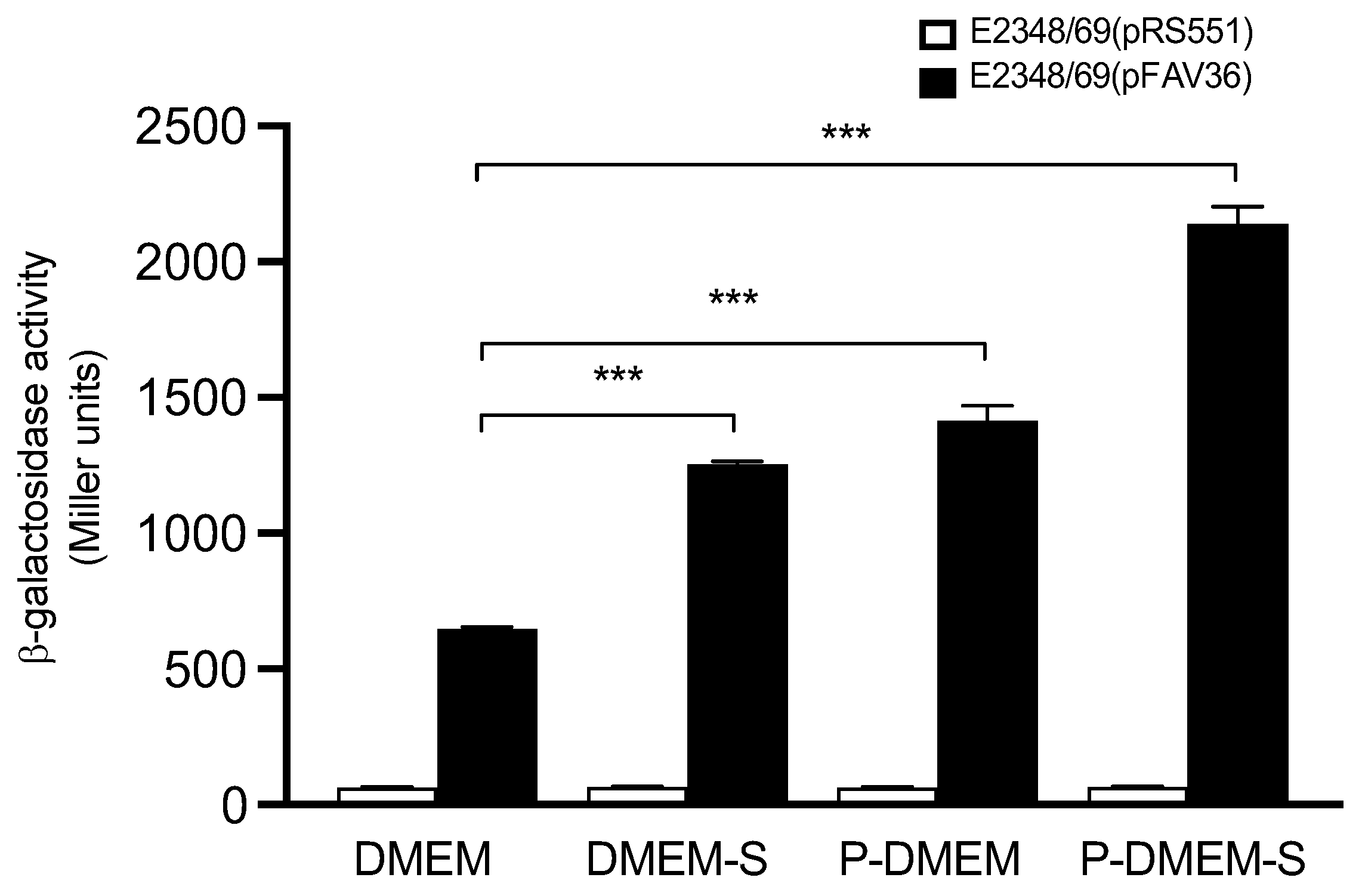
2.4. Pre-Conditioned Medium
2.5. Motility Assays
2.6. Statistical Analysis
3. Results
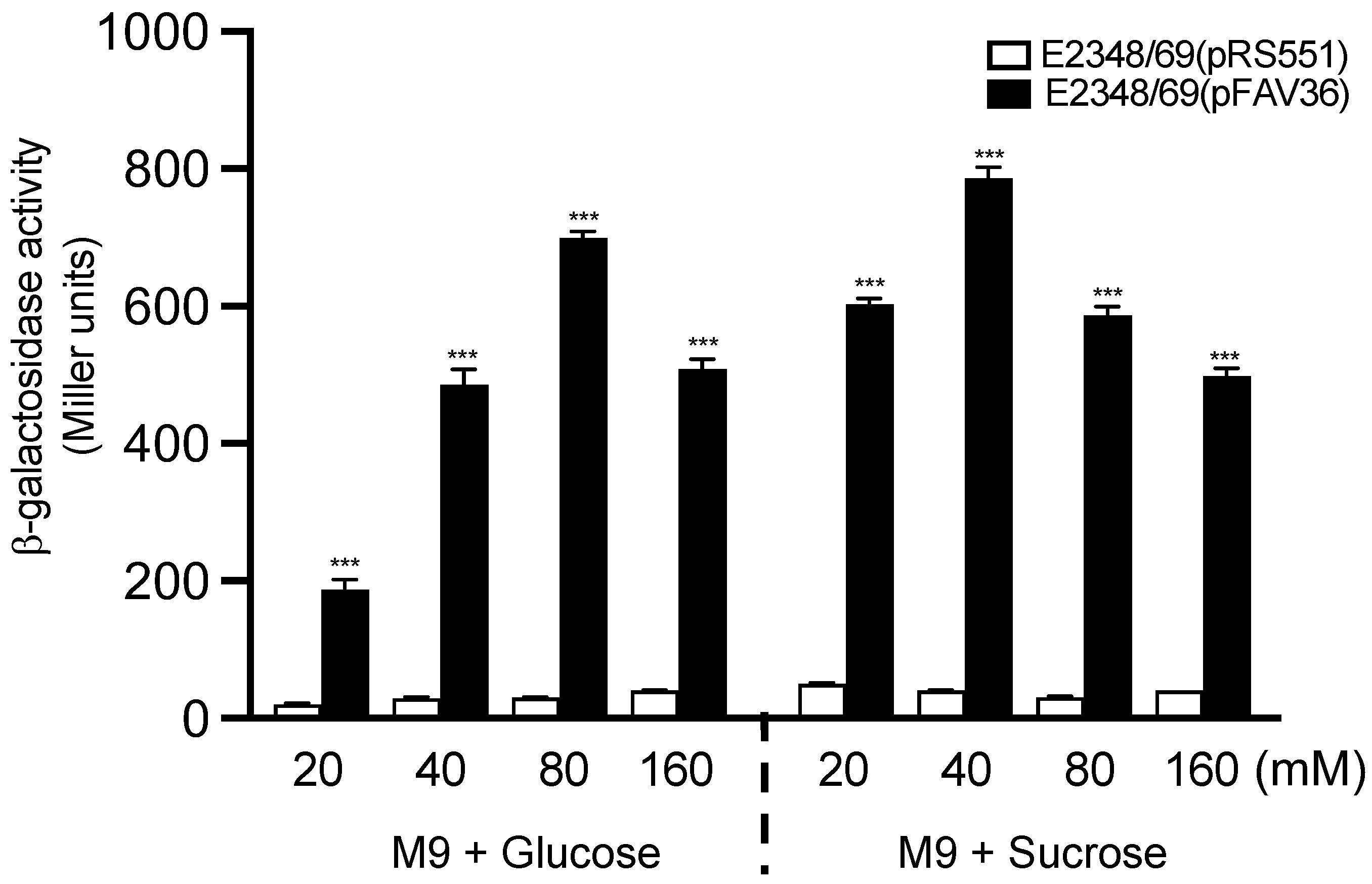
3.1. Glucose Activates fliC Expression in EPEC
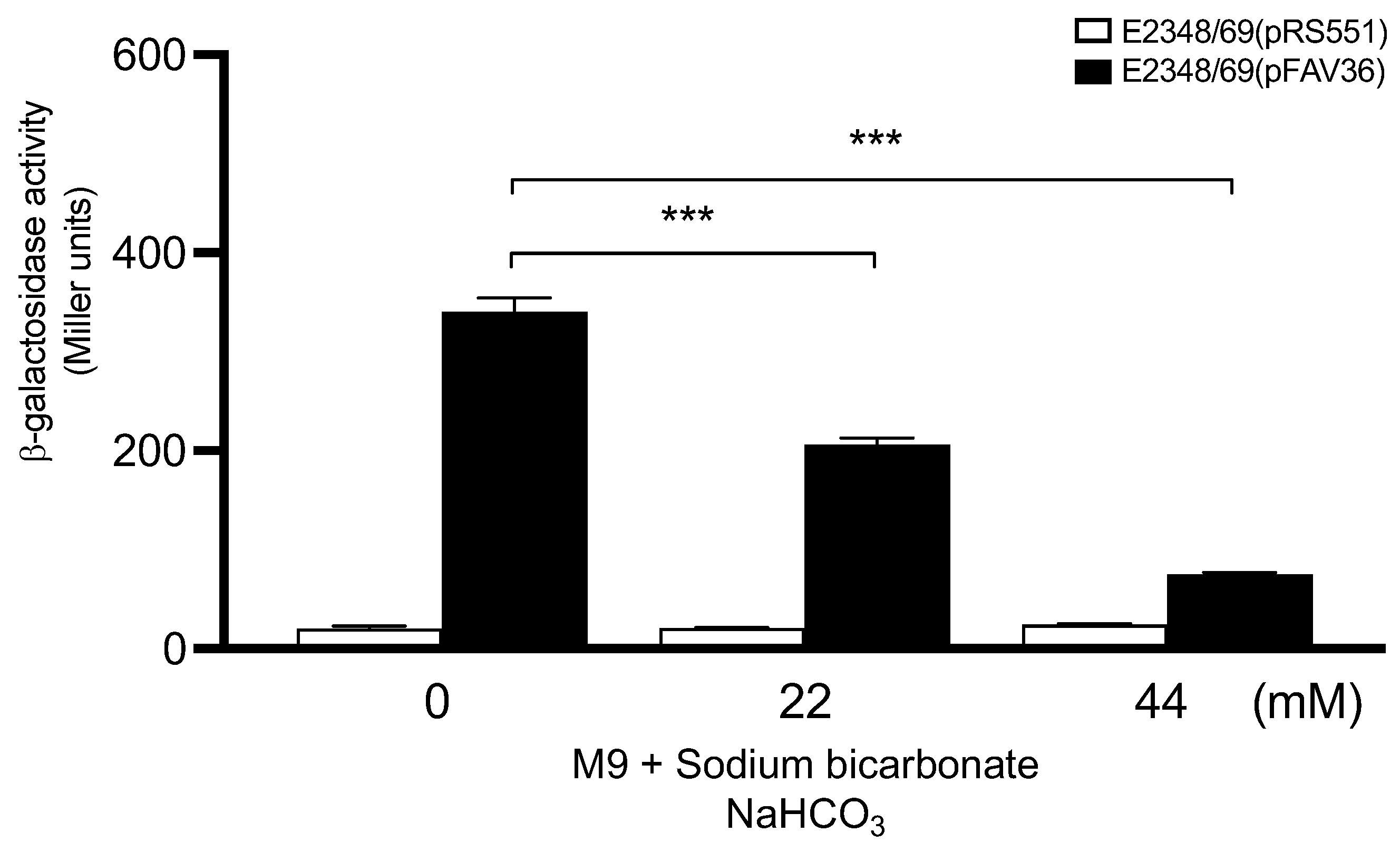
3.2. Role of Sodium Bicarbonate in Flagella Expression
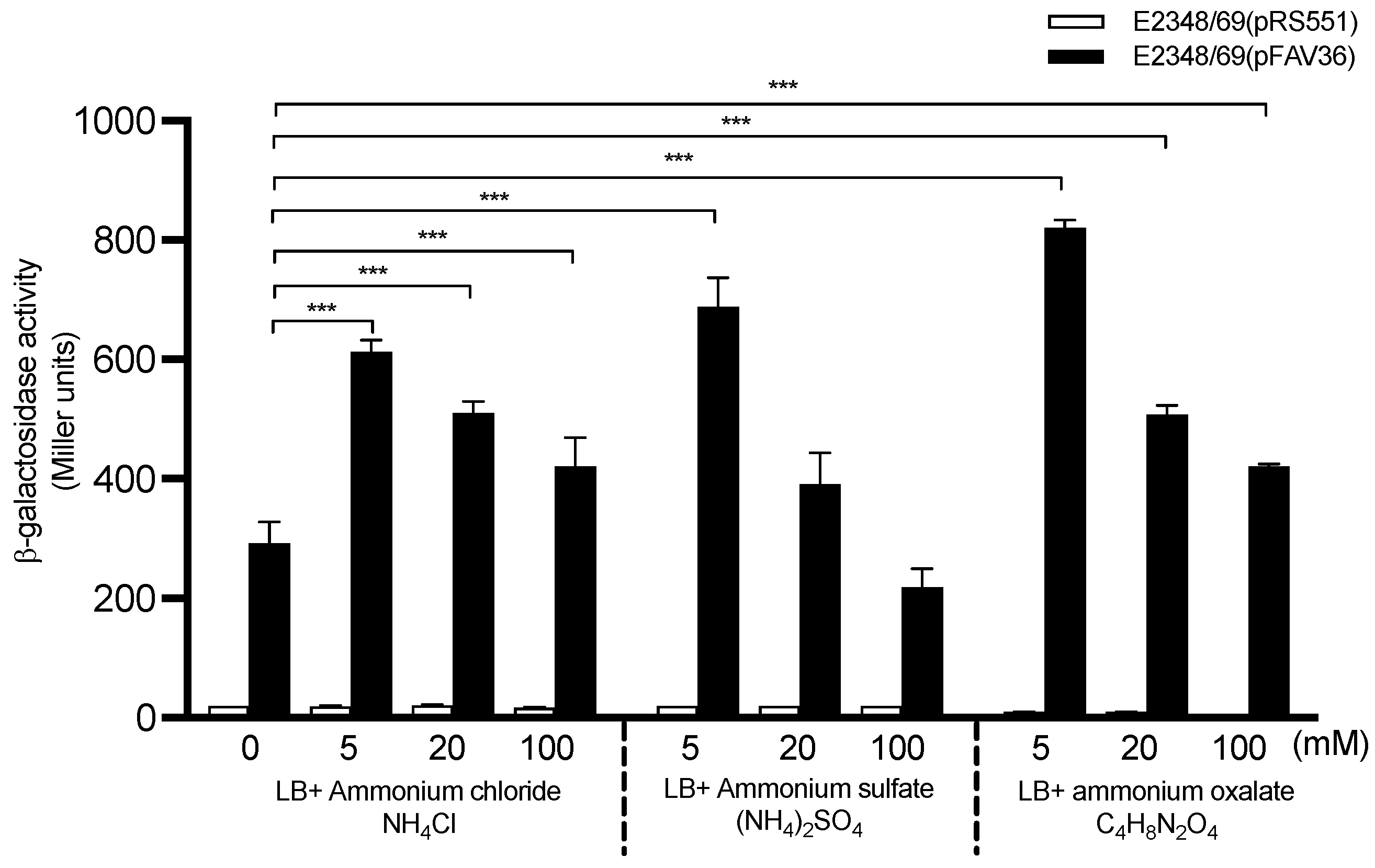
3.3. Modulation of fliC Expression in EPEC by Ammonium
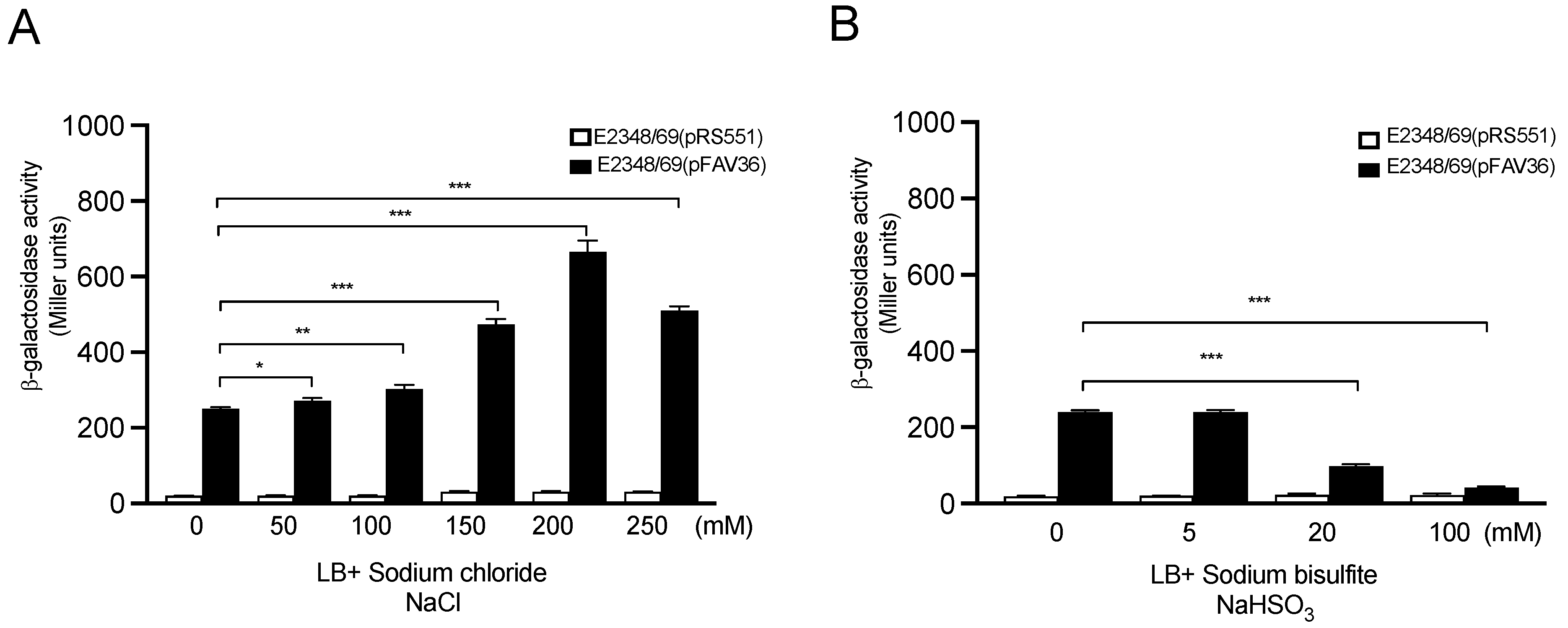
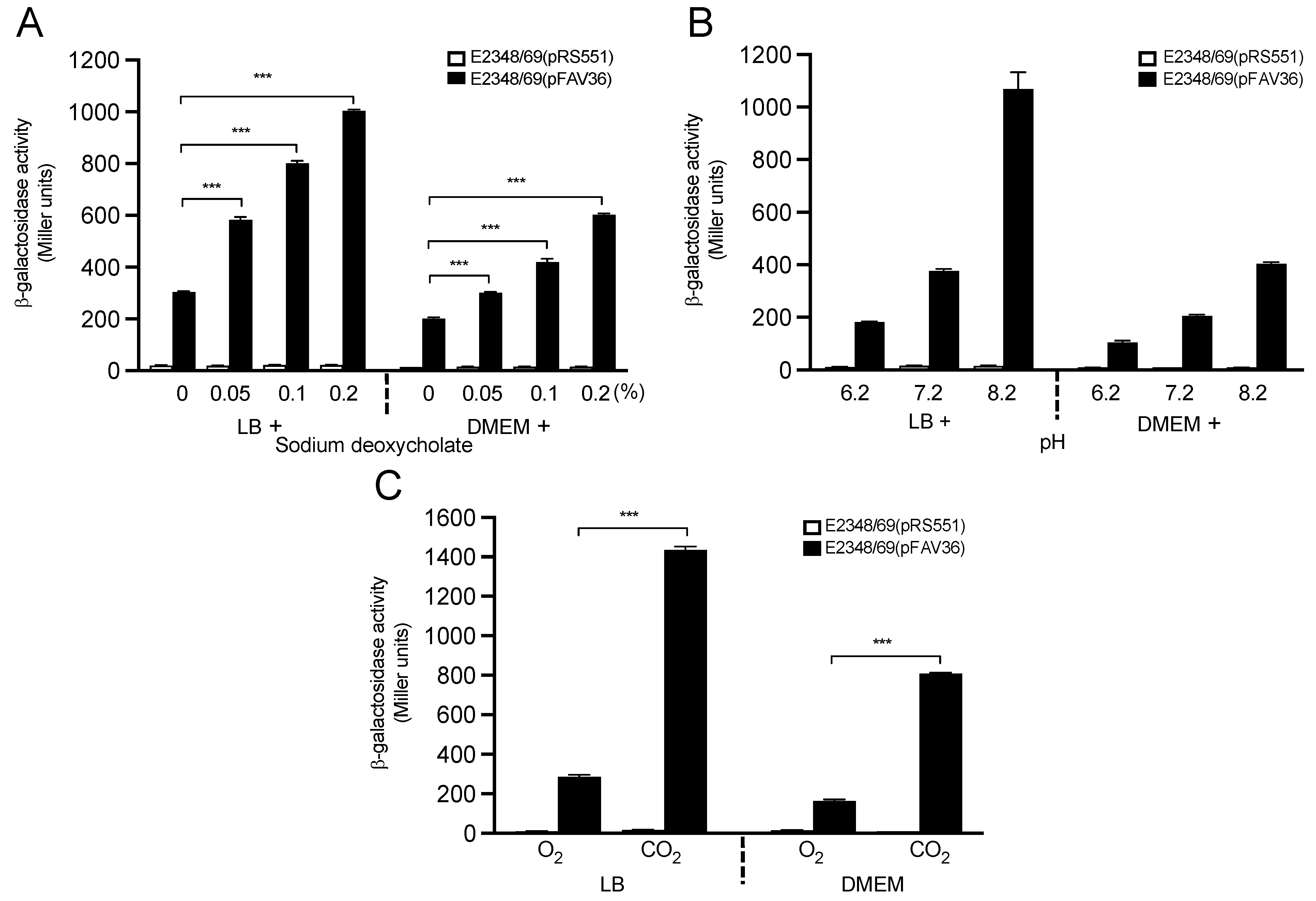
3.4. Effect of Sodium Chloride and Sodium Bisulfite on Flagella Expression
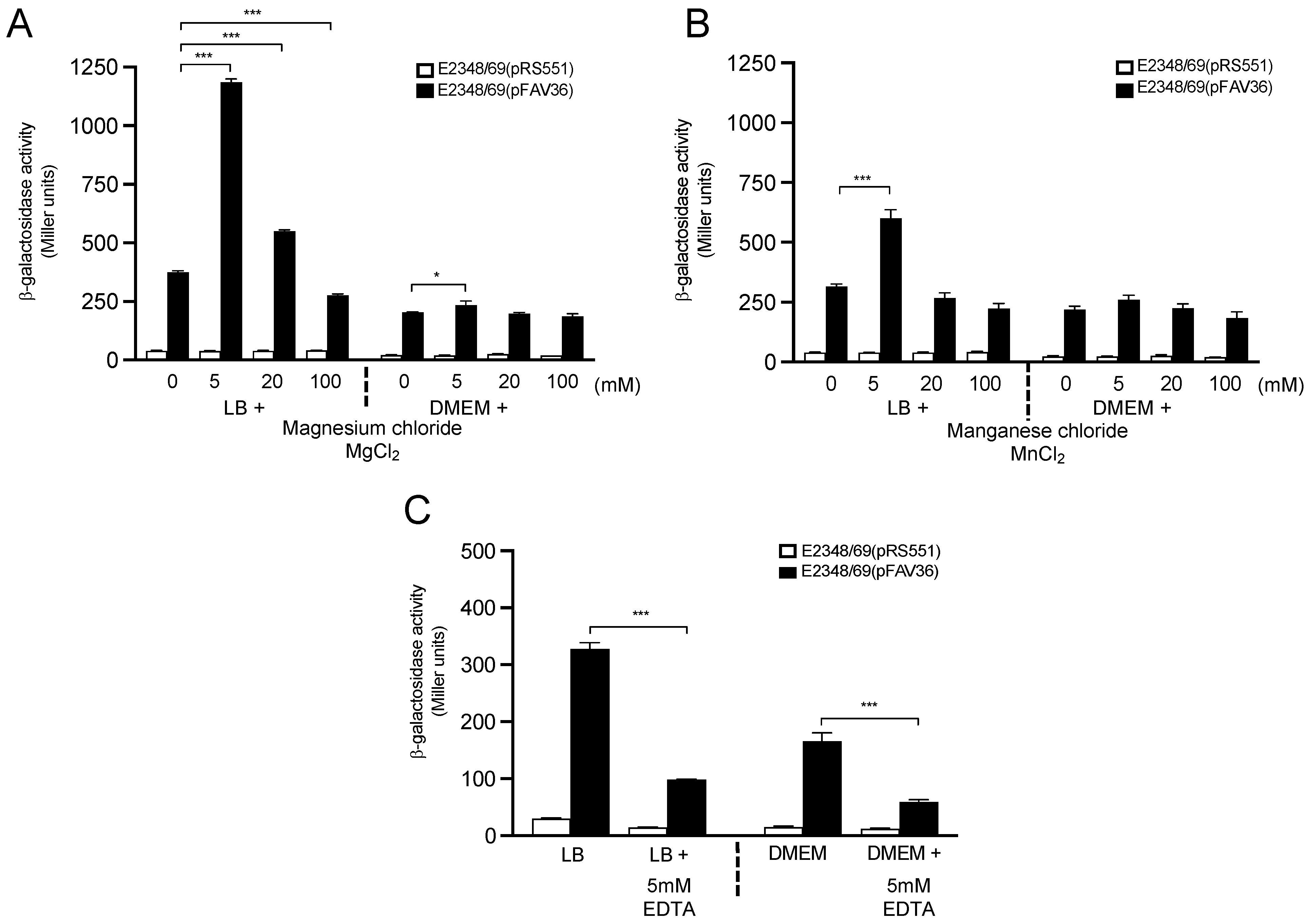
3.5. Influence of Divalent Cationic Salts on Flagella Expression
3.6. Host Gut Signals Influence Expression of EPEC fliC
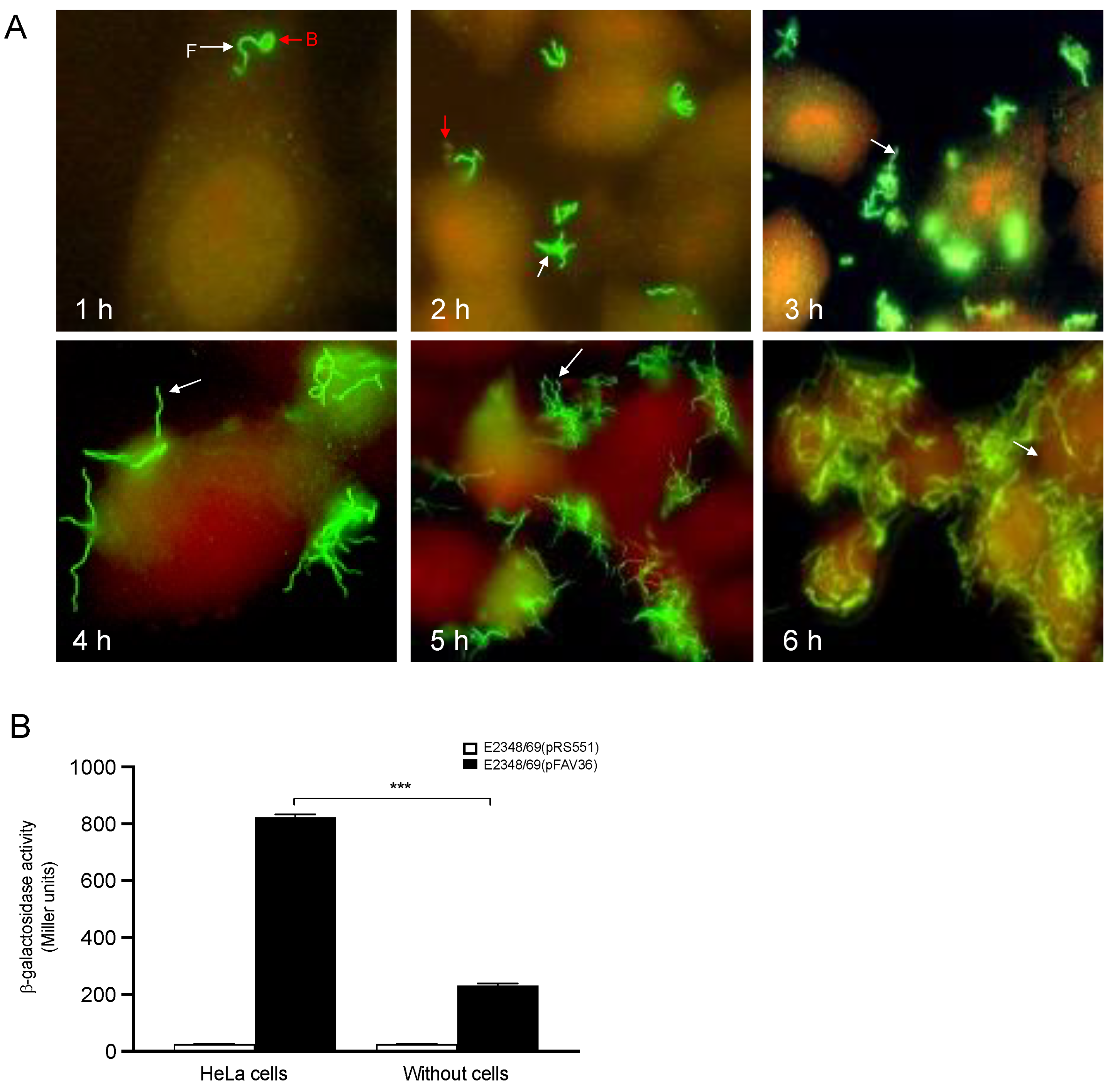
3.7. The Presence of Epithelial Cells Triggers Flagella Expression in EPEC
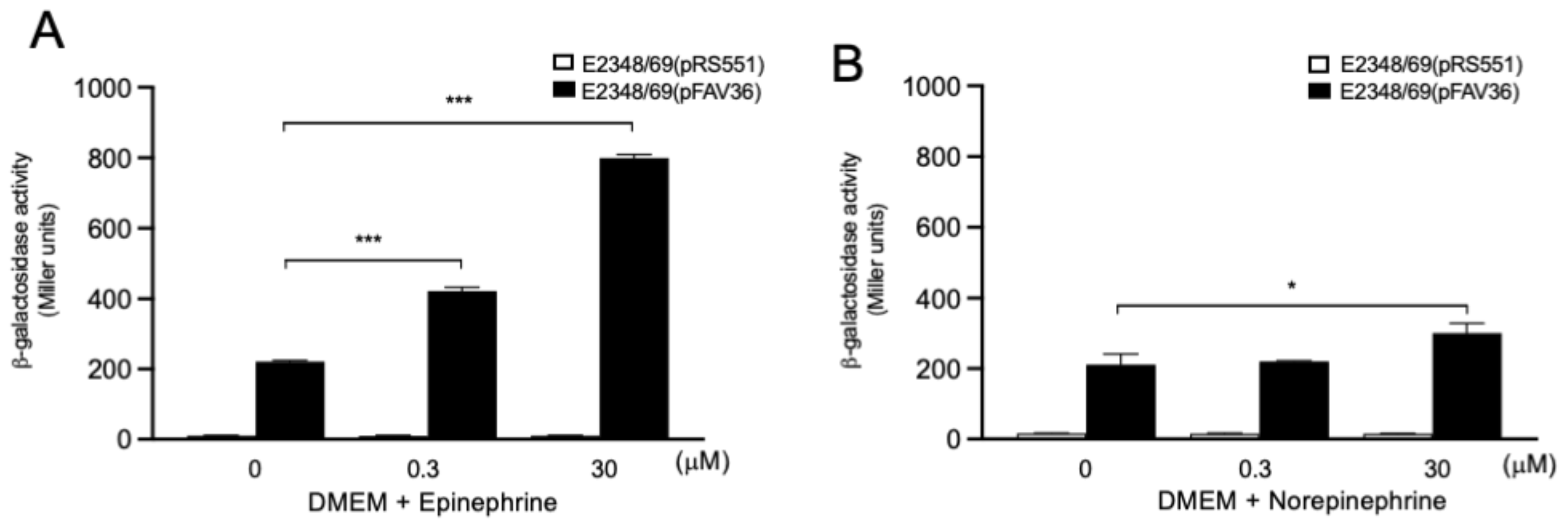
3.8. Role of Endocrine Adrenergic Molecules in EPEC fliC Expression
3.9. Host-Cell-Produced Lysophospholipids Do Not Activate fliC Transcription in EPEC
3.10. Reconstitution of a Bacteriological Medium Optimal for Flagella Expression
3.11. Motility Assay
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Nataro, J.P.; Kaper, J.B. Diarrheagenic Escherichia coli. Clin. Microbiol. Rev. 1998, 11, 142–201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Girón, J.A.; Torres, A.G.; Freer, E.; Kaper, J.B. The flagella of enteropathogenic Escherichia coli mediate adherence to epithelial cells. Mol. Microbiol. 2002, 44, 361–379. [Google Scholar] [CrossRef] [PubMed]
- Kaper, J.B.; Nataro, J.P.; Mobley, H.L. Pathogenic Escherichia coli. Nat. Rev. Microbiol. 2004, 2, 123–140. [Google Scholar] [CrossRef] [PubMed]
- Rendón, M.A.; Saldaña, Z.; Monteiro-Neto, V.; Erdem, A.L.; Vázquez, A.; Kaper, J.B.; Puente, J.L.; Girón, J.A. Commensal and pathogenic Escherichia coli use a common adherence factor for epithelial cell colonization. Proc. Natl. Acad. Sci. USA 2007, 104, 10637–10642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McDaniel, T.K.; Kaper, J.B. A cloned pathogenicity island from enteropathogenic Escherichia coli confers the attaching and effacing phenotype on E. coli K-12. Mol. Microbiol. 1997, 23, 399–407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gómez-Duarte, O.G.; Kaper, J.B. A plasmid-encoded regulatory region activates chromosomal eaeA expression in enteropathogenic Escherichia coli. Infect. Immun. 1995, 63, 1767–1776. [Google Scholar] [CrossRef] [Green Version]
- Ibarra, J.A.; Villalba, M.I.; Puente, J.L. Identification of the DNA binding sites of PerA, the transcriptional activator of the bfp and per operons in enteropathogenic Escherichia coli. J. Bacteriol. 2003, 185, 2835–2847. [Google Scholar] [CrossRef] [Green Version]
- Lara-Ochoa, C.; González-Lara, F.; Romero-González, L.E.; Jaramillo-Rodríguez, J.B.; Vázquez-Arellano, S.I.; Medrano-López, A.; Cedillo-Ramírez, L.; Martínez-Laguna, Y.; Girón, J.A.; Pérez-Rueda, E.; et al. The transcriptional activator of the bfp operon in EPEC (PerA) interacts with the RNA polymerase alpha subunit. Sci. Rep. 2021, 11, 8541. [Google Scholar] [CrossRef]
- Bustamante, V.H.; Villalba, M.I.; García-Angulo, V.A.; Vázquez, A.; Martínez, L.C.; Jiménez, R.; Puente, J.L. PerC and GrlA independently regulate Ler expression in enteropathogenic Escherichia coli. Mol. Microbiol. 2011, 82, 398–415. [Google Scholar] [CrossRef]
- Allen-Vercoe, E.; Woodward, M.J. The role of flagella, but not fimbriae, in the adherence of Salmonella enterica serotype Enteritidis to chick gut explant. J. Med. Microbiol. 1999, 48, 771–780. [Google Scholar] [CrossRef]
- Chua, K.L.; Chan, Y.Y.; Gan, Y.H. Flagella are virulence determinants of Burkholderia pseudomallei. Infect. Immun. 2003, 71, 1622–1629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gavin, R.; Merino, S.; Altarriba, M.; Canals, R.; Shaw, J.G.; Tomas, J.M. Lateral flagella are required for increased cell adherence, invasion and biofilm formation by Aeromonas spp. FEMS Microbiol. Lett. 2003, 224, 77–83. [Google Scholar] [CrossRef] [Green Version]
- Zhou, X.; Girón, J.A.; Torres, A.G.; Crawford, J.A.; Negrete, E.; Vogel, S.N.; Kaper, J.B. Flagellin of enteropathogenic Escherichia coli stimulates interleukin-8 production in T84 cells. Infect. Immun. 2003, 71, 2120–2129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xicohtencatl-Cortes, J.; Lyons, S.; Chaparro, A.P.; Hernandez, D.R.; Saldaña, Z.; Ledesma, M.A.; Rendón, M.A.; Gerwitz, A.T.; Klose, K.E.; Girón, J.A. Identification of proinflammatory flagellin proteins in supernatants of Vibrio cholerae O1 by proteomics analysis. Mol. Cell. Proteom. 2006, 5, 2374–2383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Friedlander, R.S.; Vogel, N.; Aizenberg, J. Role of flagella in adhesion of Escherichia coli to abiotic surfaces. Langmuir 2015, 31, 6137–6144. [Google Scholar] [CrossRef]
- Bente, K.; Mohammadinejad, S.; Charsooghi, M.A.; Bachmann, F.; Codutti, A.; Lefèvre, C.T.; Klumpp, S.; Faivre, D. High-speed motility originates from cooperatively pushing and pulling flagella bundles in bilophotrichous bacteria. eLife 2020, 28, e47551. [Google Scholar] [CrossRef]
- Horstmann, J.A.; Lunelli, M.; Cazzola, H.; Heidemann, J.; Kühne, C.; Steffen, P.; Szefs, S.; Rossi, C.; Lokareddy, R.K.; Wang, C.; et al. Methylation of Salmonella Typhimurium flagella promotes bacterial adhesion and host cell invasion. Nat. Commun. 2020, 11, 2013. [Google Scholar] [CrossRef] [Green Version]
- Chilcott, G.S.; Hughes, K.T. Coupling of flagellar gene expression to flagellar assembly in Salmonella enterica serovar typhimurium and Escherichia coli. Microbiol. Mol. Biol. Rev. 2000, 64, 694–708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Macnab, R.M. How bacteria assemble flagella. Annu. Rev. Microbiol. 2003, 57, 77–100. [Google Scholar] [CrossRef] [PubMed]
- Green, B.T.; Lyte, M.; Chen, C.; Xie, Y.; Casey, M.A.; Kulkarni-Narla, A.; Vulchanova, L.; Brown, D.R. Adrenergic modulation of Escherichia coli O157:H7 adherence to the colonic mucosa. Am. J. Physiol. Gastrointest. Liver. Physiol. 2004, 287, G1238–G1246. [Google Scholar] [CrossRef] [Green Version]
- Mouslim, C.; Hughes, K.T. The effect of cell growth phase on the regulatory cross-talk between flagellar and Spi1 virulence gene expression. PLoS Pathog. 2014, 10, e1003987. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Albanna, A.; Sim, M.; Hoskisson, P.A.; Gillespie, C.; Rao, C.V.; Aldridge, P.D. Driving the expression of the Salmonella enterica sv Typhimurium flagellum using flhDC from Escherichia coli results in key regulatory and cellular differences. Sci. Rep. 2018, 8, 16705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mekalanos, J.J. Environmental signals controlling expression of virulence determinants in bacteria. J. Bacteriol. 1992, 174, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soutourina, O.A.; Bertin, P.N. Regulation cascade of flagellar expression in Gram-negative bacteria. FEMS Microbiol. Rev. 2003, 27, 505–523. [Google Scholar] [CrossRef] [Green Version]
- McCarter, L.L. Regulation of flagella. Curr. Opin. Microbiol. 2006, 9, 180–186. [Google Scholar] [CrossRef]
- Lele, P.P.; Hosu, B.G.; Berg, H.C. Dynamics of mechanosensing in the bacterial flagellar motor. Proc. Natl. Acad. Sci. USA 2013, 110, 11839–11844. [Google Scholar] [CrossRef] [Green Version]
- Spöring, I.; Felgner, S.; Preuße, M.; Eckweiler, D.; Rohde, M.; Häussler, S.; Weiss, S.; Erhardt, M. Regulation of flagellum biosynthesis in response to cell envelope stress in Salmonella enterica serovar Typhimurium. mBio 2018, 9, e00736-17. [Google Scholar] [CrossRef] [Green Version]
- Adler, J.; Templeton, B. The effect of environmental conditions on the motility of Escherichia coli. J. Gen. Microbiol. 1967, 46, 175–184. [Google Scholar] [CrossRef] [Green Version]
- Alm, R.A.; Guerry, P.; Trust, T.J. The Campylobacter sigma 54 flaB flagellin promoter is subject to environmental regulation. J. Bacteriol. 1993, 175, 4448–4455. [Google Scholar] [CrossRef] [Green Version]
- Harshey, R.M. Bacterial motility on a surface: Many ways to a common goal. Annu. Rev. Microbiol. 2003, 57, 249–273. [Google Scholar] [CrossRef]
- Maurer, L.M.; Yohannes, E.; Bondurant, S.S.; Radmacher, M.; Slonczewski, J.L. pH regulates genes for flagellar motility, catabolism, and oxidative stress in Escherichia coli K-12. J. Bacteriol. 2005, 187, 304–319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hockett, K.L.; Burch, A.Y.; Lindow, S.E. Thermo-regulation of genes mediating motility and plant interactions in Pseudomonas syringae. PLoS ONE 2013, 8, e59850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rudenko, I.; Ni, B.; Glatter, T.; Sourjik, V. Inefficient secretion of anti-sigma factor FlgM inhibits bacterial motility at high temperature. iScience 2019, 16, 145–154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erdem, A.L.; Avelino, F.; Xicohtencatl-Cortes, J.; Girón, J.A. Host protein binding and adhesive properties of H6 and H7 flagella of attaching and effacing Escherichia coli. J. Bacteriol. 2007, 189, 7426–7435. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.; Si, M.; Song, Y.; Zhu, W.; Gao, F.; Wang, Y.; Zhang, L.; Zhang, W.; Wei, G.; Luo, Z.Q.; et al. Type VI secretion system transports Zn2+ to combat multiple stresses and host immunity. PLoS Pathog. 2015, 11, e1005020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Ren, F.; Cai, G.; Huang, P.; Chai, Q.; Gundogdu, O.; Jiao, X.; Huang, J. Investigating the role of FlhF identifies novel interactions with genes involved in flagellar synthesis in Campylobacter jejuni. Front. Microbiol. 2020, 11, 460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Speare, L.; Woo, M.; Bultman, K.M.; Mandel, M.J.; Wollenberg, M.S.; Septer, A.N. Host-Like conditions are required for T6SS-mediated competition among Vibrio fischeri light organ symbionts. mSphere 2021, 6, e0128820. [Google Scholar] [CrossRef] [PubMed]
- Allsopp, L.P.; Collins, A.C.Z.; Hawkins, E.; Wood, T.E.; Filloux, A. RpoN/Sfa2-dependent activation of the Pseudomonas aeruginosa H2-T6SS and its cognate arsenal of antibacterial toxins. Nucleic Acids Res. 2022, 50, 227–243. [Google Scholar] [CrossRef]
- Alvarez-Fraga, L.; Phan, M.D.; Goh, K.G.K.; Nhu, N.T.K.; Hancock, S.J.; Allsopp, L.P.; Peters, K.M.; Forde, B.M.; Roberts, L.W.; Sullivan, M.J.; et al. Differential Afa/Dr fimbriae expression in the multidrug-resistant Escherichia coli ST131 clone. mBio 2022, 13, e0351921. [Google Scholar] [CrossRef]
- Miller, J.H. Experiments in Molecular Genetics; Cold Spring Harbor: New York, NY, USA, 1972. [Google Scholar]
- Sperandio, V.; Torres, A.G.; Girón, J.A.; Kaper, J.B. Quorum sensing is a global regulatory mechanism in enterohemorrhagic Escherichia coli O157:H7. J. Bacteriol. 2001, 183, 5187–5197. [Google Scholar] [CrossRef] [Green Version]
- Schwan, W.R.; Lee, J.L.; Lenard, F.A.; Matthews, B.T.; Beck, M.T. Osmolarity and pH growth conditions regulate fim gene transcription and type 1 pilus expression in uropathogenic Escherichia coli. Infect. Immun. 2002, 70, 1391–1402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Subramanian, N.; Qadri, A. Lysophospholipid sensing triggers secretion of flagellin from pathogenic salmonella. Nat. Immunol. 2006, 7, 583–589. [Google Scholar] [CrossRef]
- Castell, D.O.; Moore, E.W. Ammonia absorption from the human colon. The role of nonionic diffusion. Gastroenterology 1971, 60, 33–42. [Google Scholar] [CrossRef]
- Ferraris, R.P.; Yasharpour, S.; Lloyd, K.C.; Mirzayan, R.; Diamond, J.M. Luminal glucose concentrations in the gut under normal conditions. Am. J. Physiol. Liver Physiol. 1990, 259, G822–G837. [Google Scholar] [CrossRef] [PubMed]
- Mossberg, S.M.; Ross, G. Ammonia movement in the small intestine: Preferential transport by the ileum. J. Clin. Investig. 1967, 46, 490–498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abe, H.; Tatsuno, I.; Tobe, T.; Okutani, A.; Sasakawa, C. Bicarbonate ion stimulates the expression of locus of enterocyte effacement-encoded genes in enterohemorrhagic Escherichia coli O157:H7. Infect. Immun. 2002, 70, 3500–3509. [Google Scholar] [CrossRef] [Green Version]
- Elliott, S.J.; Sperandio, V.; Girón, J.A.; Shin, S.; Mellies, J.L.; Wainwright, L.; Hutcheson, S.W.; McDaniel, T.K.; Kaper, J.B. The locus of enterocyte effacement (LEE)-encoded regulator controls expression of both LEE- and non-LEE-encoded virulence factors in enteropathogenic and enterohemorrhagic Escherichia coli. Infect. Immun. 2000, 68, 6115–6126. [Google Scholar] [CrossRef]
- Puente, J.L.; Bieber, D.; Ramer, S.W.; Murray, W.; Schoolnik, G.K. The bundle-forming pili of enteropathogenic Escherichia coli: Transcriptional regulation by environmental signals. Mol. Microbiol. 1996, 20, 87–100. [Google Scholar] [CrossRef]
- Gismero-Ordoñez, J.; Dall’Agnol, M.; Trabulsi, L.R.; Girón, J.A. Expression of the bundle-forming pilus by enteropathogenic Escherichia coli strains of heterologous serotypes. J. Clin. Microbiol. 2002, 40, 2291–2296. [Google Scholar] [CrossRef] [Green Version]
- Kenny, B.; Abe, A.; Stein, M.; Finlay, B.B. Enteropathogenic Escherichia coli protein secretion is induced in response to conditions similar to those in the gastrointestinal tract. Infect. Immun. 1997, 65, 2606–2612. [Google Scholar] [CrossRef] [Green Version]
- Girón, J.A.; Ho, A.S.; Schoolnik, G.K. An inducible bundle-forming pilus of enteropathogenic Escherichia coli. Science 1991, 254, 710–713. [Google Scholar] [CrossRef] [PubMed]
- Akerley, B.J.; Cotter, P.A.; Miller, J.F. Ectopic expression of the flagellar regulon alters development of the Bordetella-host interaction. Cell 1995, 80, 611–620. [Google Scholar] [CrossRef] [Green Version]
- Schmitt, C.K.; Ikeda, J.S.; Darnell, S.C.; Watson, P.R.; Bispham, J.; Wallis, T.S.; Weinstein, D.L.; Metcalf, E.S.; O’Brien, A.D. Absence of all components of the flagellar export and synthesis machinery differentially alters virulence of Salmonella enterica serovar Typhimurium in models of typhoid fever, survival in macrophages, tissue culture invasiveness, and calf enterocolitis. Infect. Immun. 2001, 69, 5619–5625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lyte, M.; Arulanandam, B.; Nguyen, K.; Frank, C.; Erickson, A.; Francis, D. Norepinephrine induced growth and expression of virulence associated factors in enterotoxigenic and enterohemorrhagic strains of Escherichia coli. Adv. Exp. Med. Biol. 1997, 412, 331–339. [Google Scholar]
- Sperandio, V.; Torres, A.G.; Jarvis, B.; Nataro, J.P.; Kaper, J.B. Bacteria-host communication: The language of hormones. Proc. Natl. Acad. Sci. USA 2003, 100, 8951–8956. [Google Scholar] [CrossRef] [Green Version]
- Bansal, T.; Englert, D.; Lee, J.; Hegde, M.; Wood, T.K.; Jayaraman, A. Differential effects of epinephrine, norepinephrine, and indole on Escherichia coli O157:H7 chemotaxis, colonization, and gene expression. Infect. Immun. 2007, 75, 4597–4607. [Google Scholar] [CrossRef] [Green Version]
- Cotter, P.A.; Miller, J.F. Triggering bacterial virulence. Science 1996, 273, 1183–1184. [Google Scholar] [CrossRef]
- De la Cruz, M.A.; Ares, M.A.; von Bargen, K.; Panunzi, L.G.; Martínez-Cruz, J.; Valdez-Salazar, H.A.; Jiménez-Galicia, C.; Torres, J. Gene expression profiling of transcription factors of Helicobacter pylori under different environmental conditions. Front. Microbiol. 2017, 8, 615. [Google Scholar] [CrossRef] [Green Version]
- Ares, M.A.; Abundes-Gallegos, J.; Rodríguez-Valverde, D.; Panunzi, L.G.; Jiménez-Galicia, C.; Jarillo-Quijada, M.D.; Cedillo, M.L.; Alcántar-Curiel, M.D.; Torres, J.; Girón, J.A.; et al. The coli surface antigen CS3 of Enterotoxigenic Escherichia coli is differentially regulated by H-NS, CRP, and CpxRA global regulators. Front. Microbiol. 2019, 10, 1685. [Google Scholar] [CrossRef]
- Do, H.; Makthal, N.; VanderWal, A.R.; Saavedra, M.O.; Olsen, R.J.; Musser, J.M.; Kumaraswami, M. Environmental pH and peptide signaling control virulence of Streptococcus pyogenes via a quorum-sensing pathway. Nat. Commun. 2019, 10, 2586. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Valverde, D.; León-Montes, N.; Soria-Bustos, J.; Martínez-Cruz, J.; González-Ugalde, R.; Rivera-Gutiérrez, S.; González-Y.-Merchand, J.A.; Rosales-Reyes, R.; García-Morales, L.; Hirakawa, H.; et al. cAMP receptor protein positively regulates the expression of genes involved in the biosynthesis of Klebsiella oxytoca tilivalline cytotoxin. Front. Microbiol. 2021, 12, 743594. [Google Scholar] [CrossRef] [PubMed]
- Valdez-Salazar, H.A.; Ares, M.A.; Fernández, F.J.; Ibarra, J.A.; Torres, J.; Bustamante, V.H.; De la Cruz, M.A. Long-chain fatty acids alter transcription of Helicobacter pylori virulence and regulatory genes. PeerJ 2021, 9, e12270. [Google Scholar] [CrossRef] [PubMed]
- Leverton, L.Q.; Kaper, J.B. Temporal expression of enteropathogenic Escherichia coli virulence genes in an in vitro model of infection. Infect. Immun. 2005, 73, 1034–1043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ronin, I.; Katsowich, N.; Rosenshine, I.; Balaban, N.Q. A long-term epigenetic memory switch controls bacterial virulence bimodality. eLife 2017, 6, e19599. [Google Scholar] [CrossRef] [PubMed]
- Platenkamp, A.; Mellies, J.L. Environment controls LEE regulation in enteropathogenic Escherichia coli. Front. Microbiol. 2018, 9, 1694. [Google Scholar] [CrossRef] [Green Version]
- Walker, S.L.; Sojka, M.; Dibb-Fuller, M.; Woodward, M.J. Effect of pH, temperature and surface contact on the elaboration of fimbriae and flagella by Salmonella serotype Enteritidis. J. Med. Microbiol. 1999, 48, 253–261. [Google Scholar] [CrossRef]
- Zhao, K.; Liu, M.; Burgess, R.R. Adaptation in bacterial flagellar and motility systems: From regulon members to ‘foraging’-like behavior in E. coli. Nucleic Acids Res. 2007, 35, 4441–4452. [Google Scholar] [CrossRef] [Green Version]
- Wang, F.; Deng, L.; Huang, F.; Wang, Z.; Lu, Q.; Xu, C. Flagellar motility is critical for Salmonella enterica serovar Typhimurium biofilm development. Front. Microbiol. 2020, 11, 1695. [Google Scholar] [CrossRef]
- Yang, J.; Hart, E.; Tauschek, M.; Price, G.D.; Hartland, E.L.; Strugnell, R.A.; Robins-Browne, R.M. Bicarbonate-mediated transcriptional activation of divergent operons by the virulence regulatory protein, RegA, from Citrobacter rodentium. Mol. Microbiol. 2008, 68, 314–327. [Google Scholar] [CrossRef]
- Abuaita, B.H.; Withey, J.H. Bicarbonate Induces Vibrio cholerae virulence gene expression by enhancing ToxT activity. Infect. Immun. 2009, 77, 4111–4120. [Google Scholar] [CrossRef] [Green Version]
- Migliore, F.; Macchi, R.; Landini, P.; Paroni, M. Phagocytosis and epithelial cell invasion by Crohn’s disease-associated adherent-invasive Escherichia coli are inhibited by the anti-inflammatory drug 6-Mercaptopurine. Front. Microbiol. 2018, 9, 964. [Google Scholar] [CrossRef] [PubMed]
- Bijlsma, J.J.; Groisman, E.A. The PhoP/PhoQ system controls the intramacrophage type three secretion system of Salmonella enterica. Mol. Microbiol. 2005, 57, 85–96. [Google Scholar] [CrossRef] [PubMed]
- De la Cruz, M.A.; Ruiz-Tagle, A.; Ares, M.A.; Pacheco, S.; Yáñez, J.A.; Cedillo, L.; Torres, J.; Girón, J.A. The expression of Longus type 4 pilus of enterotoxigenic Escherichia coli is regulated by LngR and LngS and by H-NS, CpxR and CRP global regulators. Environ. Microbiol. 2017, 19, 1761–1775. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Han, R.; Wang, J.; Yang, P.; Wang, F.; Yang, B. Magnesium sensing regulates intestinal colonization of Enterohemorrhagic Escherichia coli O157:H7. mBio 2020, 11, e02470-20. [Google Scholar] [CrossRef]
- Deng, W.; Puente, J.L.; Gruenheid, S.; Li, Y.; Vallance, B.A.; Vázquez, A.; Barba, J.; Ibarra, J.A.; O’Donnell, P.; Metalnikov, P.; et al. Dissecting virulence: Systematic and functional analyses of a pathogenicity island. Proc. Natl. Acad. Sci. USA 2004, 101, 3597–3602. [Google Scholar] [CrossRef] [Green Version]
- Gaytán, M.O.; Monjarás Feria, J.; Soto, E.; Espinosa, N.; Benítez, J.M.; Georgellis, D.; González-Pedrajo, B. Novel insights into the mechanism of SepL-mediated control of effector secretion in enteropathogenic Escherichia coli. Microbiologyopen 2017, 7, e00571. [Google Scholar] [CrossRef] [Green Version]
- Shaulov, L.; Gershberg, J.; Deng, W.; Finlay, B.B.; Sal-Man, N. The ruler protein EscP of the enteropathogenic Escherichia coli type III secretion system is involved in calcium sensing and secretion hierarchy regulation by interacting with the gatekeeper protein SepL. mBio 2017, 8, e01733-16. [Google Scholar] [CrossRef] [Green Version]
- Yarwood, J.M.; Schlievert, P.M. Oxygen and carbon dioxide regulation of toxic shock syndrome toxin 1 production by Staphylococcus aureus MN8. J. Clin. Microbiol. 2000, 38, 1797–1803. [Google Scholar] [CrossRef] [Green Version]
- Ramos-Morales, F. Acidic pH: Enemy or ally for enteric bacteria? Virulence 2012, 3, 103–106. [Google Scholar] [CrossRef] [Green Version]
- Danielsson, H. Influence of bile acids on digestion and absorption of lipids. Am. J. Clin. Nutr. 1963, 12, 214–219. [Google Scholar] [CrossRef]
- Chiang, J.Y.L.; Ferrell, J.M. Bile acids as metabolic regulators and nutrient sensors. Annu. Rev. Nutr. 2019, 39, 175–200. [Google Scholar] [CrossRef] [PubMed]
- Hung, D.T.; Mekalanos, J.J. Bile acids induce cholera toxin expression in Vibrio cholerae in a ToxT-independent manner. Proc. Natl. Acad. Sci. USA 2005, 102, 3028–3033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hung, D.T.; Zhu, J.; Sturtevant, D.; Mekalanos, J.J. Bile acids stimulate biofilm formation in Vibrio cholerae. Mol. Microbiol. 2006, 59, 193–201. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Liu, Z.; Hughes, C.; Stern, A.M.; Wang, H.; Zhong, Z.; Kan, B.; Fenical, W.; Zhu, J. Bile salt-induced intermolecular disulfide bond formation activates Vibrio cholerae virulence. Proc. Natl. Acad. Sci. USA 2013, 110, 2348–2353. [Google Scholar] [CrossRef] [Green Version]
- D’Mello, A.; Yotis, W.W. The action of sodium deoxycholate on Escherichia coli. Appl. Environ. Microbiol. 1987, 53, 1944–1946. [Google Scholar] [CrossRef] [Green Version]
- Lyte, M. Microbial endocrinology and infectious disease in the 21st century. Trends Microbiol. 2004, 12, 14–20. [Google Scholar] [CrossRef]
- Cambronel, M.; Nilly, F.; Mesguida, O.; Boukerb, A.M.; Racine, P.J.; Baccouri, O.; Borrel, V.; Martel, J.; Fécamp, F.; Knowlton, R.; et al. Influence of catecholamines (Epinephrine/Norepinephrine) on biofilm formation and adhesion in pathogenic and probiotic strains of Enterococcus faecalis. Front. Microbiol. 2020, 11, 1501. [Google Scholar] [CrossRef]
- Alverdy, J.; Holbrook, C.; Rocha, F.; Seiden, L.; Wu, R.L.; Musch, M.; Chang, E.; Ohman, D.; Suh, S. Gut-derived sepsis occurs when the right pathogen with the right virulence genes meets the right host: Evidence for in vivo virulence expression in Pseudomonas aeruginosa. Ann. Surg. 2000, 232, 480–489. [Google Scholar] [CrossRef]
- Cogan, T.A.; Thomas, A.O.; Rees, L.E.; Taylor, A.H.; Jepson, M.A.; Williams, P.H.; Ketley, J.; Humphrey, T.J. Norepinephrine increases the pathogenic potential of Campylobacter jejuni. Gut 2007, 56, 1060–1065. [Google Scholar] [CrossRef] [Green Version]
- Walters, M.; Sperandio, V. Autoinducer 3 and epinephrine signaling in the kinetics of locus of enterocyte effacement gene expression in enterohemorrhagic Escherichia coli. Infect. Immun. 2006, 74, 5445–5455. [Google Scholar] [CrossRef] [Green Version]
- Beata, S.; Michał, T.; Mateusz, O.; Urszula, W.; Choroszy, M.; Andrzej, T.; Piotr, D. Norepinephrine affects the interaction of adherent-invasive Escherichia coli with intestinal epithelial cells. Virulence 2021, 12, 630–637. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Condition or Substance Added to DMEM | fliC Transcription | Motility |
|---|---|---|
| Unmodified DMEM | - | - |
| Ammonium sulfate 5 mM | - | - |
| Sodium chloride 200 mM | ++ | + |
| Sodium bisulfite 5 mM | - | - |
| Sodium bicarbonate 44 mM * | - | - |
| Magnesium chloride 5 mM | + | + |
| Magnesium sulfate 5 mM | + | + |
| Manganese chloride 5 mM | + | + |
| Calcium chloride 5 mM | + | +++ |
| EDTA 5 mM | - | - |
| Sodium deoxycholate 0.2% | ++ | ++ |
| pH 6.2 | - | - |
| pH 8.2 | + | + |
| 5% CO2 | +++ | +++ |
| HeLa cells | +++ | +++ |
| Epinephrine 30 µM | ++ | +++ |
| Norepinephrine 30 µM | + | +++ |
| Lysophosphatidic acid 200 mM | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Avelino-Flores, F.; Soria-Bustos, J.; Saldaña-Ahuactzi, Z.; Martínez-Laguna, Y.; Yañez-Santos, J.A.; Cedillo-Ramírez, M.L.; Girón, J.A. The Transcription of Flagella of Enteropathogenic Escherichia coli O127:H6 Is Activated in Response to Environmental and Nutritional Signals. Microorganisms 2022, 10, 792. https://doi.org/10.3390/microorganisms10040792
Avelino-Flores F, Soria-Bustos J, Saldaña-Ahuactzi Z, Martínez-Laguna Y, Yañez-Santos JA, Cedillo-Ramírez ML, Girón JA. The Transcription of Flagella of Enteropathogenic Escherichia coli O127:H6 Is Activated in Response to Environmental and Nutritional Signals. Microorganisms. 2022; 10(4):792. https://doi.org/10.3390/microorganisms10040792
Chicago/Turabian StyleAvelino-Flores, Fabiola, Jorge Soria-Bustos, Zeus Saldaña-Ahuactzi, Ygnacio Martínez-Laguna, Jorge A. Yañez-Santos, María L. Cedillo-Ramírez, and Jorge A. Girón. 2022. "The Transcription of Flagella of Enteropathogenic Escherichia coli O127:H6 Is Activated in Response to Environmental and Nutritional Signals" Microorganisms 10, no. 4: 792. https://doi.org/10.3390/microorganisms10040792