
Immunomodulatory Effects of Macrolides Considering Evidence from Human and Veterinary Medicine


Abstract
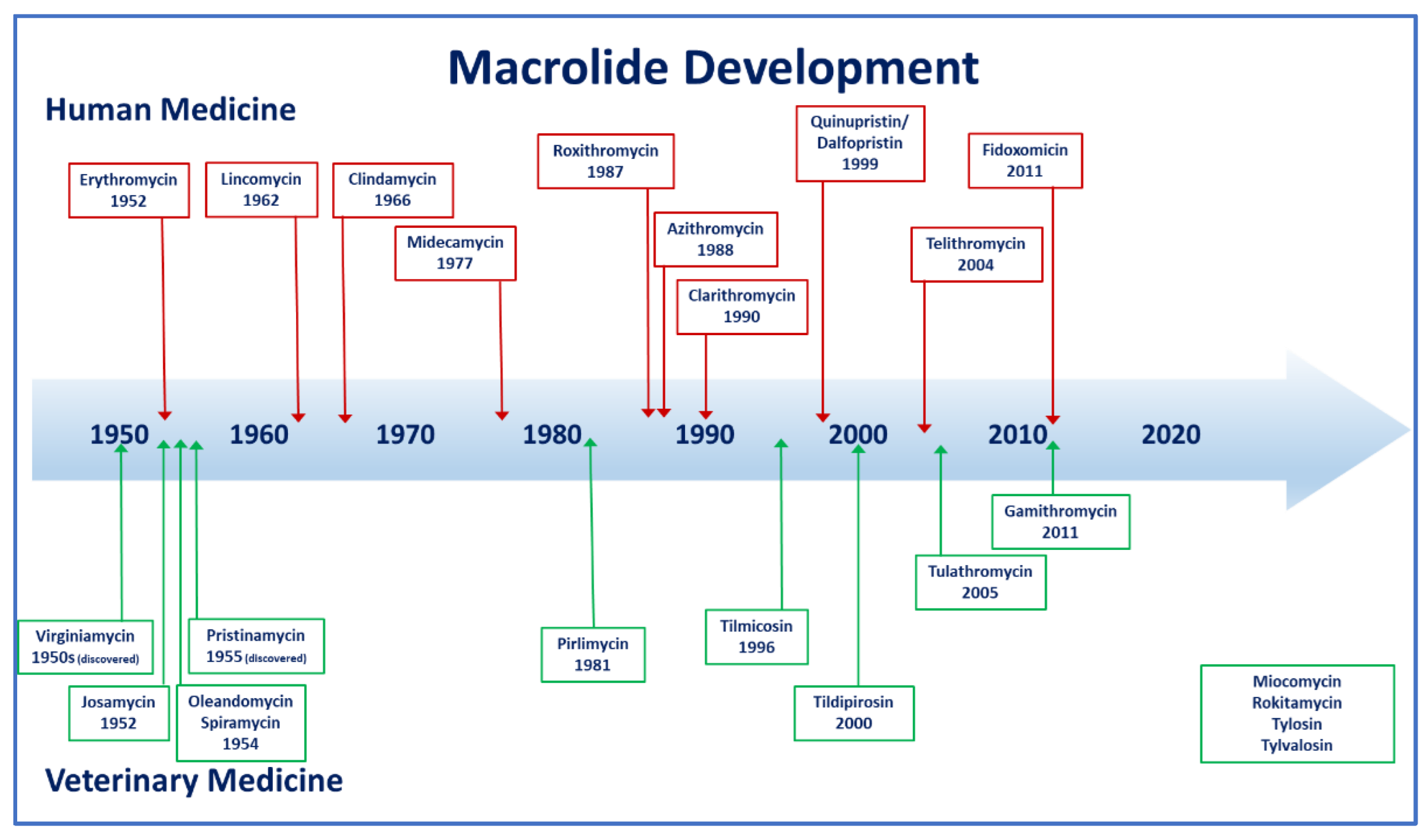
:1. Introduction
2. Classification
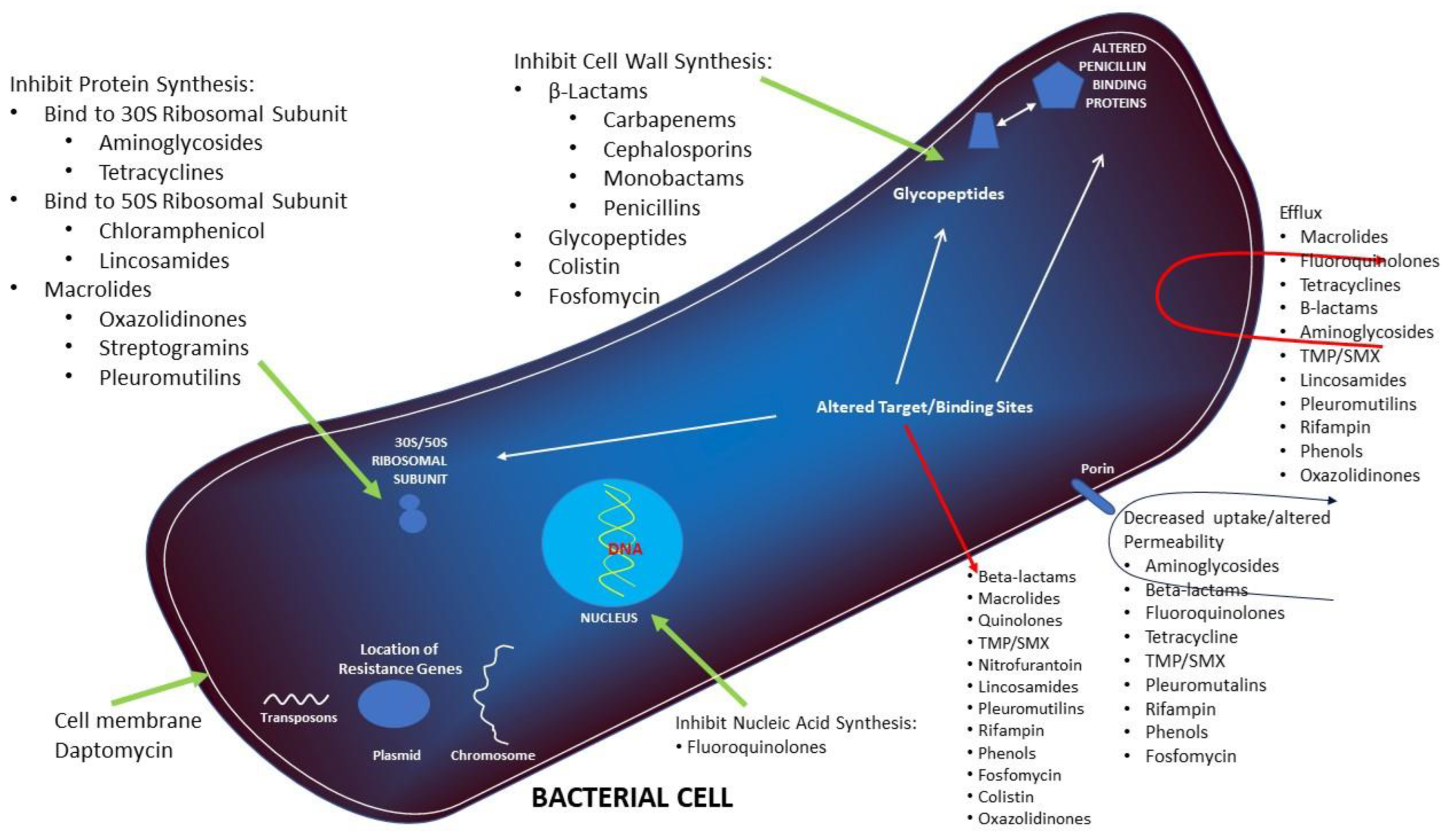
3. Mechanism of Action
4. In Vitro Activity
5. Mechanism of Resistance
6. Resistance Epidemiology
7. Pharmacokinetics
8. Pharmacodynamics
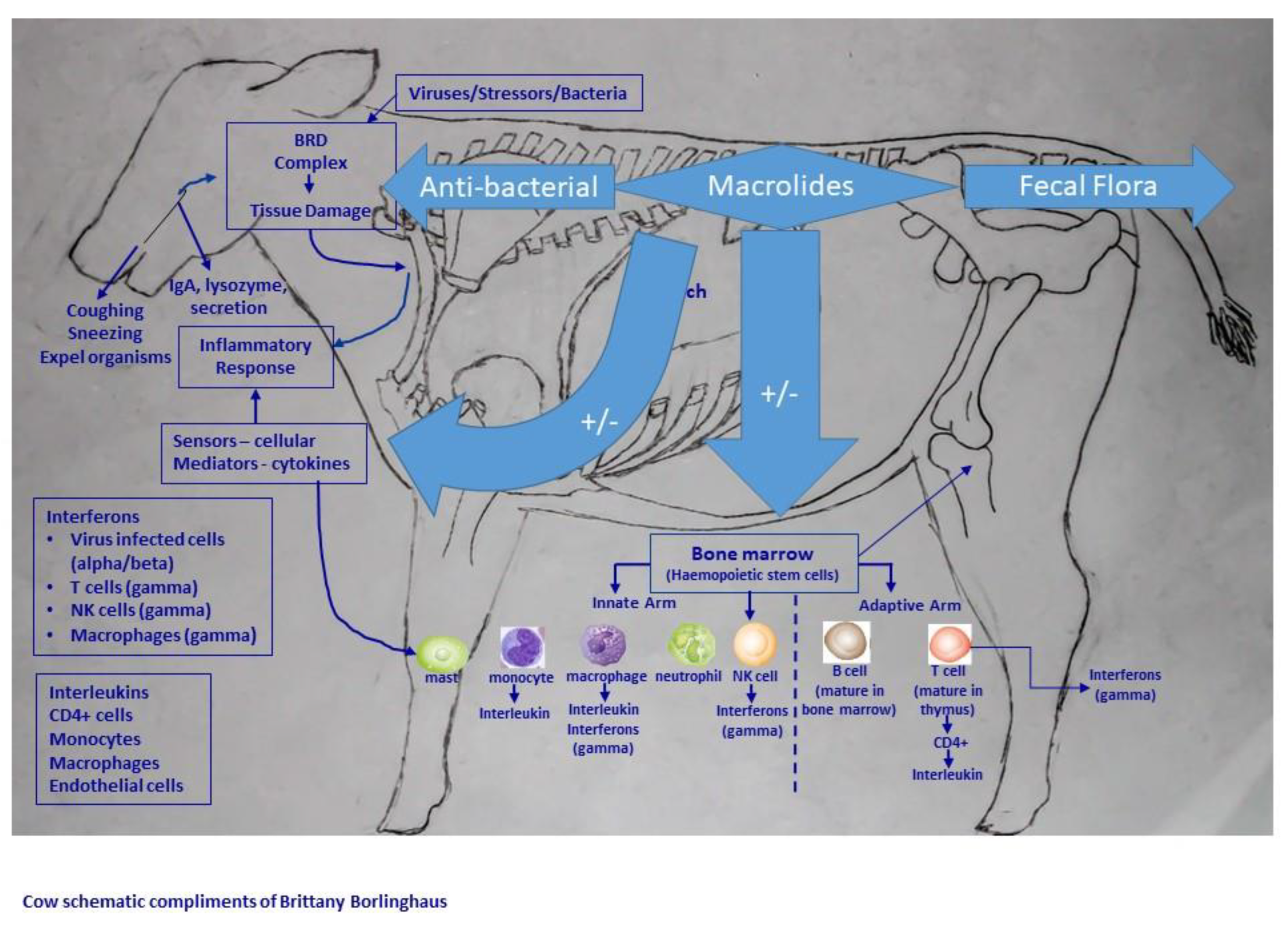
9. Inflammation
10. Anti-Inflammatory Properties
10.1. Studies in Humans
10.2. Studies in Veterinary Medicine
11. Specific Macrolide Anti-Inflammatory or Immunomodulatory Effects
- (1)
- Attenuations of biofilm function
- (2)
- Suppression of bacterial quorum sensing
- (3)
- Decrease in bacterial adherence
- (4)
- Loss of flagellar mobility
- (5)
- Reduced production of bacterial pathogens
- (6)
- Consolidation of epithelial tight junctions
- (7)
- Increasing ciliary beat frequency
- (8)
- Reduction of sputum quantity
- (9)
- Diminished sputum viscosity
- (10)
- Inhibition of synthesis of proinflammatory agents by bacteria, eosinophils, neutrophils, epithelial cells
- (11)
- Reduction of neutrophil chemotaxis
- (12)
- Stimulation of neutrophil degranulation
- (13)
- Acceleration of neutrophil apoptosis
- (14)
- Down regulation of adhesion molecules
- (15)
- Stimulation of phagocytosis by alveolar macrophages
- (16)
- Reduction in T-cell numbers and T-cell migration
- (17)
- Modulation of dendritic cell function
- (1)
- intracellular killing,
- (2)
- phagocytosis,
- (3)
- macrophage maturation,
- (4)
- chemokine release,
- (5)
- inflammasome nuclear factor-ĸB activation,
- (6)
- pro-inflammatory cytokine release,
- (7)
- neutrophil efflux,
- (8)
- apoptosis,
- (9)
- oxidative damage,
- (10)
- fibroproliferation
- (11)
- apoptosis of alveolar epithelial cells
- (1)
- biofilm and quorum sensing,
- (2)
- release of toxins and other pathogen-associated molecular patterns (PAMP),
- (3)
- phagocytosis,
- (4)
- Toll-like receptor expression and signaling,
- (5)
- intracellular killing,
- (6)
- chemokine release,
- (7)
- NETosis—neutrophil extracellular trap release,
- (8)
- chemotaxis,
- (9)
- antimicrobial peptides,
- (10)
- pro-inflammatory cytokine release,
- (11)
- tolerogenic monocyte differentiation,
- (12)
- monocyte and lymphocyte apoptosis,
- (13)
- neutrophil apoptosis,
- (14)
- release of damage-associated molecular pathogens (DAMPS),
- (15)
- efferocytosis and
- (16)
- mucous production.
12. Macrolides in Combination with Other Antimicrobials
12.1. Human Medicine
12.2. Antibiotic Combinations in Veterinary Medicine
13. Conclusions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- European Medicines Agency. Reflection Paper on the Use of Macrolides, Lincosamides and Streptogramins (MLS) in Food-Producing Animals in the European Union: Development of Resistance and Impact on Human and Animal Health. Available online: https://www.ema.europa.eu/en/documents/scientific-guideline/reflection-paper-use-macrolides-lincosamides-streptogramins-mls-food-producing-animals-european_en-0.pdf (accessed on 29 November 2021).
- McGuire, J.M.; Bunch, R.L.; Anderson, R.C.; Boaz, H.E.; Flynn, E.H.; Powell, H.M.; Smith, J.W. Ilotycin, a new antibiotic. Antibiot. Chemother. 1952, 2, 281–283. [Google Scholar]
- Dinos, G.P. The macrolide antibiotic renaissance. Br. J. Pharmacol. 2017, 174, 2967–2983. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chey, W.D.; Leontiadis, G.I.; Howden, C.W.; Moss, S.F. ACG Clinical Guideline: Treatment of Helicobacter pylori Infection. Am. J. Gastroenterol. 2017, 112, 212–239. [Google Scholar] [CrossRef] [PubMed]
- Pyörälä, S.; Baptiste, K.E.; Catry, B.; van Duijkeren, E.; Greko, C.; Moreno, M.A.; Pomba, M.C.; Rantala, M.; Ružauskas, M.; Sanders, P.; et al. Macrolides and lincosamides in cattle and pigs: Use and development of antimicrobial resistance. Vet. J. 2014, 200, 230–239. [Google Scholar] [CrossRef] [PubMed]
- Zanolari, P.; Dürr, S.; Jores, J.; Steiner, A.; Kuhnert, P. Ovine footrot: A review of current knowledge. Vet. J. 2021, 271, 105647. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, M.A. Azithromycin. Semin. Avian Exot. Pet Med. 2005, 14, 212–214. [Google Scholar] [CrossRef]
- Baptiste, K.E.; Kyvsgaard, N.C. Do antimicrobial mass medications work? A systematic review and meta-analysis of randomised clinical trials investigating antimicrobial prophylaxis or metaphylaxis against naturally occurring bovine respiratory disease. Pathog. Dis. 2017, 75, ftx083. [Google Scholar] [CrossRef] [Green Version]
- Thornsberry, C.; Marler, J.K.; Watts, J.L.; Yancey, R.J., Jr. Activity of pirlimycin against pathogens from cows with mastitis and recommendations for disk diffusion tests. Antimicrob. Agents Chemother. 1993, 37, 1122–1126. [Google Scholar] [CrossRef] [Green Version]
- Wellman, N.G.; O’Connor, A.M. Meta-analysis of treatment of cattle with bovine respiratory disease with tulathromycin. J. Vet. Pharmacol. Ther. 2007, 30, 234–241. [Google Scholar] [CrossRef]
- Nanjiani, I.A.; McKelvie, J.; Benchaoui, H.A.; Godinho, K.S.; Sherington, J.; Sunderland, S.J.; Weatherley, A.J.; Rowan, T.G. Evaluation of the therapeutic activity of tulathromycin against swine respiratory disease on farms in Europe. Vet. Ther. 2005, 6, 203–213. [Google Scholar]
- Brown, M.H.; Brightman, A.H.; Fenwick, B.W.; Rider, M.A. Infectious bovine keratoconjunctivitis: A review. J. Vet. Intern. Med. 1998, 12, 259–266. [Google Scholar] [CrossRef] [PubMed]
- Kontturi, M.; Junni, R.; Simojoki, H.; Malinen, E.; Seuna, E.; Klitgaard, K.; Kujala-Wirth, M.; Soveri, T.; Pelkonen, S. Bacterial species associated with interdigital phlegmon outbreaks in Finnish dairy herds. BMC Vet. Res. 2019, 15, 44. [Google Scholar] [CrossRef] [PubMed]
- Sifferman, R.L.; Wolffe, W.A.; Holste, J.E.; Smith, L.L.; Drag, M.D.; Yoon, S.; Kunkle, B.N.; Tessman, R.K. Field efficacy evaluation of gamithromycin for treatment of bovine respiratory disease in cattle at feedlots. Int. J. Appl. Res. Vet. Med. 2011, 9, 166–175. [Google Scholar]
- Menge, M.; Rose, M.; Bohland, C.; Zschiesche, E.; Kilp, S.; Metz, W.; Allan, M.; Röpke, R.; Nürnberger, M. Pharmacokinetics of tildipirosin in bovine plasma, lung tissue, and bronchial fluid (from live, nonanesthetized cattle). J. Vet. Pharmacol. Ther. 2012, 35, 550–559. [Google Scholar] [CrossRef]
- Yao, J.; Moellering, C.J. Antibacterial agents. In Manual of Clinical Microbiology; Murray, P.R., Baron, E.J., Jorgensen, J.H., Landry, M.L., Pfaller, M.A., Eds.; ASM Press: Washington, DC, USA, 2007; pp. 1077–1113. [Google Scholar]
- Prescott, J.F. History of Antimicrobial Usage in Agriculture: An Overview. In Antimicrobial Resistance in Bacteria of Animal Origin; Aarestrup, F.M., Ed.; ASM Press: Washington, DC, USA, 2005; pp. 19–27. [Google Scholar]
- Guardabassi, L.; Kruse, H. Principles of Prudent and Rational Use of Antimicrobials in Animals. In Guide to Antimicrobial Use in Animals; Guardabassi, L., Jensen, L.B., Kruse, H., Eds.; Blackwell Publishing Ltd.: Oxford, UK, 2008; pp. 1–12. [Google Scholar]
- Ballow, C.H.; Amsden, G.W. Azithromycin: The first azalide antibiotic. Ann. Pharmacother. 1992, 26, 1253–1261. [Google Scholar] [CrossRef] [PubMed]
- Bryskier, A.; Bergogne-Bérézin, E. Macrolides. In Antibiotic and Chemotherapy: Anti-Infective Agents and Their Use in Therapy, 9th ed.; Finch, R., Greenwood, D., Norrby, R., Whitley, R.J., Eds.; Saunders: Philadelphia, PA, USA, 2003; pp. 276–289. [Google Scholar]
- Blondeau, J.M. The evolution and role of macrolides in infectious diseases. Exp. Opin. Pharmacother. 2002, 3, 1131–1151. [Google Scholar] [CrossRef] [PubMed]
- Blondeau, J.M.; DeCarolis, E.; Metzler, K.L.; Hansen, G.T. The macrolides. Expert Opin. Investig. Drugs 2002, 11, 189–215. [Google Scholar] [CrossRef]
- Blondeau, J.M. Differential impact on macrolide compounds in the selection of macrolide nonsusceptible. Streptococcus Pneumoniae Ther. 2005, 2, 813–818. [Google Scholar]
- Blondeau, J.M. Update on the use of the macrolides for community-acquired respiratory tract infections. Therapy 2006, 3, 619–650. [Google Scholar] [CrossRef]
- Blondeau, J.M.; Chan, C.K.; Davidson, R. Judicious use of antibiotics to minimize emerging resistance: The macrolide clarithromycin as a case study. Clin. Pract. 2013, 10, 359–370. [Google Scholar] [CrossRef]
- Tsang, K.W.; Roberts, P.; Read, R.C.; Kees, F.; Wilson, R.; Cole, P.J. The concentrations of clarithromycin and its 14-hydroxy metabolite in sputum of patients with bronchiectasis following single dose oral administration. J. Antimicrob. Chemother. 1994, 33, 289–297. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.F.; Bu, M.X.; Liu, P.; Sun, J.; Liu, Y.H.; Liao, X.P. Epidemiological and PK/PD cutoff values determination and PK/PD-based dose assessment of gamithromycin against Haemophilus parasuis in piglets. BMC Vet. Res. 2020, 16, 81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfister, P.; Jenni, S.; Poehlsgaard, J.; Thomas, A.; Douthwaite, S.; Ban, N.; Böttger, E.C. The structural basis of macrolide-ribosome binding assessed using mutagenesis of 23S rRNA positions 2058 and 2059. J. Mol. Biol. 2004, 342, 1569–1581. [Google Scholar] [CrossRef] [PubMed]
- Villarino, N.; Brown, S.A.; Martín-Jiménez, T. The role of the macrolide tulathromycin in veterinary medicine. Vet. J. 2013, 198, 352–357. [Google Scholar] [CrossRef]
- Evans, N.A. Tulathromycin: An overview of a new triamilide antibiotic for livestock respiratory disease. Vet. Ther. 2005, 6, 83–95. [Google Scholar] [PubMed]
- Zeng, D.; Sun, M.; Lin, Z.; Li, M.; Gehring, R.; Zeng, Z. Pharmacokinetics and Pharmacodynamics of Tildipirosin Against Pasteurella multocida in a Murine Lung Infection Model. Front. Microbiol. 2018, 9, 1038. [Google Scholar] [CrossRef]
- Vannuffel, P.; Cocito, C. Mechanism of action of streptogramins and macrolides. Drugs 1996, 51 (Suppl. S1), 20–30. [Google Scholar] [CrossRef]
- Menninger, J.R. Functional consequences of binding macrolides to ribosomes. J. Antimicrob. Chemother. 1985, 16, 23–34. [Google Scholar] [CrossRef] [PubMed]
- Corcoran, J.W. Mode of action and resistance mechanisms of macrolides. In Macrolide Antibiotics: Chemistry, Biology, and Practice; Omura, S., Ed.; Academic Press: Orlando, FL, USA, 1984; pp. 231–259. [Google Scholar]
- Vázquez-Laslop, N.; Mankin, A.S. How Macrolide Antibiotics Work. Trends Biochem. Sci. 2018, 43, 668–684. [Google Scholar] [CrossRef]
- Blondeau, J.M.; Shebelski, S.D.; Hesje, C.K. Killing of Streptococcus pneumoniae by azithromycin, clarithromycin, erythromycin, telithromycin and gemifloxacin using drug minimum inhibitory concentrations and mutant prevention concentrations. Int. J. Antimicrob. Agents 2015, 45, 594–599. [Google Scholar] [CrossRef]
- Steigbigel, N. Macrolides and clindamycin. In Principles and Practice of Infectious Diseases, 4th ed.; Mandell, G.L., Bennett, J.E., Dolin, R., Eds.; Churchill Livingstone: New York, NY, USA, 1995; pp. 366–382. [Google Scholar]
- Hardy, D.J.; Hensey, D.M.; Beyer, J.M.; Vojtko, C.; McDonald, E.J.; Fernandes, P.B. Comparative in vitro activities of new 14-, 15- and 16-membered macrolides. Antimicrob. Agents Chemother. 1988, 32, 1710–1719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brook, I.; Wexler, H.M.; Goldstein, E.J. Antianaerobic antimicrobials: Spectrum and susceptibility testing. Clin. Microbiol. Rev. 2013, 26, 526–546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sutter, V.L.; Finegold, S.M. Susceptibility of anaerobic bacteria to 23 antimicrobial agents. Antimicrob. Agents Chemother. 1976, 10, 736–752. [Google Scholar] [CrossRef] [Green Version]
- Blondeau, J.M.; Fitch, S.D. Mutant prevention and minimum inhibitory concentration drug values for enrofloxacin, ceftiofur, florfenicol, tilmicosin and tulathromycin tested against swine pathogens Actinobacillus pleuropneumoniae, Pasteurella multocida and Streptococcus suis. PLoS ONE 2019, 14, e0210154. [Google Scholar] [CrossRef]
- Kucerova, Z.; Hradecka, H.; Nechvatalova, K.; Nedbalcova, K. Antimicrobial susceptibility of Actinobacillus pleuropneumoniae isolates from clinical outbreaks of porcine respiratory diseases. Vet. Microbiol. 2011, 150, 203–206. [Google Scholar] [CrossRef]
- El Garch, F.; de Jong, A.; Simjee, S.; Moyaert, H.; Klein, U.; Ludwig, C.; Marion, H.; Haag-Diergarten, S.; Richard-Mazet, A.; Thomas, V.; et al. Monitoring of antimicrobial susceptibility of respiratory tract pathogens isolated from diseased cattle and pigs across Europe, 2009–2012: VetPath results. Vet. Microbiol. 2016, 194, 11–22. [Google Scholar] [CrossRef]
- Vilaró, A.; Novell, E.; Enrique-Tarancón, V.; Balielles, J.; Vilalta, C.; Martinez, S.; Fraile Sauce, L.J. Antimicrobial Susceptibility Pattern of Porcine Respiratory Bacteria in Spain. Antibiotics 2020, 9, 402. [Google Scholar] [CrossRef] [PubMed]
- Zactran. Available online: www.zactran.com (accessed on 4 December 2022).
- Lunha, K.; Chumpol, W.; Samngamnim, S.; Jiemsup, S.; Assavacheep, P.; Yongkiettrakul, S. Antimicrobial Susceptibility of Streptococcus suis Isolated from Diseased Pigs in Thailand, 2018–2020. Antibiotics 2022, 11, 410. [Google Scholar] [CrossRef]
- Alhamami, T.; Chowdhury, P.R.; Gomes, N.; Carr, M.; Veltman, T.; Khazandi, M.; Mollinger, J.; Deutscher, A.T.; Turni, C.; Mahdi, L.; et al. First Emergence of resistance to macrolides and tetracycline identified in Mannheimia haemolytica and Pasteurella multocida isolates from beef feedlots in Australia. Microorganisms 2021, 9, 1322. [Google Scholar] [CrossRef]
- Anholt, R.M.; Klima, C.; Allan, N.; Matheson-Bird, H.; Schatz, C.; Ajitkumar, P.; Otto, S.J.; Peters, D.; Schmid, K.; Olson, M.; et al. Antimicrobial susceptibility of bacteria that cause bovine respiratory disease complex in Alberta, Canada. Front. Vet. Sci. 2017, 4, 207. [Google Scholar] [CrossRef] [Green Version]
- Portis, E.; Lindeman, C.; Johansen, L.; Stoltman, G. A ten-year (2000–2009) study of antimicrobial susceptibility of bacteria that cause bovine respiratory disease complex—Mannheimia haemolytica, Pasteurella multocida, and Histophilus somni—In the United States and Canada. J. Vet. Diagn. Investig. 2012, 24, 932–944. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DeDonder, K.D.; Apley, M.D.; Li, M.; Gehring, R.; Harhay, D.M.; Lubbers, B.V.; White, B.J.; Capik, S.F.; KuKanich, B.; Riviere, J.E.; et al. Pharmacokinetics and pharmacodynamics of gamithromycin in pulmonary epithelial lining fluid in naturally occurring bovine respiratory disease in multisource commingled feedlot cattle. J. Vet. Pharmacol. Ther. 2016, 39, 157–166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berghaus, L.J.; Giguère, S.; Sturgill, T.L.; Bade, D.; Malinski, T.J.; Huang, R. Plasma pharmacokinetics, pulmonary distribution, and in vitro activity of gamithromycin in foals. J. Vet. Pharmacol. Therap. 2012, 35, 59–66. [Google Scholar] [CrossRef] [PubMed]
- Goldstein, E.J.; Citron, D.M.; Merriam, C.V. Linezolid activity compared to those of selected macrolides and other agents against aerobic and anaerobic pathogens isolated from soft tissue bite infections in humans. Antimicrob. Agents Chemother. 1999, 43, 1469–1474. [Google Scholar] [CrossRef] [PubMed]
- Blondeau, J.M.; Borsos, S.; Blondeau, L.D.; Blondeau, B.J.; Hesje, C. Comparative minimum inhibitory and mutant prevention drug concentrations of enrofloxacin, ceftiofur, florfenicol, tilmicosin and tulathromycin against bovine isolates of Mannheimia haemolytica. Vet. Microbiol. 2012, 160, 85–90. [Google Scholar] [CrossRef]
- De Koster, J.; Tena, J.K.; Stegemann, M.R. Treatment of bovine respiratory disease with a single administration of tulathromycin and ketoprofen. Vet. Rec. 2022, 190, e834. [Google Scholar] [CrossRef]
- Leclercq, R.; Courvalin, P. Bacterial resistance to macrolide, lincosamide and streptogramin antibiotics by target modification. Antimicrob. Agents Chemother. 1991, 35, 1267–1272. [Google Scholar] [CrossRef] [Green Version]
- Leclercq, R.; Courvalin, P. Intrinsic and unusual resistance to macrolide, lincosamide, and streptogramin antibiotics in bacteria. Antimicrob. Agents Chemother. 1991, 35, 1273–1276. [Google Scholar] [CrossRef] [Green Version]
- Tazumi, A.; Maeda, Y.; Goldsmith, C.E.; Coulter, W.A.; Mason, C.; Millar, B.C.; McCalmont, M.; Rendall, J.; Elborn, J.S.; Matsuda, M.; et al. Molecular characterization of macrolide resistance determinants [erm(B) and mef(A)] in Streptococcus pneumoniae and viridans group streptococci (VGS) isolated from adult patients with cystic fibrosis (CF). J. Antimicrob. Chemother. 2009, 64, 501–506. [Google Scholar] [CrossRef]
- Zhanel, G.G.; Dueck, M.; Hoban, D.J.; Vercaigne, L.M.; Embil, J.M.; Gin, A.S.; Karlowsky, J.A. Review of macrolides and ketolides: Focus on respiratory tract infections. Drugs 2001, 61, 443–498. [Google Scholar] [CrossRef] [PubMed]
- Weisblum, B. Erythromycin resistance by ribosome modification. Antimicrob. Agents Chemother. 1995, 39, 577–585. [Google Scholar] [CrossRef] [Green Version]
- Allen, N.E. Biochemical mechanisms of resistance to non-cell wall antibacterial agents. Prog. Med. Chem. 1995, 32, 157–238. [Google Scholar] [CrossRef]
- O’Hara, K.; Yamamoto, K. Reaction of roxithromycin and clarithromycin with macrolide-inactivating enzymes from highly erythromycin-resistant Escherichia coli. Antimicrob. Agents Chemother. 1996, 40, 1036–1038. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McClary, D.G.; Loneragan, G.H.; Shryock, T.R.; Carter, B.L.; Guthrie, C.A.; Corbin, M.J.; Mechor, G.D. Relationship of in vitro minimum inhibitory concentrations of tilmicosin against Mannheimia haemolytica and Pasteurella multocida and in vivo tilmicosin treatment outcome among calves with signs of bovine respiratory disease. J. Am. Vet. Med. Assoc. 2011, 239, 129–135. [Google Scholar] [CrossRef] [PubMed]
- Barcia-Macay, M.; Seral, C.; Mingeot-Leclercq, M.P.; Tulkens, P.M.; Van Bambeke, F. Pharmacodynamic evaluation of the intracellular activities of antibiotics against Staphylococcus aureus in a model of THP-1 macrophages. Antimicrob. Agents Chemother. 2006, 50, 841–851. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fohner, A.E.; Sparreboom, A.; Altman, R.B.; Klein, T.E. PharmGKB summary: Macrolide antibiotic pathway, pharmacokinetics/pharmacodynamics. Pharmacogenet. Genomics 2017, 27, 164–167. [Google Scholar] [CrossRef] [Green Version]
- Madgwick, L.; Mayer, S.; Keen, P. Penetration of antibiotics into bovine neutrophils and their activity against intracellular Staphylococcus aureus. J. Antimicrob. Chemother. 1989, 24, 709–718. [Google Scholar] [CrossRef]
- Bongers, S.; Hellebrekers, P.; Leenen, L.P.H.; Koenderman, L.; Hietbrink, F. Intracellular Penetration and Effects of Antibiotics on Staphylococcus aureus Inside Human Neutrophils: A Comprehensive Review. Antibiotics 2019, 8, 54. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, S.; Hunter, L.; Qin, A.; Mann, B.J.; van Hoek, M.L. Azithromycin effectiveness against intracellular infections of Francisella. BMC Microbiol. 2010, 10, 123. [Google Scholar] [CrossRef] [Green Version]
- Modric, S.; Webb, A.I.; Derendorf, H. Pharmacokinetics and pharmacodynamics of tilmicosin in sheep and cattle. J. Vet. Pharmacol. Ther. 1998, 21, 444–452. [Google Scholar] [CrossRef]
- Modric, S.; Webb, A.I.; Davidson, M. Effect of respiratory tract disease on pharmacokinetics of tilmicosin in rats. Lab. Anim. Sci. 1999, 49, 248–253. [Google Scholar] [PubMed]
- Giguère, S.; Huang, R.; Malinski, T.J.; Dorr, P.M.; Tessman, R.K.; Somerville, B.A. Disposition of gamithromycin in plasma, pulmonary epithelial lining fluid, bronchoalveolar cells, and lung tissue in cattle. Am. J. Vet. Res. 2011, 72, 326–330. [Google Scholar] [CrossRef] [PubMed]
- Villarino, N.; Brown, S.A.; Martín-Jiménez, T. Understanding the pharmacokinetics of tulathromycin: A pulmonary perspective. J. Vet. Pharmacol. Ther. 2014, 37, 211–221. [Google Scholar] [CrossRef] [PubMed]
- Prohl, A.; Lohr, M.; Ostermann, C.; Liebler-Tenorio, E.; Berndt, A.; Schroedl, W.; Rothe, M.; Schubert, E.; Sachse, K.; Reinhold, P. Enrofloxacin and macrolides alone or in combination with rifampicin as antimicrobial treatment in a bovine model of acute Chlamydia psittaci infection. PLoS ONE 2015, 10, e0119736. [Google Scholar] [CrossRef] [PubMed]
- Westphal, J.F. Macrolide-induced clinically relevant drug interactions with cytochrome P-450A (CYP) 3A4: An update focused on clarithromycin, azithromycin and dirithromycin. Br. J. Clin. Pharmacol. 2000, 50, 285–295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zuckerman, J.M.; Qamar, F.; Bono, B.R. Review of macrolides (azithromycin, clarithromycin), ketolids (telithromycin) and glycylcyclines (tigecycline). Med. Clin. N. Am. 2011, 95, 761–791. [Google Scholar] [CrossRef]
- Furman, D.; Campisi, J.; Verdin, E.; Carrera-Bastos, P.; Targ, S.; Franceschi, C.; Ferrucci, L.; Gilroy, D.W.; Fasano, A.; Miller, G.W.; et al. Chronic inflammation in the etiology of disease across the life span. Nat. Med. 2019, 25, 1822–1832. [Google Scholar] [CrossRef]
- Fajgenbaum, D.C.; June, C.H. Cytokine Storm. N. Engl. J. Med. 2020, 383, 2255–2273. [Google Scholar] [CrossRef]
- Buret, A.G. Immuno-modulation and anti-inflammatory benefits of antibiotics: The example of tilmicosin. Can. J. Vet. Res. 2010, 74, 1–10. [Google Scholar]
- Zuckerman, J.M. Macrolides and ketolides: Azithromycin, clarithromycin and telithromycin. Infect. Dis. Clin. N. Am. 2004, 18, 621–649. [Google Scholar] [CrossRef]
- Poliak, M.S. Content of monomycin in bile and its effectiveness in inflammatory diseases and surgery of the biliary tract. Antibiotiki 1963, 8, 83–87. [Google Scholar] [PubMed]
- Parfenova, E.N.; Ryviakova, E.V. Use of erythromycin in non-specific inflammatory diseases of the urogenital system. Urologiia 1963, 28, 29–31. [Google Scholar] [PubMed]
- Gluzman, I.S. Erythromycin ointment therapy of inflammatory diseases of eyelid, conjunctiva and cornea (Clinico-experimental studies). Oftalmol. Zh. 1964, 19, 450–454. [Google Scholar] [PubMed]
- Plewig, G.; Schöpf, E. Anti-inflammatory effects of antimicrobial agents: An in vivo study. J. Invest. Dermatol. 1975, 65, 532–536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kudoh, S.; Azuma, A.; Yamamoto, M.; Izumi, T.; Ando, M. Improvement of survival in patients with diffuse panbronchiolitis treated with low-dose erythromycin. Am. J. Respir. Crit. Care Med. 1998, 157, 1829–1832. [Google Scholar] [CrossRef]
- Urban-Chmiel, R.; Wernicki, A.; Lisiecka, U.; Puchalski, A.; Dec, M.; Stegierska, D.; Kurek, L. The effect of selected macrolides on the properites of bovine leukocytes. Vet. Arhiv. 2017, 87, 571–584. [Google Scholar] [CrossRef]
- Cervin, A. The anti-inflammatory effect of erythromycin and its derivatives, with special reference to nasal polyposis and chronic sinusitis. Acta Otolaryngol. 2001, 121, 83–92. [Google Scholar] [CrossRef]
- Ichikawa, Y.; Ninomiya, H.; Koga, H.; Tanaka, M.; Kinoshita, M.; Tokunaga, N.; Yano, T.; Oizumi, K. Erythromycin reduces neutrophils and neutrophil-derived elastolytic-like activity in the lower respiratory tract of bronchiolitis patients. Am. Rev. Respir. Dis. 1992, 146, 196–203. [Google Scholar] [CrossRef]
- Mainguy-Seers, S.; Vargas, A.; Labrecque, O.; Bédard, C.; Hélie, P.; Lavoie, J.P. Randomised study of the immunomodulatory effects of azithromycin in severely asthmatic horses. Vet. Rec. 2019, 185, 143. [Google Scholar] [CrossRef]
- Chin, A.C.; Lee, W.D.; Murrin, K.A.; Morck, D.W.; Merrill, J.K.; Dick, P.; Buret, A.G. Tilmicosin induces apoptosis in bovine peripheral neutrophils in the presence or in the absence of Pasteurella haemolytica and promotes neutrophil phagocytosis by macrophages. Antimicrob. Agents Chemother. 2000, 44, 2465–2470. [Google Scholar] [CrossRef] [Green Version]
- Fujita, K.; Shimizu, T.; Majima, Y.; Sakakura, Y. Effects of macrolides on interleukin-8 secretion from human nasal epithelial cells. Eur. Arch. Otorhinolaryngol. 2000, 257, 199–204. [Google Scholar] [CrossRef] [PubMed]
- Peric, A.; Vojvodic, D.; Baletic, N.; Peric, A.; Miljanovic, O. Influence of allergy on the immunomodulatory and clinical effects of long-term low-dose macrolide treatment of nasal polyposis. Biomed. Pap. Med. Fac. Univ. Palacky Olomouc Czech Repub. 2010, 154, 327–333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perić, A.; Vojvodić, D.; Matković-Jožin, S. Effect of long-term, low-dose clarithromycin on T helper 2 cytokines, eosinophilic cationic protein and the ‘regulated on activation, normal T cell expressed and secreted’ chemokine in the nasal secretions of patients with nasal polyposis. J. Laryngol. Otol. 2012, 126, 495–502. [Google Scholar] [CrossRef] [PubMed]
- Yamada, T.; Fujieda, S.; Mori, S.; Yamamoto, H.; Saito, H. Macrolide treatment decreased the size of nasal polyps and IL-8 levels in nasal lavage. Am. J. Rhinol. 2000, 14, 143–148. [Google Scholar] [CrossRef] [PubMed]
- MacLeod, C.M.; Hamid, Q.A.; Cameron, L.; Tremblay, C.; Brisco, W. Anti-inflammatory activity of clarithromycin in adults with chronically inflamed sinus mucosa. Adv. Ther. 2001, 18, 75–82. [Google Scholar] [CrossRef]
- Cervin, A.; Wallwork, B.; Mackay-Sim, A.; Coman, W.B.; Greiff, L. Effects of long-term clarithromycin treatment on lavage-fluid markers of inflammation in chronic rhinosinusitis. Clin. Physiol. Funct. Imaging 2009, 29, 136–142. [Google Scholar] [CrossRef] [Green Version]
- Wallwork, B.; Coman, W.; Mackay-Sim, A.; Cervin, A. Effect of clarithromycin on nuclear factor-kappa B and transforming growth factor-beta in chronic rhinosinusitis. Laryngoscope 2004, 114, 286–290. [Google Scholar] [CrossRef]
- Suzuki, H.; Shimomura, A.; Ikeda, K.; Oshima, T.; Takasaka, T. Effects of long-term low-dose macrolide administration on neutrophil recruitment and IL-8 in the nasal discharge of chronic sinusitis patients. Tohoku J. Exp. Med. 1997, 182, 115–124. [Google Scholar] [CrossRef] [Green Version]
- Wallwork, B.; Coman, W.; Mackay-Sim, A.; Greiff, L.; Cervin, A. A double-blind, randomized, placebo-controlled trial of macrolide in the treatment of chronic rhinosinusitis. Laryngoscope 2006, 116, 189–193. [Google Scholar] [CrossRef]
- Fonseca-Aten, M.; Okada, P.J.; Bowlware, K.L.; Chavez-Bueno, S.; Mejias, A.; Rios, A.M.; Katz, K.; Olsen, K.; Ng, S.; Jafri, H.S.; et al. Effect of clarithromycin on cytokines and chemokines in children with an acute exacerbation of recurrent wheezing: A double-blind, randomized, placebo-controlled trial. Ann. Allergy. Asthma. Immunol. 2006, 97, 457–463. [Google Scholar] [CrossRef]
- Zhang, L.; Su, Z.; Zhang, Z.; Lin, J.; Li, D.Q.; Pflugfelder, S.C. Effects of azithromycin on gene expression profiles of proinflammatory and anti-inflammatory mediators in the eyelid margin and conjunctiva of patients with meibomian gland disease. JAMA Ophthalmol. 2015, 133, 1117–1123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gong, Y.; Lu, J.; Ding, X.; Yu, Y. Effect of adjunctive roxithromycin therapy on interleukin-1β, transforming growth factor-β1 and vascular endothelial growth factor in gingival crevicular fluid of cyclosporine A-treated patients with gingival overgrowth. J. Periodontal Res. 2014, 49, 448–457. [Google Scholar] [CrossRef] [PubMed]
- Ho, W.; Eubank, T.; Leblebicioglu, B.; Marsh, C.; Walters, J. Azithromycin decreases crevicular fluid volume and mediator content. J. Dent. Res. 2010, 89, 831–835. [Google Scholar] [CrossRef]
- Cameron, E.J.; Chaudhuri, R.; Mair, F.; McSharry, C.; Greenlaw, N.; Weir, C.J.; Jolly, L.; Donnelly, I.; Gallacher, K.; Morrison, D.; et al. Randomised controlled trial of azithromycin in smokers with asthma. Eur. Respir. J. 2013, 42, 1412–1415. [Google Scholar] [CrossRef]
- He, J.; Zhu, N.; Chen, X. Clinical impacts of azithromycin on lung function and cytokines for asthmatic patients. Fudan Univ. J. Med. Sci. 2009, 36, 719–722. [Google Scholar]
- Amayasu, H.; Yoshida, S.; Ebana, S.; Yamamoto, Y.; Nishikawa, T.; Shoji, T.; Nakagawa, H.; Hasegawa, H.; Nakabayashi, M.; Ishizaki, Y. Clarithromycin suppresses bronchial hyperresponsiveness associated with eosinophilic inflammation in patients with asthma. Ann. Allergy Asthma Immunol. 2000, 84, 594–598. [Google Scholar] [CrossRef] [PubMed]
- Simpson, J.L.; Powell, H.; Boyle, M.J.; Scott, R.J.; Gibson, P.G. Clarithromycin targets neutrophilic airway inflammation in refractory asthma. Am. J. Respir. Crit. Care Med. 2008, 177, 148–155. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Zhang, S.; Qu, Y. Effect of clarithromycin on non-eosinophilic refractory asthma. J. Clin. Pulm. Med. 2012, 17, 1948–1951. [Google Scholar]
- Shoji, T.; Yoshida, S.; Sakamoto, H.; Hasegawa, H.; Nakagawa, H.; Amayasu, H. Anti-inflammatory effect of roxithromycin in patients with aspirin-intolerant asthma. Clin. Exp. Allergy 1999, 29, 950–956. [Google Scholar] [CrossRef]
- Liu, J.; Zhong, X.; He, Z.; Wei, L.; Zheng, X.; Zhang, J.; Bai, J.; Zhong, W.; Zhong, D. Effect of low-dose, long-term roxithromycin on airway inflammation and remodeling of stable noncystic fibrosis bronchiectasis. Mediat. Inflamm. 2014, 2014, 708608. [Google Scholar] [CrossRef] [Green Version]
- Parnham, M.J.; Culić, O.; Eraković, V.; Munić, V.; Popović-Grle, S.; Barisić, K.; Bosnar, M.; Brajsa, K.; Cepelak, I.; Cuzić, S.; et al. Modulation of neutrophil and inflammation markers in chronic obstructive pulmonary disease by short-term azithromycin treatment. Eur. J. Pharmacol. 2005, 517, 132–143. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, D.; Honeybourne, D.; Khair, O.A. The effect of oral clarithromycin on bronchial airway inflammation in moderate-to-severe stable COPD: A randomized controlled trial. Treat. Respir. Med. 2004, 3, 59–65. [Google Scholar] [CrossRef] [PubMed]
- He, Z.Y.; Ou, L.M.; Zhang, J.Q.; Bai, J.; Liu, G.N.; Li, M.H.; Deng, J.M.; MacNee, W.; Zhong, X.N. Effect of 6 months of erythromycin treatment on inflammatory cells in induced sputum and exacerbations in chronic obstructive pulmonary disease. Respiration 2010, 80, 445–452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Equi, A.; Balfour-Lynn, I.M.; Bush, A.; Rosenthal, M. Long term azithromycin in children with cystic fibrosis: A randomised, placebo-controlled crossover trial. Lancet 2002, 360, 978–984. [Google Scholar] [CrossRef]
- Pukhalsky, A.L.; Shmarina, G.V.; Kapranov, N.I.; Kokarovtseva, S.N.; Pukhalskaya, D.; Kashirskaja, N.J. Anti-inflammatory and immunomodulating effects of clarithromycin in patients with cystic fibrosis lung disease. Mediat. Inflamm. 2004, 13, 111–117. [Google Scholar] [CrossRef] [Green Version]
- Criqui, G.I.; Solomon, C.; Welch, B.S.; Ferrando, R.E.; Boushey, H.A.; Balmes, J.R. Effects of azithromycin on ozone-induced airway neutrophilia and cytokine release. Eur. Respir. J. 2000, 15, 856–862. [Google Scholar] [CrossRef] [Green Version]
- Piacentini, G.L.; Peroni, D.G.; Bodini, A.; Pigozzi, R.; Costella, S.; Loiacono, A.; Boner, A.L. Azithromycin reduces bronchial hyperresponsiveness and neutrophilic airway inflammation in asthmatic children: A preliminary report. Allergy Asthma Proc. 2007, 28, 194–198. [Google Scholar] [CrossRef]
- Kraft, M.; Cassell, G.H.; Pak, J.; Martin, R.J. Mycoplasma pneumoniae and Chlamydia pneumoniae in asthma: Effect of clarithromycin. Chest 2002, 121, 1782–1788. [Google Scholar] [CrossRef]
- Yalçin, E.; Kiper, N.; Ozçelik, U.; Doğru, D.; Firat, P.; Sahin, A.; Ariyürek, M.; Mocan, G.; Gürcan, N.; Göçmen, A. Effects of claritromycin on inflammatory parameters and clinical conditions in children with bronchiectasis. J. Clin. Pharm. Ther. 2006, 31, 49–55. [Google Scholar] [CrossRef]
- Park, S.J.; Lee, Y.C.; Rhee, Y.K.; Lee, H.B. The effect of long-term treatment with erythromycin on Th1 and Th2 cytokines in diffuse panbronchiolitis. Biochem. Biophys. Res. Commun. 2004, 324, 114–117. [Google Scholar] [CrossRef]
- Oda, H.; Kadota, J.; Kohno, S.; Hara, K. Erythromycin inhibits neutrophil chemotaxis in bronchoalveoli of diffuse panbronchiolitis. Chest 1994, 106, 1116–1123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kadota, J.; Sakito, O.; Kohno, S.; Sawa, H.; Mukae, H.; Oda, H.; Kawakami, K.; Fukushima, K.; Hiratani, K.; Hara, K. A mechanism of erythromycin treatment in patients with diffuse panbronchiolitis. Am. Rev. Respir. Dis. 1993, 147, 153–159. [Google Scholar] [CrossRef] [PubMed]
- Katsuki, M. Neutrophil chemotactic activity in bronchoalveolar lavage fluid recovered from patients with diffuse panbronchiolitis. Kurume Med. J. 1996, 43, 279–287. [Google Scholar] [CrossRef] [PubMed]
- Sakito, O.; Kadota, J.; Kohno, S.; Abe, K.; Shirai, R.; Hara, K. Interleukin 1 beta, tumor necrosis factor alpha, and interleukin 8 in bronchoalveolar lavage fluid of patients with diffuse panbronchiolitis: A potential mechanism of macrolide therapy. Respiration 1996, 63, 42–48. [Google Scholar] [CrossRef]
- Doğru, D.; Dalgiç, F.; Kiper, N.; Ozçelik, U.; Yalçin, E.; Aslan, A.T.; Gürcan, N.; Saricaoğlu, F.; Gür, D.; Karayazgan, Y.; et al. Long-term clarithromycin in cystic fibrosis: Effects on inflammatory markers in BAL and clinical status. Turk. J. Pediatr. 2009, 51, 416–423. [Google Scholar] [PubMed]
- Verleden, S.E.; Vandooren, J.; Vos, R.; Willems, S.; Dupont, L.J.; Verleden, G.M.; Van Raemdonck, D.E.; Opdenakker, G.; Vanaudenaerde, B.M. Azithromycin decreases MMP-9 expression in the airways of lung transplant recipients. Transpl. Immunol. 2011, 25, 159–162. [Google Scholar] [CrossRef]
- Aubert, J.D.; Juillerat-Jeanneret, L.; Fioroni, P.; Dayer, P.; Plan, P.A.; Leuenberger, P. Function of human alveolar macrophages after a 3-day course of azithromycin in healthy volunteers. Pulm. Pharmacol. Ther. 1998, 11, 263–269. [Google Scholar] [CrossRef]
- Fouka, E.; Lamprianidou, E.; Arvanitidis, K.; Filidou, E.; Kolios, G.; Miltiades, P.; Paraskakis, E.; Antoniadis, A.; Kotsianidis, I.; Bouros, D. Low-dose clarithromycin therapy modulates Th17 response in non-cystic fibrosis bronchiectasis patients. Lung 2014, 192, 849–855. [Google Scholar] [CrossRef]
- Umeki, S. Anti-inflammatory action of erythromycin. Its inhibitory effect on neutrophil NADPH oxidase activity. Chest 1993, 104, 1191–1193. [Google Scholar] [CrossRef]
- Ratjen, F.; Saiman, L.; Mayer-Hamblett, N.; Lands, L.C.; Kloster, M.; Thompson, V.; Emmett, P.; Marshall, B.; Accurso, F.; Sagel, S.; et al. Effect of azithromycin on systemic markers of inflammation in patients with cystic fibrosis uninfected with Pseudomonas aeruginosa. Chest 2012, 142, 1259–1266. [Google Scholar] [CrossRef] [Green Version]
- Berg, H.F.; Maraha, B.; Scheffer, G.J.; Peeters, M.F.; Kluytmans, J.A. Effect of clarithromycin on inflammatory markers in patients with atherosclerosis. Clin. Diagn. Lab. Immunol. 2003, 10, 525–528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Culić, O.; Eraković, V.; Cepelak, I.; Barisić, K.; Brajsa, K.; Ferencić, Z.; Galović, R.; Glojnarić, I.; Manojlović, Z.; Munić, V.; et al. Azithromycin modulates neutrophil function and circulating inflammatory mediators in healthy human subjects. Eur. J. Pharmacol. 2002, 450, 277–289. [Google Scholar] [CrossRef] [PubMed]
- Kamoi, H.; Kurihara, N.; Fujiwara, H.; Hirata, K.; Takeda, T. The macrolide antibacterial roxithromycin reduces bronchial hyperresponsiveness and superoxide anion production by polymorphonuclear leukocytes in patients with asthma. J. Asthma 1995, 32, 191–197. [Google Scholar] [CrossRef] [PubMed]
- Kudoh, S.; Uetake, T.; Hagiwara, K.; Hirayama, M.; Hus, L.H.; Kimura, H.; Sugiyama, Y. Clinical effects of low-dose long-term erythromycin chemotherapy on diffuse panbronchiolitis. Nihon Kyobu Shikkan Gakkai Zasshi 1987, 25, 632–642. [Google Scholar]
- Crosbie, P.A.; Woodhead, M.A. Long-term macrolide therapy in chronic inflammatory airway diseases. Eur. Respir. J. 2009, 33, 171–181. [Google Scholar] [CrossRef]
- Shimizu, T.; Suzaki, H. Past, present and future of macrolide therapy for chronic rhinosinusitis in Japan. Auris. Nasus. Larynx 2016, 43, 131–136. [Google Scholar] [CrossRef]
- Spagnolo, P.; Fabbri, L.M.; Bush, A. Long-term macrolide treatment for chronic respiratory disease. Eur. Respir. J. 2013, 42, 239–251. [Google Scholar] [CrossRef]
- Imperi, F.; Leoni, L.; Visca, P. Antivirulence activity of azithromycin in Pseudomonas aeruginosa. Front. Microbiol. 2014, 5, 178. [Google Scholar] [CrossRef]
- Strausbaugh, S.D.; Davis, P.B. Cystic fibrosis: A review of epidemiology and pathobiology. Clin. Chest Med. 2007, 28, 279–288. [Google Scholar] [CrossRef]
- Saiman, L.; Marshall, B.C.; Mayer-Hamblett, N.; Burns, J.L.; Quittner, A.L.; Cibene, D.A.; Coquillette, S.; Fieberg, A.Y.; Accurso, F.J.; Campbell, P.W., 3rd. Azithromycin in patients with cystic fibrosis chronically infected with Pseudomonas aeruginosa: A randomized controlled trial. JAMA 2003, 290, 1749–1756. [Google Scholar] [CrossRef] [Green Version]
- Pirzada, O.M.; McGaw, J.; Taylor, C.J.; Everard, M.L. Improved lung function and body mass index associated with long-term use of Macrolide antibiotics. J. Cyst. Fibros. 2003, 2, 69–71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Howe, R.A.; Spencer, R.C. Macrolides for the treatment of Pseudomonas aeruginosa infections? J. Antimicrob. Chemother. 1997, 40, 153–155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carbon, C.; Poole, M.D. The role of newer macrolides in the treatment of community-acquired respiratory tract infection. A review of experimental and clinical data. J. Chemother. 1999, 11, 107–118. [Google Scholar] [CrossRef] [PubMed]
- Ishiguro, M.; Koga, H.; Kohno, S.; Hayashi, T.; Yamaguchi, K.; Hirota, M. Penetration of macrolides into human polymorphonuclear leucocytes. J. Antimicrob. Chemother. 1989, 24, 719–729. [Google Scholar] [CrossRef] [PubMed]
- Akamatsu, H.; Niwa, Y.; Sasaki, H.; Asada, Y.; Horio, T. Enhanced effect of clarithromycin on neutrophil function in vitro. J. Int. Med. Res. 1996, 24, 185–189. [Google Scholar] [CrossRef]
- Anderson, R. Erythromycin and roxithromycin potentiate human neutrophil locomotion in vitro by inhibition of leukoattractant-activated superoxide generation and autooxidation. J. Infect. Dis. 1989, 159, 966–973. [Google Scholar] [CrossRef]
- Fraschini, F.; Scaglione, F.; Ferrara, F.; Marelli, O.; Braga, P.C.; Teodori, F. Evaluation of the immunostimulating activity of erythromycin in man. Chemotherapy 1986, 32, 286–290. [Google Scholar] [CrossRef]
- Mitsuyama, T.; Tanaka, T.; Hidaka, K.; Abe, M.; Hara, N. Inhibition by erythromycin of superoxide anion production by human polymorphonuclear leukocytes through the action of cyclic AMP-dependent protein kinase. Respiration 1995, 62, 269–273. [Google Scholar] [CrossRef]
- Tanaka, E.; Kanthakumar, K.; Cundell, D.R.; Tsang, K.W.; Taylor, G.W.; Kuze, F.; Cole, P.J.; Wilson, R. The effect of erythromycin on Pseudomonas aeruginosa and neutrophil mediated epithelial damage. J. Antimicrob. Chemother. 1994, 33, 765–775. [Google Scholar] [CrossRef]
- Hirakata, Y.; Kaku, M.; Mizukane, R.; Ishida, K.; Furuya, N.; Matsumoto, T.; Tateda, K.; Yamaguchi, K. Potential effects of erythromycin on host defense systems and virulence of Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 1992, 36, 1922–1927. [Google Scholar] [CrossRef] [Green Version]
- Kita, E.; Sawaki, M.; Oku, D.; Hamuro, A.; Mikasa, K.; Konishi, M.; Emoto, M.; Takeuchi, S.; Narita, N.; Kashiba, S. Suppression of virulence factors of Pseudomonas aeruginosa by erythromycin. J. Antimicrob. Chemother. 1991, 27, 273–284. [Google Scholar] [CrossRef] [PubMed]
- Mizukane, R.; Hirakata, Y.; Kaku, M.; Ishii, Y.; Furuya, N.; Ishida, K.; Koga, H.; Kohno, S.; Yamaguchi, K. Comparative in vitro exoenzyme-suppressing activities of azithromycin and other macrolide antibiotics against Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 1994, 38, 528–533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Molinari, G.; Guzmán, C.A.; Pesce, A.; Schito, G.C. Inhibition of Pseudomonas aeruginosa virulence factors by subinhibitory concentrations of azithromycin and other macrolide antibiotics. J. Antimicrob. Chemother. 1993, 31, 681–688. [Google Scholar] [CrossRef] [PubMed]
- Molinari, G.; Paglia, P.; Schito, G.C. Inhibition of motility of Pseudomonas aeruginosa and Proteus mirabilis by subinhibitory concentrations of azithromycin. Eur. J. Clin. Microbiol. Infect. Dis. 1992, 11, 469–471. [Google Scholar] [CrossRef]
- Schneierson, S.S.; Amsterdam, D.; Perlman, E. Inhibition of Pseudomonas aeruginosa pigment formation by chloramphenicol and erythromycin. Antibiot. Chemother. 1960, 10, 30–33. [Google Scholar]
- Lesan, A.; Lamle, A.E. Short review on the diagnosis and treatment of bronchiectasis. Med. Pharm. Rep. 2019, 92, 111–116. [Google Scholar] [CrossRef]
- Cymbala, A.A.; Edmonds, L.C.; Bauer, M.A.; Jederlinic, P.J.; May, J.J.; Victory, J.M.; Amsden, G.W. The disease-modifying effects of twice-weekly oral azithromycin in patients with bronchiectasis. Treat. Respir. Med. 2005, 4, 117–122. [Google Scholar] [CrossRef]
- Davies, G.; Wilson, R. Prophylactic antibiotic treatment of bronchiectasis with azithromycin. Thorax 2004, 59, 540–541. [Google Scholar]
- Koh, Y.Y.; Lee, M.H.; Sun, Y.H.; Sung, K.W.; Chae, J.H. Effect of roxithromycin on airway responsiveness in children with bronchiectasis: A double-blind, placebo-controlled study. Eur. Respir. J. 1997, 10, 994–999. [Google Scholar] [CrossRef] [Green Version]
- Tsang, K.W.; Ho, P.I.; Chan, K.N.; Ip, M.S.; Lam, W.K.; Ho, C.S.; Yuen, K.Y.; Ooi, G.C.; Amitani, R.; Tanaka, E. A pilot study of low-dose erythromycin in bronchiectasis. Eur. Respir. J. 1999, 13, 361–364. [Google Scholar] [CrossRef]
- Banerjee, D.; Khair, O.A.; Honeybourne, D. The effect of oral clarithromycin on health status and sputum bacteriology in stable COPD. Respir. Med. 2005, 99, 208–215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilkinson, A.; Elliot, C.A.; Mawson, S.; Amrstrong, I.; Billings, C.; Diely, D.G. A randomized controlled trial to investigate the effects of a physiotherapist-led rehabilitation programme on exercise capacity and quality of life measures in patients with pulmonary hypertension. In Proceedings of the British Thoracic Society Winter Meeting, London, UK, 1 December 2007; p. A16. [Google Scholar]
- Sedaghat, A.R. Chronic Rhinosinusitis. Am. Fam. Physician 2017, 96, 500–506. [Google Scholar] [PubMed]
- Cervin, A.; Kalm, O.; Sandkull, P.; Lindberg, S. One-year low-dose erythromycin treatment of persistent chronic sinusitis after sinus surgery: Clinical outcome and effects on mucociliary parameters and nasal nitric oxide. Otolaryngol. Head Neck Surg. 2002, 126, 481–489. [Google Scholar] [CrossRef] [PubMed]
- Piccirillo, J.F.; Merritt, M.G., Jr.; Richards, M.L. Psychometric and clinimetric validity of the 20-Item Sino-Nasal Outcome Test (SNOT-20). Otolaryngol. Head Neck Surg. 2002, 126, 41–47. [Google Scholar] [CrossRef]
- Kostadima, E.; Tsiodras, S.; Alexopoulos, E.I.; Kaditis, A.G.; Mavrou, I.; Georgatou, N.; Papamichalopoulos, A. Clarithromycin reduces the severity of bronchial hyperresponsiveness in patients with asthma. Eur. Respir. J. 2004, 23, 714–717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnston, S.L.; Blasi, F.; Black, P.N.; Martin, R.J.; Farrell, D.J.; Nieman, R.B. The effect of telithromycin in acute exacerbations of asthma. N. Engl. J. Med. 2006, 354, 1589–1600. [Google Scholar] [CrossRef] [Green Version]
- Black, P.N.; Blasi, F.; Jenkins, C.R.; Scicchitano, R.; Mills, G.D.; Rubinfeld, A.R.; Ruffin, R.E.; Mullins, P.R.; Dangain, J.; Cooper, B.C.; et al. Trial of roxithromycin in subjects with asthma and serological evidence of infection with Chlamydia pneumoniae. Am. J. Respir. Crit. Care Med. 2001, 164, 536–541. [Google Scholar] [CrossRef]
- Kamada, A.K.; Hill, M.R.; Iklé, D.N.; Brenner, A.M.; Szefler, S.J. Efficacy and safety of low-dose troleandomycin therapy in children with severe, steroid-requiring asthma. J. Allergy Clin. Immunol. 1993, 91, 873–882. [Google Scholar] [CrossRef]
- Nelson, H.S.; Hamilos, D.L.; Corsello, P.R.; Levesque, N.V.; Buchmeier, A.D.; Bucher, B.L. A double-blind study of troleandomycin and methylprednisolone in asthmatic subjects who require daily corticosteroids. Am. Rev. Respir. Dis. 1993, 147, 398–404. [Google Scholar] [CrossRef]
- Thomas, D.; Gibson, P.G. Long-Term, Low-Dose Azithromycin for Uncontrolled Asthma in Children. Chest 2022, 162, 27–29. [Google Scholar] [CrossRef]
- Ghimire, J.J.; Jat, K.R.; Sankar, J.; Lodha, R.; Iyer, V.K.; Gautam, H.; Sood, S.; Kabra, S.K. Azithromycin for Poorly Controlled Asthma in Children: A Randomized Controlled Trial. Chest 2022, 161, 1456–1464. [Google Scholar] [CrossRef] [PubMed]
- Ci, X.; Chu, X.; Xiang, H.; Li, X.; Deng, X. Anti-inflammatory effects of tilmicosin in a noninfectious mouse model of allergic asthma. Immunopharmacol. Immunotoxicol. 2011, 33, 626–632. [Google Scholar] [CrossRef] [PubMed]
- Hayes, D., Jr. A review of bronchiolitis obliterans syndrome and therapeutic strategies. J. Cardiothorac. Surg. 2011, 6, 92. [Google Scholar] [CrossRef] [Green Version]
- Hakim, A.; Cooke, K.R.; Pavletic, S.Z.; Khalid, M.; Williams, K.M.; Hashmi, S.K. Diagnosis and treatment of bronchiolitis obliterans syndrome accessible universally. Bone Marrow Transplant. 2019, 54, 383–392. [Google Scholar] [CrossRef] [PubMed]
- Gerhardt, S.G.; McDyer, J.F.; Girgis, R.E.; Conte, J.V.; Yang, S.C.; Orens, J.B. Maintenance azithromycin therapy for bronchiolitis obliterans syndrome: Results of a pilot study. Am. J. Respir. Crit. Care Med. 2003, 168, 121–125. [Google Scholar] [CrossRef] [PubMed]
- Verleden, G.M.; Dupont, L.J. Azithromycin therapy for patients with bronchiolitis obliterans syndrome after lung transplantation. Transplantation 2004, 77, 1465–1467. [Google Scholar] [CrossRef] [PubMed]
- Shitrit, D.; Bendayan, D.; Gidon, S.; Saute, M.; Bakal, I.; Kramer, M.R. Long-term azithromycin use for treatment of bronchiolitis obliterans syndrome in lung transplant recipients. J. Heart Lung Transplant. 2005, 24, 1440–1443. [Google Scholar] [CrossRef]
- Khalid, M.; Al Saghir, A.; Saleemi, S.; Al Dammas, S.; Zeitouni, M.; Al Mobeireek, A.; Chaudhry, N.; Sahovic, E. Azithromycin in bronchiolitis obliterans complicating bone marrow transplantation: A preliminary study. Eur. Respir. J. 2005, 25, 490–493. [Google Scholar] [CrossRef] [Green Version]
- Verleden, G.M.; Vanaudenaerde, B.M.; Dupont, L.J.; Van Raemdonck, D.E. Azithromycin reduces airway neutrophilia and interleukin-8 in patients with bronchiolitis obliterans syndrome. Am. J. Respir. Crit. Care Med. 2006, 174, 566–570. [Google Scholar] [CrossRef]
- Yates, B.; Murphy, D.M.; Forrest, I.A.; Ward, C.; Rutherford, R.M.; Fisher, A.J.; Lordan, J.L.; Dark, J.H.; Corris, P.A. Azithromycin reverses airflow obstruction in established bronchiolitis obliterans syndrome. Am. J. Respir. Crit. Care Med. 2005, 172, 772–775. [Google Scholar] [CrossRef]
- Piccinno, M.; Lillo, E.; Rizzo, A.; Sciorsci, R.L. Unconventional effexts of anti-microbial agents in bovine reproduction. Large Anim. Rev. 2022, 28, 3–9. [Google Scholar]
- Granovsky-Grisaru, S.; Ilan, D.; Grisaru, D.; Lavie, O.; Aboulafia, I.; Diamant, Y.Z.; Hanani, M. Effects of erythromycin on contractility of isolated myometrium from pregnant rats. Am. J. Obstet. Gynecol. 1998, 178, 171–174. [Google Scholar] [CrossRef] [PubMed]
- Celik, H.; Ayar, A. Clarithromycin inhibits myometrial contractions in isolated human myometrium independent of stimulus. Physiol. Res. 2002, 51, 239–245. [Google Scholar] [PubMed]
- Abdel-Motal, S.M.; Shams, G.A.; Edress, N.E.; Anwer, A.M.; Mohamed, G.A. Immunomodulatory Effects of Tulathromycin in Rabbits. Zagazig Vet. J. 2017, 45, 1–10. [Google Scholar] [CrossRef]
- Renehan, A.G.; Booth, C.; Potten, C.S. What is apoptosis, and why is it important? BMJ 2001, 322, 1536–1538. [Google Scholar] [CrossRef] [Green Version]
- Tamaoki, J.; Kadota, J.; Takizawa, H. Clinical implications of the immunomodulatory effects of macrolides. Am. J. Med. 2004, 117 (Suppl. S9A), 5–11. [Google Scholar] [CrossRef]
- Cao, X.Y.; Dong, M.; Shen, J.Z.; Wu, B.B.; Wu, C.M.; Du, X.D.; Wang, Z.; Qi, Y.T.; Li, B.Y. Tilmicosin and tylosin have anti-inflammatory properties via modulation of COX-2 and iNOS gene expression and production of cytokines in LPS-induced macrophages and monocytes. Int. J. Antimicrob. Agents 2006, 27, 431–438. [Google Scholar] [CrossRef]
- Shinkai, M.; Henke, M.O.; Rubin, B.K. Macrolide antibiotics as immunomodulatory medications: Proposed mechanisms of action. Pharmacol. Ther. 2008, 117, 393–405. [Google Scholar] [CrossRef]
- Zimmermann, P.; Ziesenitz, V.C.; Curtis, N.; Ritz, N. The Immunomodulatory Effects of Macrolides-A Systematic Review of the Underlying Mechanisms. Front. Immunol. 2018, 9, 302. [Google Scholar] [CrossRef] [Green Version]
- Otsuka, M.; Nishi, Y.; Tsukano, K.; Tsuchiya, M.; Suzuki, K. The 15-membered ring macrolide tulathromycin inhibits plasma endotoxin activity in endotoxemic calves. In Proceedings of the Fifty-First Annual Conference, American Association of Bovine Practitioners, Phoenix, AZ, USA, 13–15 September 2018. [Google Scholar]
- Cavaillon, J.M. Exotoxins and endotoxins: Inducers of inflammatory cytokines. Toxicon 2018, 149, 45–53. [Google Scholar] [CrossRef]
- Er, A.; Yazar, E. Effects of tylosin, tilmicosin and tulathromycin on inflammatory mediators in bronchoalveolar lavage fluid of lipopolysaccharide-induced lung injury. Acta Vet. Hung. 2012, 60, 465–476. [Google Scholar] [CrossRef] [PubMed]
- Couëtil, L.L.; Cardwell, J.M.; Gerber, V.; Lavoie, J.P.; Léguillette, R.; Richard, E.A. Inflammatory Airway Disease of Horses--Revised Consensus Statement. J. Vet. Intern. Med. 2016, 30, 503–515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morck, D.W.; Merrill, J.K.; Gard, M.S.; Olson, M.E.; Nation, P.N. Treatment of experimentally induced pneumonic pasteurellosis of young calves with tilmicosin. Can. J. Vet. Res. 1997, 61, 187–192. [Google Scholar] [PubMed]
- Martínez-Cortés, I.; Acevedo-Domínguez, N.A.; Olguin-Alor, R.; Cortés-Hernández, A.; Álvarez-Jiménez, V.; Campillo-Navarro, M.; Sumano-López, H.S.; Gutiérrez-Olvera, L.; Martínez-Gómez, D.; Maravillas-Montero, J.L.; et al. Tilmicosin modulates the innate immune response and preserves casein production in bovine mammary alveolar cells during Staphylococcus aureus infection. J. Anim. Sci. 2019, 97, 644–656. [Google Scholar] [CrossRef] [PubMed]
- Tucureanu, M.M.; Rebleanu, D.; Constantinescu, C.A.; Deleanu, M.; Voicu, G.; Butoi, E.; Calin, M.; Manduteanu, I. Lipopolysaccharide-induced inflammation in monocytes/macrophages is blocked by liposomal delivery of G(i)-protein inhibitor. Int. J. Nanomed. 2018, 13, 63–76. [Google Scholar] [CrossRef] [Green Version]
- Reuter, R.R.; Carroll, J.A.; Dailey, J.W.; Cook, B.J.; Galyean, M.L. Effects of dietary energy source and level and injection of tilmicosin phosphate on immune function in lipopolysaccharide-challenged beef steers. J. Anim. Sci. 2008, 86, 1963–1976. [Google Scholar] [CrossRef]
- Kegley, E.B.; Ball, J.J.; Beck, P.A. BILL E. KUNKLE INTERDISCIPLINARY BEEF SYMPOSIUM: Impact of mineral and vitamin status on beef cattle immune function and health. J. Anim. Sci. 2016, 94, 5401–5413. [Google Scholar] [CrossRef]
- Ingvartsen, K.L.; Moyes, K. Nutrition, immune function and health of dairy cattle. Anim. Int. J. Anim. Biosci. 2013, 7 (Suppl. S1), 112–122. [Google Scholar] [CrossRef] [Green Version]
- Moges, R.; De Lamache, D.D.; Sajedy, S.; Renaux, B.S.; Hollenberg, M.D.; Muench, G.; Abbott, E.M.; Buret, A.G. Anti-Inflammatory Benefits of Antibiotics: Tylvalosin Induces Apoptosis of Porcine Neutrophils and Macrophages, Promotes Efferocytosis, and Inhibits Pro-Inflammatory CXCL-8, IL1α, and LTB(4) Production, While Inducing the Release of Pro-Resolving Lipoxin A(4) and Resolvin D1. Front. Vet. Sci. 2018, 5, 57. [Google Scholar] [CrossRef] [Green Version]
- Somacarrera, M.L.; Hernández, G.; Acero, J.; Moskow, B.S. Factors related to the incidence and severity of cyclosporin-induced gingival overgrowth in transplant patients. A longitudinal study. J. Periodontol. 1994, 65, 671–675. [Google Scholar] [CrossRef]
- Pernu, H.E.; Pernu, L.M.; Huttunen, K.R.; Nieminen, P.A.; Knuuttila, M.L. Gingival overgrowth among renal transplant recipients related to immunosuppressive medication and possible local background factors. J. Periodontol. 1992, 63, 548–553. [Google Scholar] [CrossRef] [PubMed]
- Lewis, J.R.; Reiter, A.M. Management of generalized gingival enlargement in a dog--case report and literature review. J. Vet. Dent. 2005, 22, 160–169. [Google Scholar] [CrossRef] [PubMed]
- Williamson, M.S.; Miller, E.K.; Plemons, J.; Rees, T.; Iacopino, A.M. Cyclosporine A upregulates interleukin-6 gene expression in human gingiva: Possible mechanism for gingival overgrowth. J. Periodontol. 1994, 65, 895–903. [Google Scholar] [CrossRef] [PubMed]
- Schincaglia, G.P.; Forniti, F.; Cavallini, R.; Piva, R.; Calura, G.; del Senno, L. Cyclosporin-A increases type I procollagen production and mRNA level in human gingival fibroblasts in vitro. J. Oral Pathol. Med. 1992, 21, 181–185. [Google Scholar] [CrossRef] [PubMed]
- Nash, M.M.; Zaltzman, J.S. Efficacy of azithromycin in the treatment of cyclosporine-induced gingival hyperplasia in renal transplant recipients. Transplantation 1998, 65, 1611–1615. [Google Scholar] [CrossRef] [PubMed]
- Ljutić, D.; Rumboldt, Z. Possible interaction between azithromycin and cyclosporin: A case report. Nephron 1995, 70, 130. [Google Scholar] [CrossRef]
- Clementini, M.; Vittorini, G.; Crea, A.; Gualano, M.R.; Macrì, L.A.; Deli, G.; La Torre, G. Efficacy of AZM therapy in patients with gingival overgrowth induced by Cyclosporine A: A systematic review. BMC Oral Health 2008, 8, 34. [Google Scholar] [CrossRef] [Green Version]
- Rosenberg, A.; Rosenkrantz, W.; Griffin, C.; Angus, J.; Keys, D. Evaluation of azithromycin in systemic and toothpaste forms for the treatment of ciclosporin-associated gingival overgrowth in dogs. Vet. Dermatol. 2013, 24, 337-e75. [Google Scholar] [CrossRef]
- Diesel, A.; Moriello, K. Medical Management of Cyclosporine-Induced Gingival Overgrowth Using Oral Azithromycin in Six Dogs. Vet. Sci. 2015, 2, 13–22. [Google Scholar] [CrossRef]
- Kim, J.Y.; Park, S.H.; Cho, K.S.; Kim, H.J.; Lee, C.K.; Park, K.K.; Choi, S.H.; Chung, W.Y. Mechanism of azithromycin treatment on gingival overgrowth. J. Dent. Res. 2008, 87, 1075–1079. [Google Scholar] [CrossRef]
- Condé, S.A.; Bastos, M.G.; Vieira, B.J.; Aarestrup, F.M. Down-regulation of transforming growth factor beta-2 expression is associated with the reduction of cyclosporin induced gingival overgrowth in rats treated with roxithromycin: An experimental study. BMC Oral Health 2009, 9, 33. [Google Scholar] [CrossRef]
- Matzneller, P.; Krasniqi, S.; Kinzig, M.; Sörgel, F.; Hüttner, S.; Lackner, E.; Müller, M.; Zeitlinger, M. Blood, tissue, and intracellular concentrations of azithromycin during and after end of therapy. Antimicrob. Agents Chemother. 2013, 57, 1736–1742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altenburg, J.; de Graaff, C.S.; van der Werf, T.S.; Boersma, W.G. Immunomodulatory effects of macrolide antibiotics—Part 1: Biological mechanisms. Respiration 2011, 81, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Sauer, A.; Peukert, K.; Putensen, C.; Bode, C. Antibiotics as immunomodulators: A potential pharmacologic approach for ARDS treatment. Eur. Respir. Rev. Off. J. Eur. Respir. Soc. 2021, 30, 210093. [Google Scholar] [CrossRef] [PubMed]
- Reijnders, T.D.Y.; Saris, A.; Schultz, M.J.; van der Poll, T. Immunomodulation by macrolides: Therapeutic potential for critical care. Lancet Respir. Med. 2020, 8, 619–630. [Google Scholar] [CrossRef]
- König, R.; Cao, X.; Oswald, M.; Forstner, C.; Rohde, G.; Rupp, J.; Witzenrath, M.; Welte, T.; Kolditz, M.; Pletz, M. Macrolide combination therapy for patients hospitalised with community-acquired pneumonia? An individualised approach supported by machine learning. Eur. Respir. J. 2019, 54, 1900824. [Google Scholar] [CrossRef]
- Metlay, J.P.; Waterer, G.W.; Long, A.C.; Anzueto, A.; Brozek, J.; Crothers, K.; Cooley, L.A.; Dean, N.C.; Fine, M.J.; Flanders, S.A.; et al. Diagnosis and Treatment of Adults with Community-acquired Pneumonia. An Official Clinical Practice Guideline of the American Thoracic Society and Infectious Diseases Society of America. Am. J. Respir. Crit. Care Med. 2019, 200, e45–e67. [Google Scholar] [CrossRef]
- Ito, A.; Ishida, T.; Tachibana, H.; Tokumasu, H.; Yamazaki, A.; Washio, Y. Azithromycin combination therapy for community-acquired pneumonia: Propensity score analysis. Sci. Rep. 2019, 9, 18406. [Google Scholar] [CrossRef] [Green Version]
- Bonne, S.L.; Kadri, S.S. Evaluation and Management of Necrotizing Soft Tissue Infections. Infect. Dis. Clin. N. Am. 2017, 31, 497–511. [Google Scholar] [CrossRef]
- Bernardo, K.; Pakulat, N.; Fleer, S.; Schnaith, A.; Utermohlen, O.; Krut, O.; Muller, S.; Kronke, M. Subinhibitory concentrations of linezolid reduce Staphylococcus aureus virulence factor expression. Antimicrob. Agents Chemother. 2004, 48, 546–555. [Google Scholar] [CrossRef] [Green Version]
- Hodille, E.; Badiou, C.; Bouveyron, C.; Bes, M.; Tristan, A.; Vandenesch, F.; Lina, G.; Dumitrescu, O. Clindamycin suppresses virulence expression in inducible clindamycin-resistant Staphylococcus aureus strains. Ann. Clin. Microbiol. Antimicrob. 2018, 17, 38. [Google Scholar] [CrossRef] [PubMed]
- Vanrompay, D.; Harkinezhad, T.; van de Walle, M.; Beeckman, D.; van Droogenbroeck, C.; Verminnen, K.; Leten, R.; Martel, A.; Cauwerts, K. Chlamydophila psittaci transmission from pet birds to humans. Emerg. Infect. Dis. 2007, 13, 1108–1110. [Google Scholar] [CrossRef] [PubMed]
- Heddema, E.R.; van Hannen, E.J.; Duim, B.; de Jongh, B.M.; Kaan, J.A.; van Kessel, R.; Lumeij, J.T.; Visser, C.E.; Vandenbroucke-Grauls, C. An outbreak of psittacosis due to Chlamydophila psittaci genotype A in a veterinary teaching hospital. J. Med. Microbiol. 2006, 55, 1571–1575. [Google Scholar] [CrossRef]
- Yu, Y.; Fang, J.T.; Zheng, M.; Zhang, Q.; Walsh, T.R.; Liao, X.P.; Sun, J.; Liu, Y.H. Combination Therapy Strategies Against Multiple-Resistant Streptococcus Suis. Front. Pharmacol. 2018, 9, 489. [Google Scholar] [CrossRef]
- Sweeney, M.T.; Brumbaugh, G.W.; Watts, J.L. In vitro activities of tulathromycin and ceftiofur combined with other antimicrobial agents using bovine Pasteurella multocida and Mannheimia haemolytica isolates. Vet. Ther. 2008, 9, 212–222. [Google Scholar] [PubMed]
- Booker, C.W.; Perrett, T.; Fenton, R.K.; Behlke, E.J.; Hannon, S.J.; Wildman, B.K.; Rademacher, R.D.; McMullen, C.A.; Hunsaker, B.D.; Hill, T.; et al. An evaluation of concomitant therapy for treatment of arrival fever in feedlot calves at ultra-high risk of developing undifferentiated fever/bovine respiratory disease. Int. J. Appl. Res. Vet. Med. 2017, 15, 85–98. [Google Scholar]
- Dudek, K.; Bednarek, D.; Ayling, R.D.; Kycko, A.; Reichert, M. Preliminary study on the effects of enrofloxacin, flunixin meglumine and pegbovigrastim on Mycoplasma bovis pneumonia. BMC Vet. Res. 2019, 15, 371. [Google Scholar] [CrossRef] [Green Version]
- Coetzee, J.F.; Magstadt, D.R.; Sidhu, P.K.; Follett, L.; Schuler, A.M.; Krull, A.C.; Cooper, V.L.; Engelken, T.J.; Kleinhenz, M.D.; O’Connor, A.M. Association between antimicrobial drug class for treatment and retreatment of bovine respiratory disease (BRD) and frequency of resistant BRD pathogen isolation from veterinary diagnostic laboratory samples. PLoS ONE 2019, 14, e0219104. [Google Scholar] [CrossRef] [Green Version]
- Espinasse, J.; Allairae, R.; Raynaud, J.P.; Van Gool, F.; Tixier, G. Combined corticorsteroid and antibiotic therapy in bovine respiratory disease (BRD) of young cattle: Clinical results. Bov. Pract. 1986, 21, 59–61. [Google Scholar]
- Llor, C.; Bjerrum, L. Antimicrobial resistance: Risk associated with antibiotic overuse and initiatives to reduce the problem. Ther. Adv. Drug Saf. 2014, 5, 229–241. [Google Scholar] [CrossRef] [Green Version]
- Bosnar, M.; Kragol, G.; Koštrun, S.; Vujasinović, I.; Bošnjak, B.; Bencetić Mihaljević, V.; Marušić Ištuk, Z.; Kapić, S.; Hrvačić, B.; Brajša, K.; et al. N′-substituted-2′-O,3′-N-carbonimidoyl bridged macrolides: Novel anti-inflammatory macrolides without antimicrobial activity. J. Med. Chem. 2012, 55, 6111–6123. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Drug | Indications | Organism on Label |
|---|---|---|
| Pirlimycin | Mastitis in cattle [9] | S. aureus S. agalactiae S. dysagalactiae S. uberis |
| Tulathromycin | BRD * [10] | M. haemolytica P. multocida H. somni M. bovis |
| SRD * [11] | A. pleuropneumoniae H. parasuis P. multocida B. bronchiseptica | |
| IBKC * [12] | Moraxella bovis | |
| Bovine Foot Rot [13] | Fusobacterium necrophorum, Porphyromonas levii | |
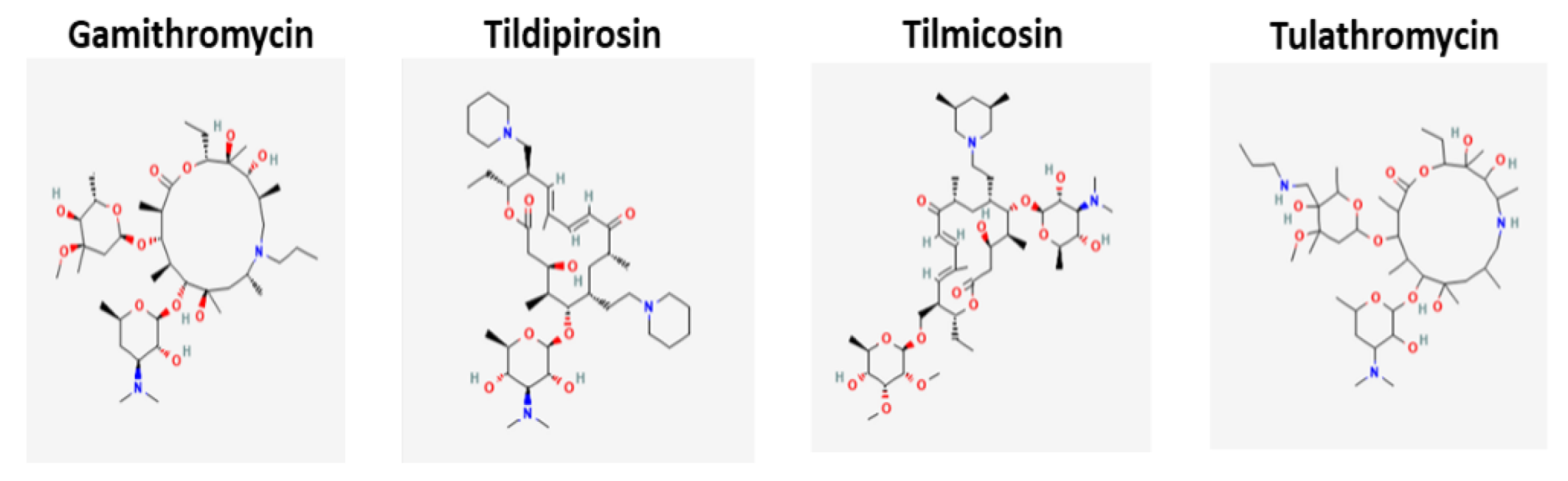
| Gamithromycin | BRD [14] | M. haemolytica P. multocida H. somni M. bovis |
| Tildipirosin | BRD [15] | M. haemolytica P. multocida H. somni |
| Tilmicosin | BRD | M. haemolytica P. multocida H. somni |
| Tylosin | BRC | P. multocida A. pyogenes |
| Foot rot/calf diphtheria | F. necrophorum | |
| Metritis | A. pyogenes | |
| Swine arthritis | M. hyosynoviae | |
| Swine pneumonia | Pasteurella spp. | |
| Swine erysipelas | E. rhusiopathiae | |
| Swine dysentery | T. hyodysenteriae |
| Gamithromycin | Tildipirosin | Tilmicosin | Tulathromycin | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Organism | No. | MIC50/90 | Range | No. | MIC50/90 | Range | No. | MIC50/90 | Range | No. | MIC50/90 | Range |
| A. pleuropneumoniae [41] | 67 | 2/4 | 1–8 | 67 | 1/2 | 0.5–8 | ||||||
| Swine [42] | 242 | 4/8 | 1–256 | 242 | 8/16 | 1–32 | ||||||
| Swine 2009–2012 [43] | 158 | 16/16 | 2–32 | 158 | 32/32 | 4–64 | ||||||
| Swine 2017–2019 [44] | 162 | 4/8 | 1–64 | 162 | 8/16 | 4–64 | 162 | 32/64 | 8–64 | |||
| P. multocida (swine) [41,45] | 73 | 2/4 | 1–8 | 73 | 0.25/0.5 | 0.063–1 | ||||||
| 2009–2012 | 152 | 8/16 | 1–32 | 152 | 2/4 | 1–8 | ||||||
| 2017–2019 [44] | 130 | 1/2 | 0.5–64 | 130 | 8/16 | 1–64 | 130 | 2/4 | 0.5–65 | |||
| S. suis [41,45] | 59 | ≥8/≥8 | ≥8 | 59 | ≥8/≥8 | ≥8 | ||||||
| 2018–2022 [46] | 246 | 128/128 | 4–128 | 246 | 64/64 | 1–64 | ||||||
| Swine 2009–2012 | 152 | >64/>64 | 0.5–>64 | 151 | >64/>64 | 1–>64 | ||||||
| M. haemolytica [45] | 89 | 1/1 | 0.5–>32 | |||||||||
| 2014–2018 [47] | 55 | 0.5/1 | 1–2 | 55 | 1/1 | 1–2 | 55 | 4/8 | 4–16 | 55 | 8/16 | 1–16 |
| 2019 [47] | 33 | 1/2 | 1–16 | 33 | 1/1 | 1–4 | 33 | 4/8 | 2–32 | 33 | 8/8 | 8–128 |
| 2014–2015 [48] | 251 | 16/64 | 4–64 | 251 | 16/64 | 1–64 | ||||||
| 2009 [49] | 304 | 8/64 | 1–>64 | 304 | 8/32 | 0.5–>64 | ||||||
| 2009–2012 | 149 | 0.5/1 | 0.12–1 | 149 | 8/16 | 1–64 | 149 | 4/8 | 1–128 | |||
| P. multocida (bovine) | 79 | 0.5/1 | 0.12–>32 | |||||||||
| 2014–2018 [47] | 75 | 1/8 | 1–16 | 75 | 1/8 | 1–16 | 75 | 4/8 | 4–64 | 75 | 1/8 | 0.5–128 |
| 2019 [47] | 65 | 1/16 | 1–16 | 65 | 1/2 | 1–16 | 65 | 8/32 | 2–32 | 65 | 8/64 | 8–64 |
| 2014–2015 [48] | 118 | 16/64 | 4–64 | 118 | 4/64 | 1–64 | ||||||
| 2009 [49] | 328 | 8/>64 | 0.5–>64 | 328 | 2/16 | 0.5–>64 | ||||||
| 2009–2012 | 134 | 0.25 | 0.06–2 | |||||||||
| H. somni [50] | 32 | 0.5/0.5 | 0.25–1 | |||||||||
| 2014–2015 [48] | 80 | 4/≥64 | 4–≥64 | 80 | 8/≥64 | 1–≥64 | ||||||
| 2009 [49] | 174 | 8/64 | 2–>64 | 174 | 8/>64 | 1–>64 | ||||||
| 2009–2012 | 66 | 0.5/0.5 | 0.25–1 | 66 | 4/8 | 0.5–16 | 66 | 4/8 | 0.12–8 | |||
| R. equi | ||||||||||||
| Macrolide S [51] | 1/1 | 0.5–1 | ||||||||||
| Macrolide R [51] | 30 | 64/128 | 32–128 | |||||||||
| S. zooepidemicus | 22 | 0.06/0.125 | 0.03–0.125 | |||||||||
| B. bronchiseptica (Swine) | ||||||||||||
| 2009–2012 | 118 | 32/32 | 8–64 | 118 | 8/8 | 1–32 | ||||||
| 2017–2019 [44] | 29 | 4/8 | 29 | 32/64 | 29 | 8/8 | ||||||
| M. bovis | ||||||||||||
| 2014–2015 [48] | 234 | 64/64 | 4–64 | 234 | 64/64 | 1–64 | ||||||
| T. pyogenes | ||||||||||||
| 2014–2015 [48] | 94 | 64/64 | 4–≥64 | 94 | 64/64 | 1–≥64 | ||||||
| H. parasuis (Swine) | ||||||||||||
| 2009–2012 | 68 | 1/2 | 0.12–4 | 68 | ½ | 0.06–4 | ||||||
| Marker | Function | Macrolide Impact |
|---|---|---|
| Total cell count |
|
|
| Leukocyte count |
|
|
| Neutrophil count |
|
|
| Neutrophil oxidative burst |
|
|
| Neutrophil chemotaxis |
|
|
| Neutrophil lactoferrin |
|
|
| Neutrophil elastase |
|
|
| Mucus |
|
|
| Neutrophil apoptosis |
| |
| Macrophage count |
|
|
| Eosinophil count |
|
|
| Eosinophil cationic protein |
|
|
| Thrombocyte count |
|
|
| IL-1beta |
|
|
| IL-2 |
|
|
| IL-4 |
|
|
| IL-5 |
|
|
| IL-6 |
|
|
| IL-8 |
|
|
| IL-12 |
|
|
| IL-17 |
|
|
| TNF- alpha |
|
|
| Lymphocytes |
|
|
| IFN-gamma |
|
|
| TGF-beta |
|
|
| Monocytes |
|
|
| GM-CSF |
|
|
| VEGF |
|
|
| Matrix metalloproteinase-9 |
|
|
| E-selectin |
|
|
| C-reactive protein |
|
|
| Serum amyloid A |
|
|
| ICAM-1 (C054) (Intracellular adhesion molecule) |
|
|
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Blondeau, J.M. Immunomodulatory Effects of Macrolides Considering Evidence from Human and Veterinary Medicine. Microorganisms 2022, 10, 2438. https://doi.org/10.3390/microorganisms10122438
Blondeau JM. Immunomodulatory Effects of Macrolides Considering Evidence from Human and Veterinary Medicine. Microorganisms. 2022; 10(12):2438. https://doi.org/10.3390/microorganisms10122438
Chicago/Turabian StyleBlondeau, Joseph M. 2022. "Immunomodulatory Effects of Macrolides Considering Evidence from Human and Veterinary Medicine" Microorganisms 10, no. 12: 2438. https://doi.org/10.3390/microorganisms10122438