
Characterization of Some Salt-Tolerant Bacterial Hydrolases with Potential Utility in Cultural Heritage Bio-Cleaning

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial and Fungal Sources of Hydrolytic Enzymes
2.2. Taxonomic Identification of the Fungal Strains
2.3. Culture Conditions for Enzyme Production
2.4. Quantitative Evaluation of Hydrolytic Activities
2.4.1. Protease Activity Assay
2.4.2. Esterase Activity Assay
2.4.3. Cellulase Activity Assay
2.4.4. Xylanase Activity Assay
2.5. Precipitation of Extracellular Hydrolases
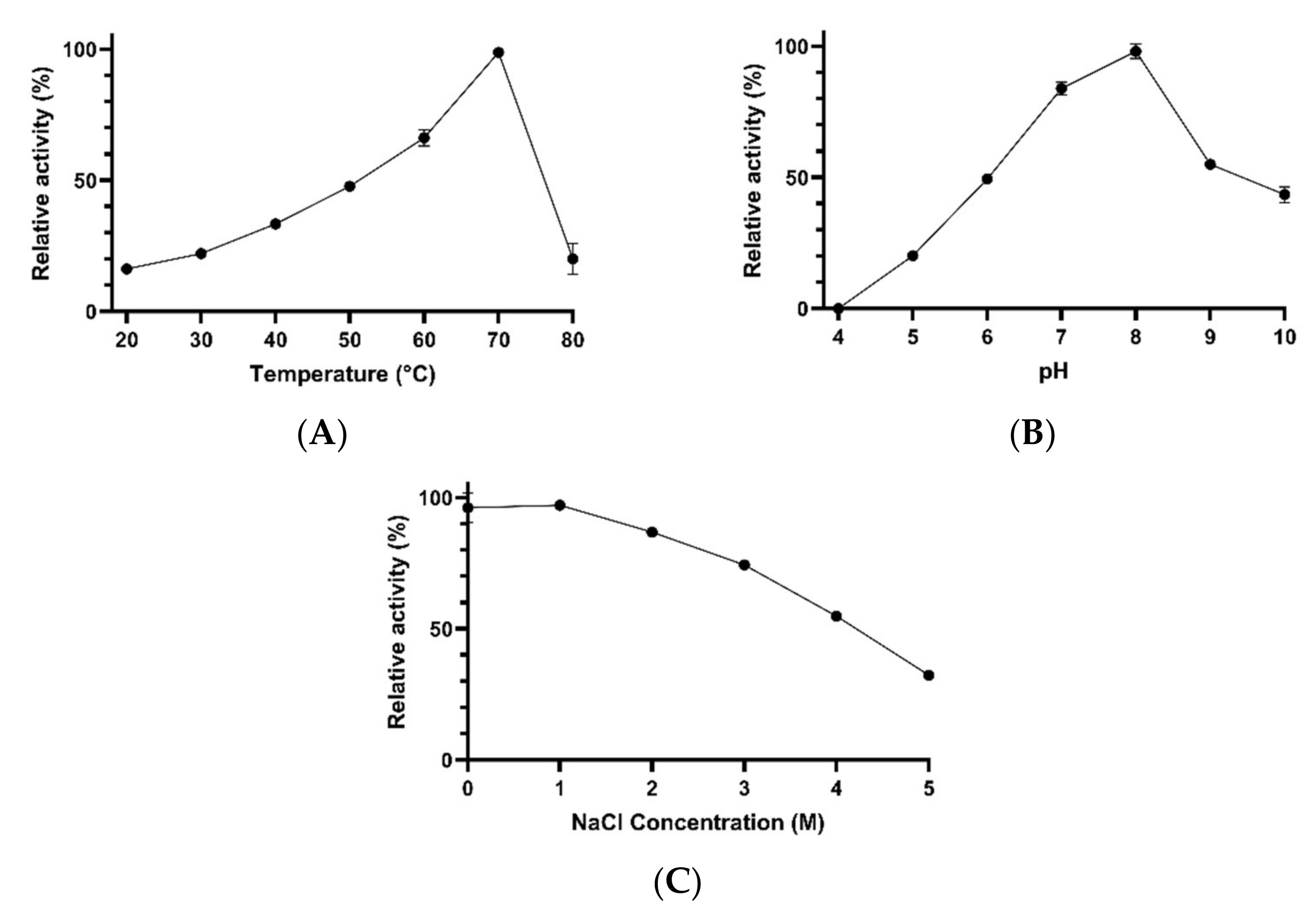
2.6. Evaluation of the Effects of Physicochemical Factors on the Activity and Stability of Enzymes
2.7. Preliminary Evaluation of the Efficacy of Halotolerant Esterases in Bio-Cleaning Applications
3. Results
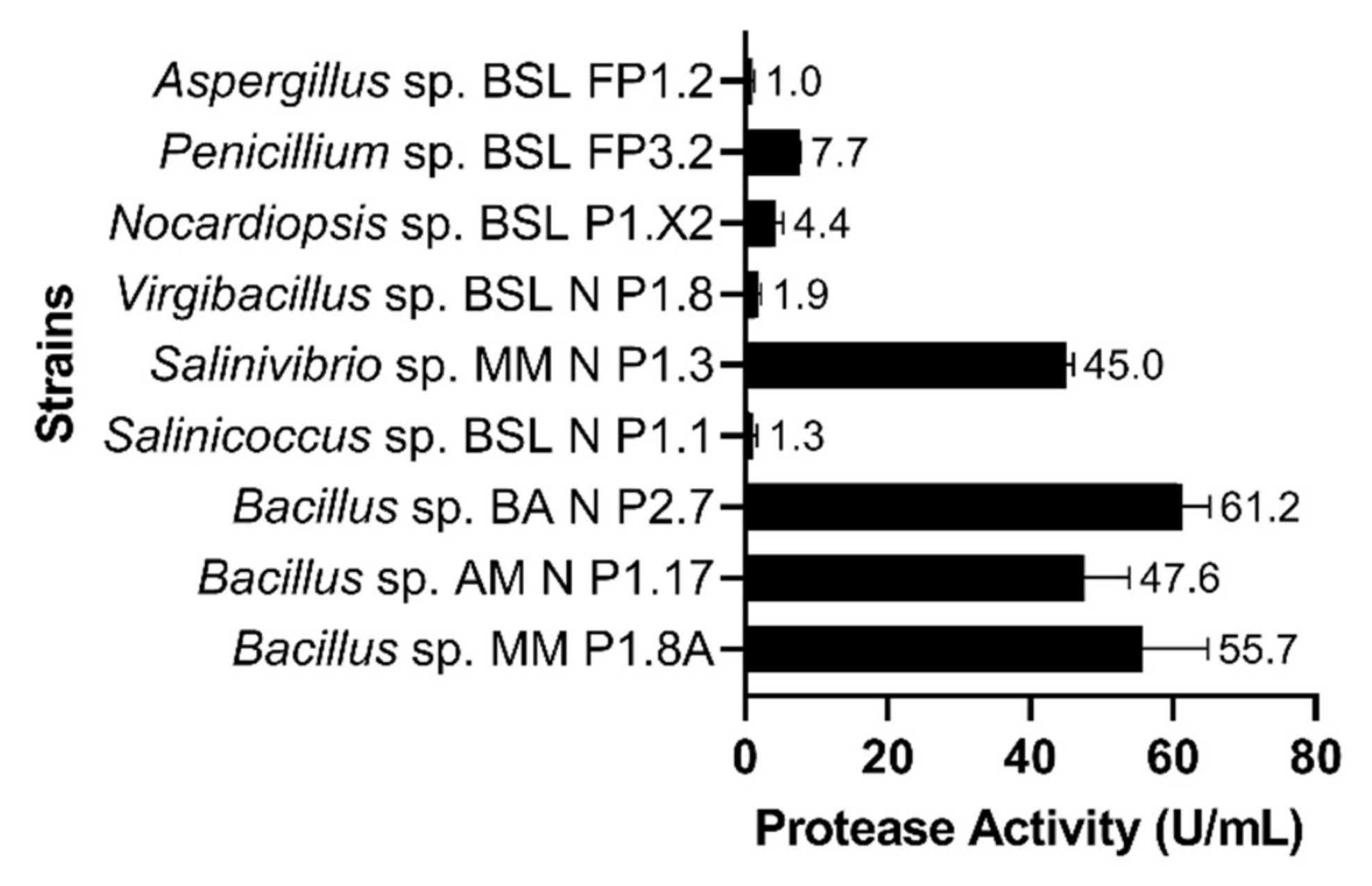
3.1. Characterization of Protease Activities
3.2. Characterization of Esterase Activities
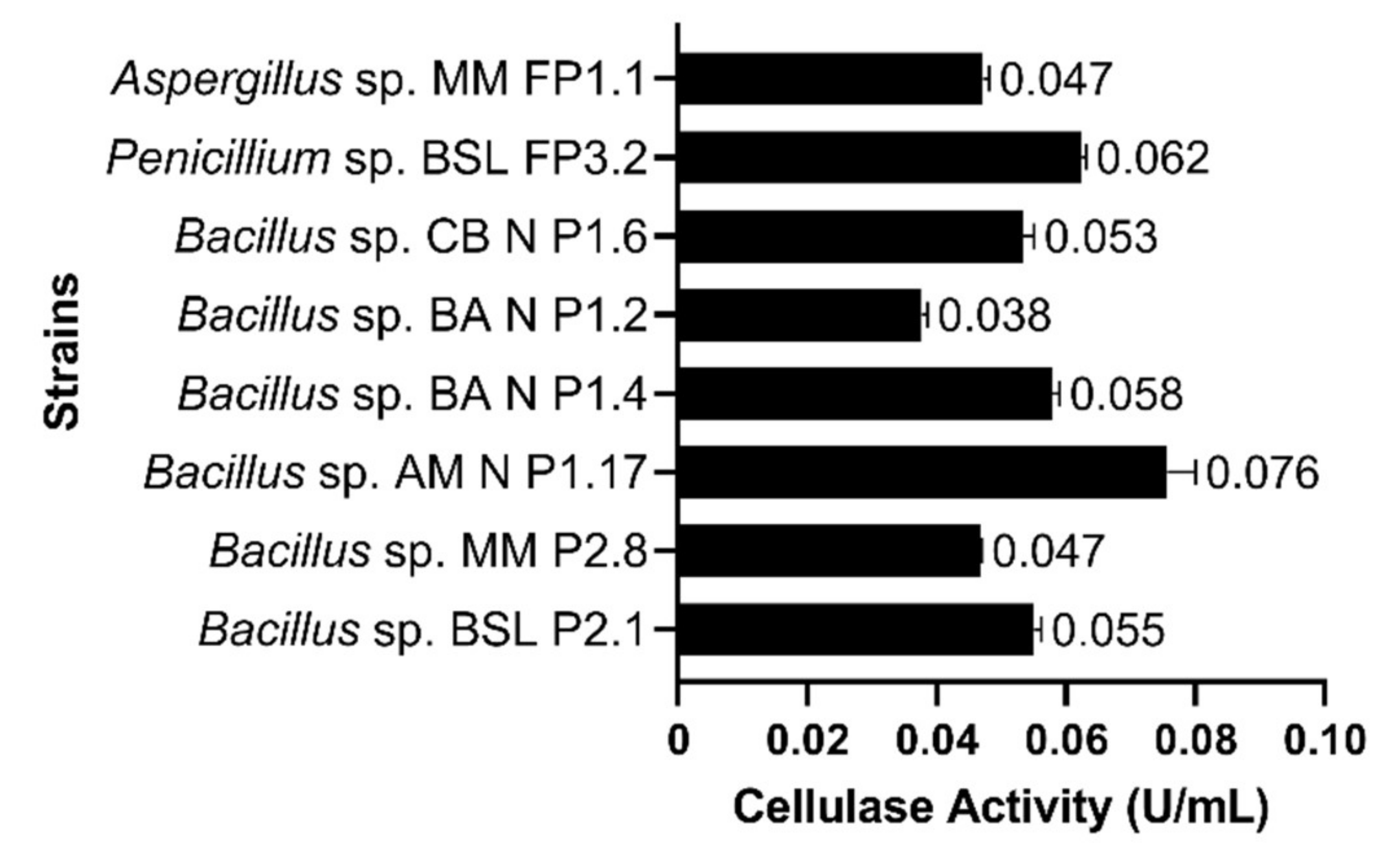
3.3. Characterization of Cellulase Activities
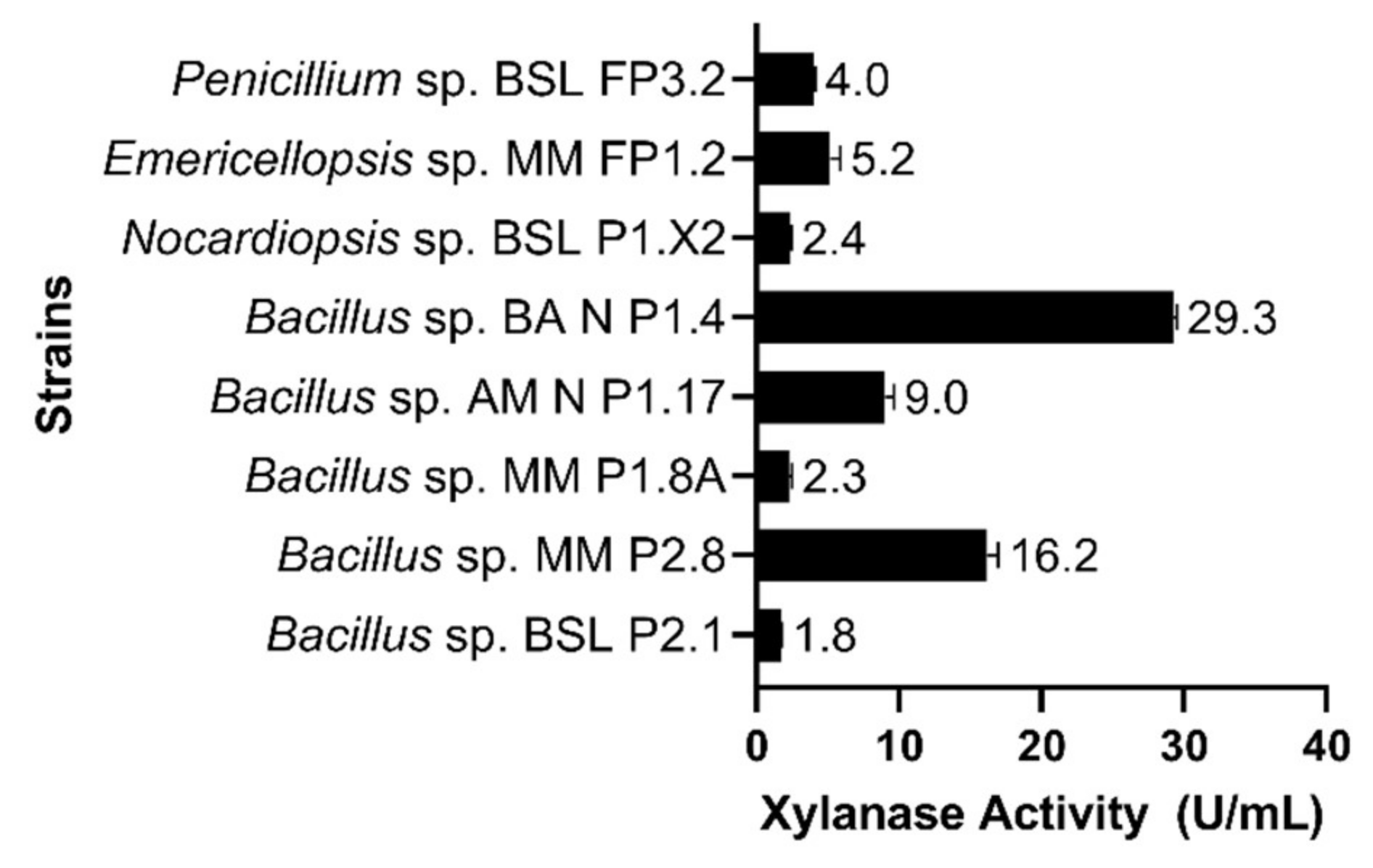
3.4. Characterization of Xylanase Activities
3.5. The Efficacy of Halotolerant Esterases in the Removal of Paraloid B-72 and Oil
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Robinson, P.K. Enzymes: Principles and Biotechnological Applications. Essays Biochem. 2015, 59, 1–41. [Google Scholar] [CrossRef] [PubMed]
- Chapman, J.; Ismail, A.; Dinu, C. Industrial Applications of Enzymes: Recent Advances, Techniques, and Outlooks. Catalysts 2018, 8, 238. [Google Scholar] [CrossRef] [Green Version]
- BBC Research. Available online: https://www.bccresearch.com/market-research/biotechnology/global-markets-for-enzymes-in-industrial-applications.html (accessed on 9 November 2021).
- Nigam, P. Microbial Enzymes with Special Characteristics for Biotechnological Applications. Biomolecules 2013, 3, 597. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.; Qin, H.; Chen, X.; Zhang, Y.; Xue, Y.; Ma, Y. A Novel Alkaline Protease from Alkaliphilic Idiomarina Sp. C9-1 with Potential Application for Eco-Friendly Enzymatic Dehairing in the Leather Industry. Sci. Rep. 2018, 8, 16467. [Google Scholar] [CrossRef] [PubMed]
- Karan, R.; Mathew, S.; Muhammad, R.; Bautista, D.B.; Vogler, M.; Eppinger, J.; Oliva, R.; Cavallo, L.; Arold, S.T.; Rueping, M. Understanding High-Salt and Cold Adaptation of a Polyextremophilic Enzyme. Microorganisms 2020, 8, 1594. [Google Scholar] [CrossRef] [PubMed]
- McGenity, T.J.; Oren, A. Hypersaline Environments. In Life at Extremes: Environments, Organisms and Strategies for Survival; Bell, E.M., Ed.; CAB International: Wallingford, UK, 2012; pp. 402–437. [Google Scholar] [CrossRef]
- Tadeo, X.; López-Méndez, B.; Trigueros, T.; Laín, A.; Castaño, D.; Millet, O. Structural Basis for the Aminoacid Composition of Proteins from Halophilic Archea. PLoS Biol. 2009, 7, e1000257. [Google Scholar] [CrossRef] [Green Version]
- Siglioccolo, A.; Paiardini, A.; Piscitelli, M.; Pascarella, S. Structural Adaptation of Extreme Halophilic Proteins through Decrease of Conserved Hydrophobic Contact Surface. BMC Struct. Biol. 2011, 11, 50. [Google Scholar] [CrossRef] [Green Version]
- Amoozegar, M.A.; Safarpour, A.; Noghabi, K.A.; Bakhtiary, T.; Ventosa, A. Halophiles and Their Vast Potential in Biofuel Production. Front. Microbiol. 2019, 10, 1895. [Google Scholar] [CrossRef]
- Cai, Z.-W.; Ge, H.-H.; Yi, Z.-W.; Zeng, R.-Y.; Zhang, G.-Y. Characterization of a Novel Psychrophilic and Halophilic β-1,3-Xylanase from Deep-Sea Bacterium, Flammeovirga pacifica Strain WPAGA1. Int. J. Biol. Macromol. 2018, 118, 2176–2184. [Google Scholar] [CrossRef]
- Le Borgne, S.; Paniagua, D.; Vazquez-Duhalt, R. Biodegradation of Organic Pollutants by Halophilic Bacteria and Archaea. J. Mol. Microbiol. Biotechnol. 2008, 15, 74–92. [Google Scholar] [CrossRef]
- Fathepure, B.Z. Recent Studies in Microbial Degradation of Petroleum Hydrocarbons in Hypersaline Environments. Front. Microbiol. 2014, 5, 173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romano, I.; Abbate, M.; Poli, A.; D’Orazio, L. Bio-Cleaning of Nitrate Salt Efflorescence on Stone Samples Using Extremophilic Bacteria. Sci. Rep. 2019, 9, 1668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomoiu, I.; Radvan, R.; Ghervase, L.; Mohanu, I.; Enache, M.; Neagu, S.; Ruginescu, R.; Cojoc, R. Cleaning of mural paintings and mortars: Review. Rev. Rom. Mat. 2020, 50, 485–492. [Google Scholar]
- Charola, A.E.; Bläuer, C. Salts in Masonry: An Overview of the Problem. Restor. Build. Monum. 2015, 21, 119–135. [Google Scholar] [CrossRef]
- Ettenauer, J.D.; Jurado, V.; Piñar, G.; Miller, A.Z.; Santner, M.; Saiz-Jimenez, C.; Sterflinger, K. Halophilic Microorganisms Are Responsible for the Rosy Discolouration of Saline Environments in Three Historical Buildings with Mural Paintings. PLoS ONE 2014, 9, e103844. [Google Scholar] [CrossRef] [PubMed]
- Parulekar-Berde, C.; Ghoble, S.S.; Salvi, S.P.; Berde, V.B. Microorganisms and Their Enzymes as Biorestoration Agents. In Microbial Biotechnology Approaches to Monuments of Cultural Heritage; Yadav, A.N., Rastegari, A.A., Gupta, V.K., Yadav, N., Eds.; Springer: Singapore, 2020; pp. 71–86. [Google Scholar] [CrossRef]
- Ranalli, G.; Alfano, G.; Belli, C.; Lustrato, G.; Colombini, M.P.; Bonaduce, I.; Zanardini, E.; Abbruscato, P.; Cappitelli, F.; Sorlini, C. Biotechnology Applied to Cultural Heritage: Biorestoration of Frescoes Using Viable Bacterial Cells and Enzymes. J. Appl. Microbiol. 2005, 98, 73–83. [Google Scholar] [CrossRef] [PubMed]
- Bosch-Roig, P.; Regidor Ros, J.L.; Estellés, R.M. Biocleaning of Nitrate Alterations on Wall Paintings by Pseudomonas Stutzeri. Int. Biodeterior. Biodegrad. 2013, 84, 266–274. [Google Scholar] [CrossRef] [Green Version]
- Bosch-Roig, P.; Ranalli, G. The Safety of Biocleaning Technologies for Cultural Heritage. Front. Microbiol. 2014, 5, 155. [Google Scholar] [CrossRef] [Green Version]
- Ranalli, G.; Zanardini, E.; Rampazzi, L.; Corti, C.; Andreotti, A.; Colombini, M.P.; Bosch-Roig, P.; Lustrato, G.; Giantomassi, C.; Zari, D.; et al. Onsite Advanced Biocleaning System for Historical Wall Paintings Using New Agar-gauze Bacteria Gel. J. Appl. Microbiol. 2019, 126, 1785–1796. [Google Scholar] [CrossRef]
- Soffritti, I.; D’Accolti, M.; Lanzoni, L.; Volta, A.; Bisi, M.; Mazzacane, S.; Caselli, E. The Potential Use of Microorganisms as Restorative Agents: An Update. Sustainability 2019, 11, 3853. [Google Scholar] [CrossRef] [Green Version]
- Ranalli, G.; Zanardini, E. Biocleaning on Cultural Heritage: New Frontiers of Microbial Biotechnologies. J. Appl. Microbiol. 2021, 131, 583–603. [Google Scholar] [CrossRef] [PubMed]
- Ruginescu, R.; Gomoiu, I.; Popescu, O.; Cojoc, R.; Neagu, S.; Lucaci, I.; Batrinescu-Moteau, C.; Enache, M. Bioprospecting for Novel Halophilic and Halotolerant Sources of Hydrolytic Enzymes in Brackish, Saline and Hypersaline Lakes of Romania. Microorganisms 2020, 8, 1903. [Google Scholar] [CrossRef] [PubMed]
- White, T.J.; Bruns, T.D.; Lee, S.B.; Taylor, J.W. Amplification and direct sequencing of fungal ribosomal RNA Genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: New York, NY, USA, 1990; pp. 315–322. [Google Scholar] [CrossRef]
- Basic Local Alignment Search Tool. Available online: https://blast.ncbi.nlm.nih.gov/Blast.cgi (accessed on 15 October 2021).
- Ventosa, A.; Quesada, E.; Rodriguez-Valera, F.; Ruiz-Berraquero, F.; Ramos-Cormenzana, A. Numerical Taxonomy of Moderately Halophilic Gram-Negative Rods. Microbiology 1982, 128, 1959–1968. [Google Scholar] [CrossRef] [Green Version]
- Anson, M.L. The Estimation of Pepsin, Trypsin, Papain, And Cathepsin with Hemoglobin. J. Gen. Physiol. 1938, 22, 79–89. [Google Scholar] [CrossRef] [PubMed]
- Cupp-Enyard, C. Sigma’s Non-Specific Protease Activity Assay—Casein as a Substrate. JoVE 2008, 19, e899. [Google Scholar] [CrossRef]
- Shirai, K.; Jackson, R.L. Lipoprotein Lipase-Catalyzed Hydrolysis of p-Nitrophenyl Butyrate. Interfacial Activation by Phospholipid Vesicles. J. Biol. Chem. 1982, 257, 1253–1258. [Google Scholar] [CrossRef]
- De Lourdes Moreno, M.; Garcia, M.T.; Ventosa, A.; Mellado, E. Characterization of Salicola sp. IC10, a Lipase- and Protease-Producing Extreme Halophile: Hydrolytic Enzymes from Extreme Halophiles. FEMS Microbiol. Ecol. 2009, 68, 59–71. [Google Scholar] [CrossRef] [PubMed]
- Miller, G.L. Use of Dinitrosalicylic Acid Reagent for Determination of Reducing Sugar. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- Botelho, D.; Wall, M.J.; Vieira, D.B.; Fitzsimmons, S.; Liu, F.; Doucette, A. Top-Down and Bottom-Up Proteomics of SDS-Containing Solutions Following Mass-Based Separation. J. Proteome Res. 2010, 9, 2863–2870. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Rohban, R.; Amoozegar, M.A.; Ventosa, A. Screening and Isolation of Halophilic Bacteria Producing Extracellular Hydrolyses from Howz Soltan Lake, Iran. J. Ind. Microbiol. Biotechnol. 2009, 36, 333–340. [Google Scholar] [CrossRef] [PubMed]
- Menasria, T.; Aguilera, M.; Hocine, H.; Benammar, L.; Ayachi, A.; Si Bachir, A.; Dekak, A.; Monteoliva-Sánchez, M. Diversity and Bioprospecting of Extremely Halophilic Archaea Isolated from Algerian Arid and Semi-Arid Wetland Ecosystems for Halophilic-Active Hydrolytic Enzymes. Microbiol. Res. 2018, 207, 289–298. [Google Scholar] [CrossRef]
- Menasria, T.; Monteoliva-Sánchez, M.; Benammar, L.; Benhadj, M.; Ayachi, A.; Hacène, H.; Gonzalez-Paredes, A.; Aguilera, M. Culturable Halophilic Bacteria Inhabiting Algerian Saline Ecosystems: A Source of Promising Features and Potentialities. World J. Microbiol. Biotechnol. 2019, 35, 132. [Google Scholar] [CrossRef] [PubMed]
- Kaitouni, L.B.D.; Anissi, J.; Sendide, K.; El Hassouni, M. Diversity of Hydrolase-Producing Halophilic Bacteria and Evaluation of Their Enzymatic Activities in Submerged Cultures. Ann. Microbiol. 2020, 70, 33. [Google Scholar] [CrossRef]
- De Lourdes Moreno, M.; Pérez, D.; García, M.; Mellado, E. Halophilic Bacteria as a Source of Novel Hydrolytic Enzymes. Life 2013, 3, 38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Annamalai, N.; Rajeswari, M.V.; Thavasi, R.; Vijayalakshmi, S.; Balasubramanian, T. Optimization, Purification and Characterization of Novel Thermostable, Haloalkaline, Solvent Stable Protease from Bacillus Halodurans CAS6 Using Marine Shellfish Wastes: A Potential Additive for Detergent and Antioxidant Synthesis. Bioprocess Biosyst. Eng. 2013, 36, 873–883. [Google Scholar] [CrossRef]
- Esakkiraj, P.; Prabakaran, G.; Maruthiah, T.; Immanuel, G.; Palavesam, A. Purification and Characterization of Halophilic Alkaline Lipase from Halobacillus sp. Proc. Natl. Acad. Sci. India Sect. B Biol. Sci. 2016, 86, 309–314. [Google Scholar] [CrossRef]
- Li, X.; Yu, H.-Y. Purification and Characterization of an Organic-Solvent-Tolerant Cellulase from a Halotolerant Isolate, Bacillus sp. L1. J. Ind. Microbiol. Biotechnol. 2012, 39, 1117–1124. [Google Scholar] [CrossRef]
- Prakash, B.; Vidyasagar, M.; Madhukumar, M.S.; Muralikrishna, G.; Sreeramulu, K. Production, Purification, and Characterization of Two Extremely Halotolerant, Thermostable, and Alkali-Stable α-Amylases from Chromohalobacter sp. TVSP 101. Process Biochem. 2009, 44, 210–215. [Google Scholar] [CrossRef]
- Bhatt, H.B.; Singh, S.P. Cloning, Expression, and Structural Elucidation of a Biotechnologically Potential Alkaline Serine Protease from a Newly Isolated Haloalkaliphilic Bacillus lehensis JO-26. Front. Microbiol. 2020, 11, 16. [Google Scholar] [CrossRef]
- Gutiérrez-Arnillas, E.; Arellano, M.; Deive, F.J.; Rodríguez, A.; Sanromán, M.Á. Unravelling the Suitability of Biological Induction for Halophilic Lipase Production by Halomonas sp. LM1C Cultures. Bioresour. Technol. 2017, 239, 368–377. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Dong, J.; Lv, X.; Wen, Y.; Chen, S. Recombinant Expression and Characterization of Two Glycoside Hydrolases from Extreme Alklinphilic Bacterium Cellulomonas bogoriensis 69B4T. AMB Express 2020, 10, 44. [Google Scholar] [CrossRef] [PubMed]
- Harwood, C.R. Bacillus subtilis and Its Relatives: Molecular Biological and Industrial Workhorses. Trends Biotechnol. 1992, 10, 247–256. [Google Scholar] [CrossRef]
- Fujinami, S.; Fujisawa, M. Industrial Applications of Alkaliphiles and Their Enzymes—Past, Present and Future. Environ. Technol. 2010, 31, 845–856. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Total Activity (U) | Total Protein (mg) | Specific Activity (U⋅mg−1) | Yield (%) | Purification Fold |
|---|---|---|---|---|---|
| Culture supernatant | 47.5 | 27.1 | 1.75 | 100 | 1 |
| Precipitated proteins | 22 | 1.9 | 11.6 | 46.3 | 6.6 |
| Sample | Total Activity (U) | Total Protein (mg) | Specific Activity (U⋅mg−1) | Yield (%) | Purification Fold |
|---|---|---|---|---|---|
| Culture supernatant | 0.175 | 10.75 | 0.016 | 100 | 1 |
| Precipitated proteins | 0.06 | 0.485 | 0.124 | 34 | 7.75 |
| Sample | Total Activity (U) | Total Protein (mg) | Specific Activity (U⋅mg−1) | Yield (%) | Purification Fold |
|---|---|---|---|---|---|
| Culture supernatant | 0.071 | 8.66 | 0.008 | 100 | 1 |
| Precipitated proteins | 0.043 | 1.33 | 0.032 | 60.5 | 4 |
| Sample | Total Activity (U) | Total Protein (mg) | Specific Activity (U⋅mg−1) | Yield (%) | Purification Fold |
|---|---|---|---|---|---|
| Culture supernatant | 29.2 | 12 | 2.43 | 100 | 1 |
| Precipitated proteins | 21 | 1.86 | 11.3 | 72 | 4.65 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ruginescu, R.; Enache, M.; Popescu, O.; Gomoiu, I.; Cojoc, R.; Batrinescu-Moteau, C.; Maria, G.; Dumbravician, M.; Neagu, S. Characterization of Some Salt-Tolerant Bacterial Hydrolases with Potential Utility in Cultural Heritage Bio-Cleaning. Microorganisms 2022, 10, 644. https://doi.org/10.3390/microorganisms10030644
Ruginescu R, Enache M, Popescu O, Gomoiu I, Cojoc R, Batrinescu-Moteau C, Maria G, Dumbravician M, Neagu S. Characterization of Some Salt-Tolerant Bacterial Hydrolases with Potential Utility in Cultural Heritage Bio-Cleaning. Microorganisms. 2022; 10(3):644. https://doi.org/10.3390/microorganisms10030644
Chicago/Turabian StyleRuginescu, Robert, Madalin Enache, Octavian Popescu, Ioana Gomoiu, Roxana Cojoc, Costin Batrinescu-Moteau, Gabriel Maria, Maria Dumbravician, and Simona Neagu. 2022. "Characterization of Some Salt-Tolerant Bacterial Hydrolases with Potential Utility in Cultural Heritage Bio-Cleaning" Microorganisms 10, no. 3: 644. https://doi.org/10.3390/microorganisms10030644