
Bifidobacterium longum, Lactobacillus plantarum and Pediococcus acidilactici Reversed ETEC-Inducing Intestinal Inflammation in Mice
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains Preparation
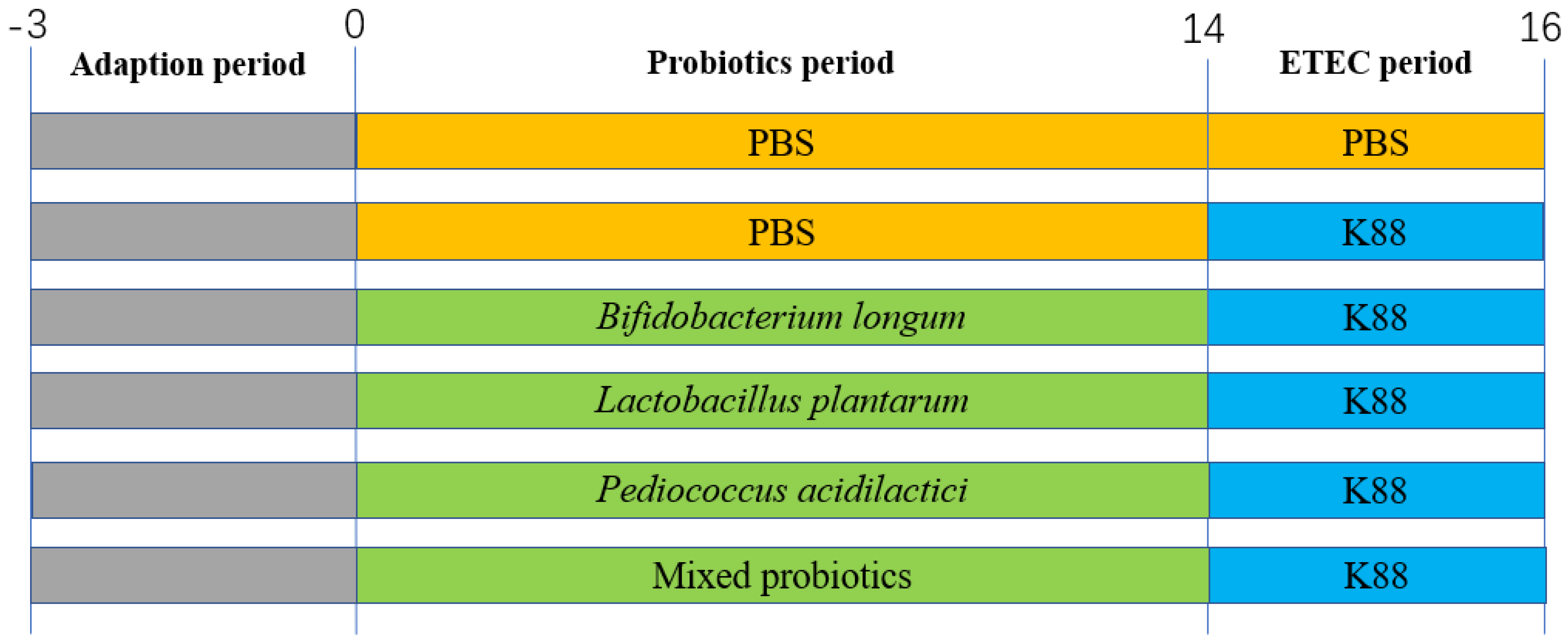
2.2. Animal Experimental Design
2.3. Sample Collection and Treatment
2.4. Detection of Inflammatory Cytokines and Immunoglobulin in Mice Serum
2.5. Intestinal Morphology Analysis
2.6. RNA Extraction and Quantitative Real-Time PCR
2.7. Microbial Analysis of Colon Contents
2.8. Statistical Analysis
3. Results
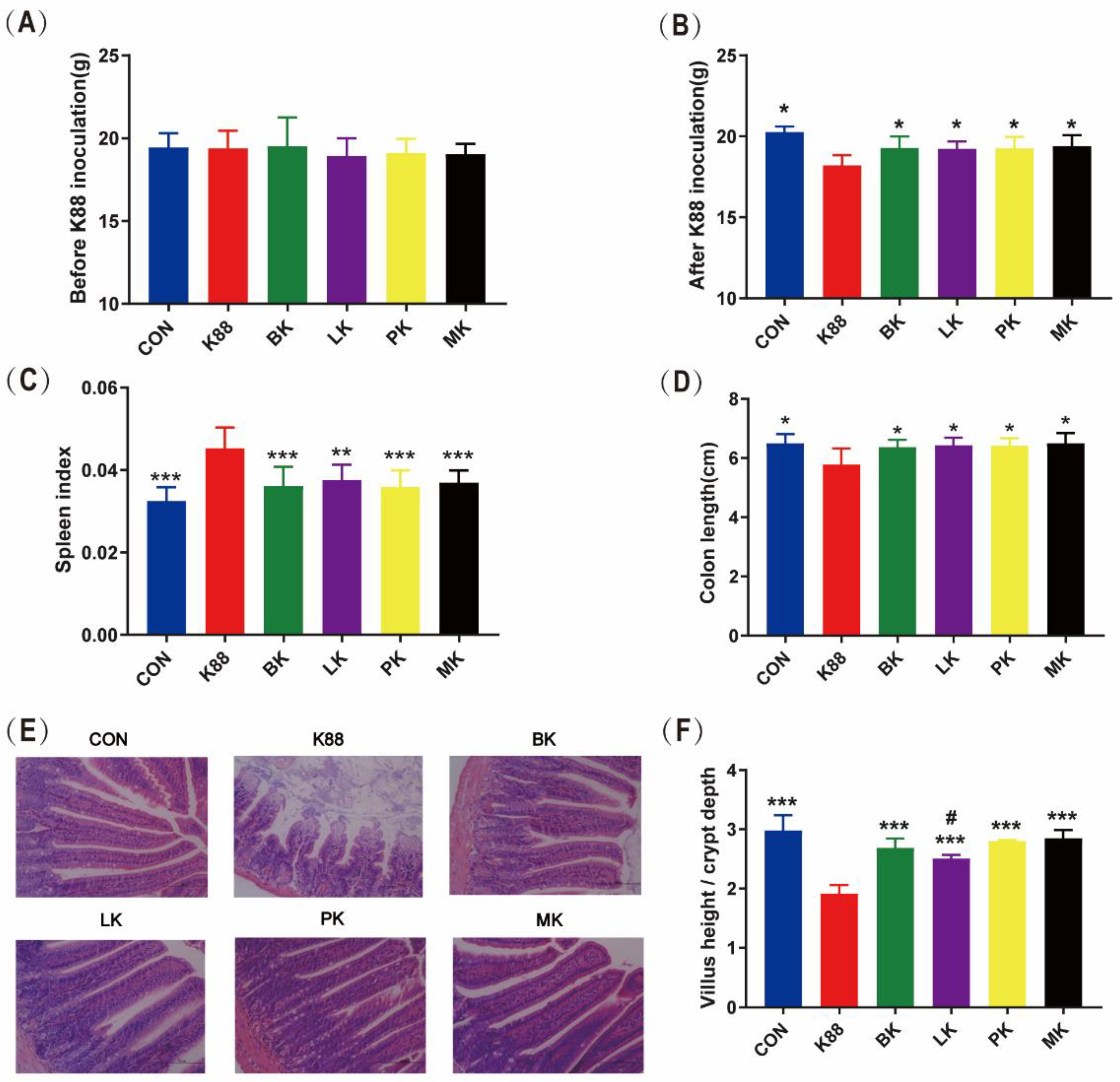
3.1. Effect of Probiotics Administration on Growth Performance of Mice
3.2. Probiotics Recovered the Jejunum Tissue Damage
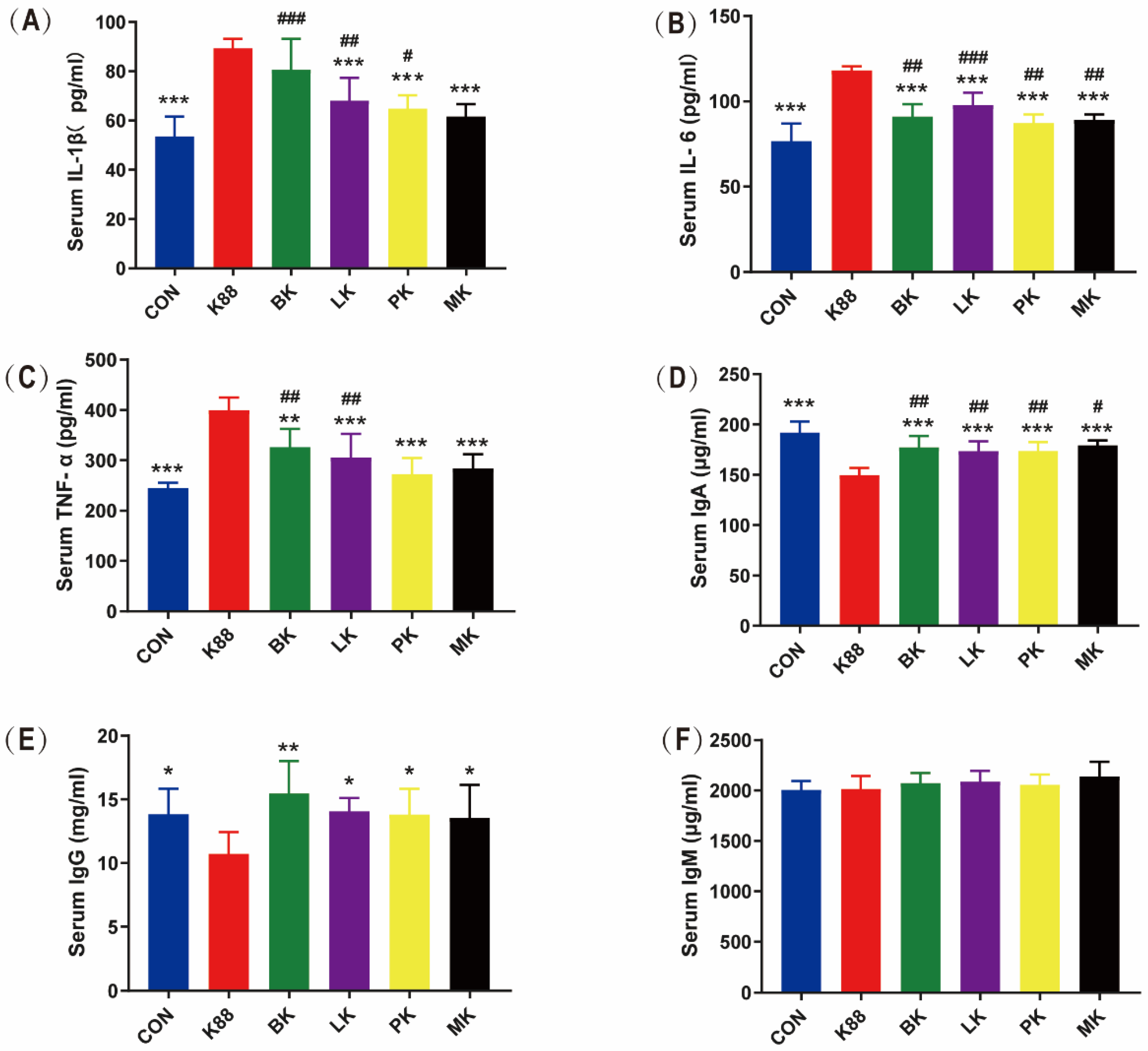
3.3. Effects of Probiotics on Inflammatory Cytokines and Immunoglobulin in Mice Serum
3.4. Effects of Probiotics on Inflammatory Cytokines and Toll-like Receptor mRNA Expression in Jejunum of Mice
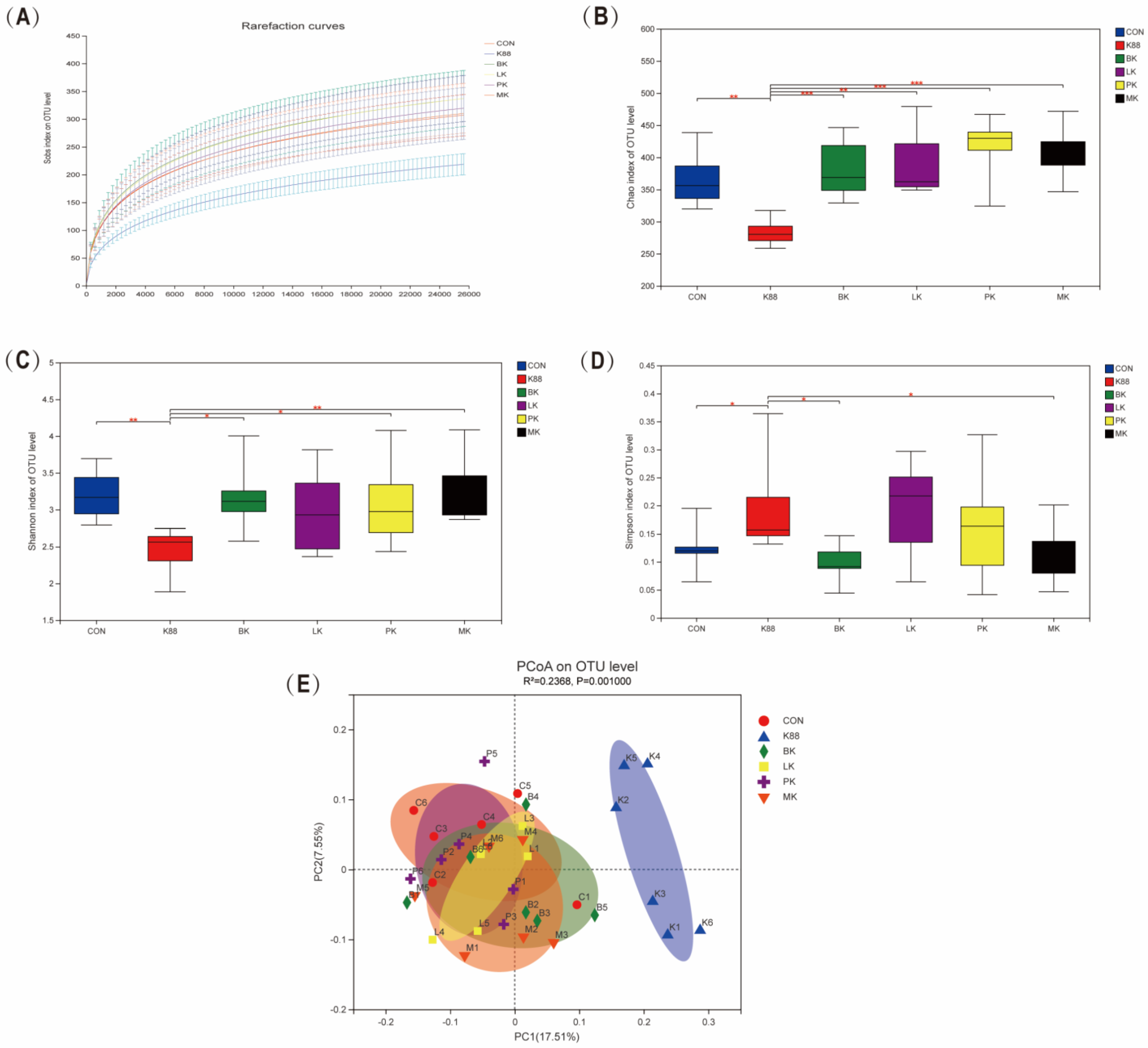
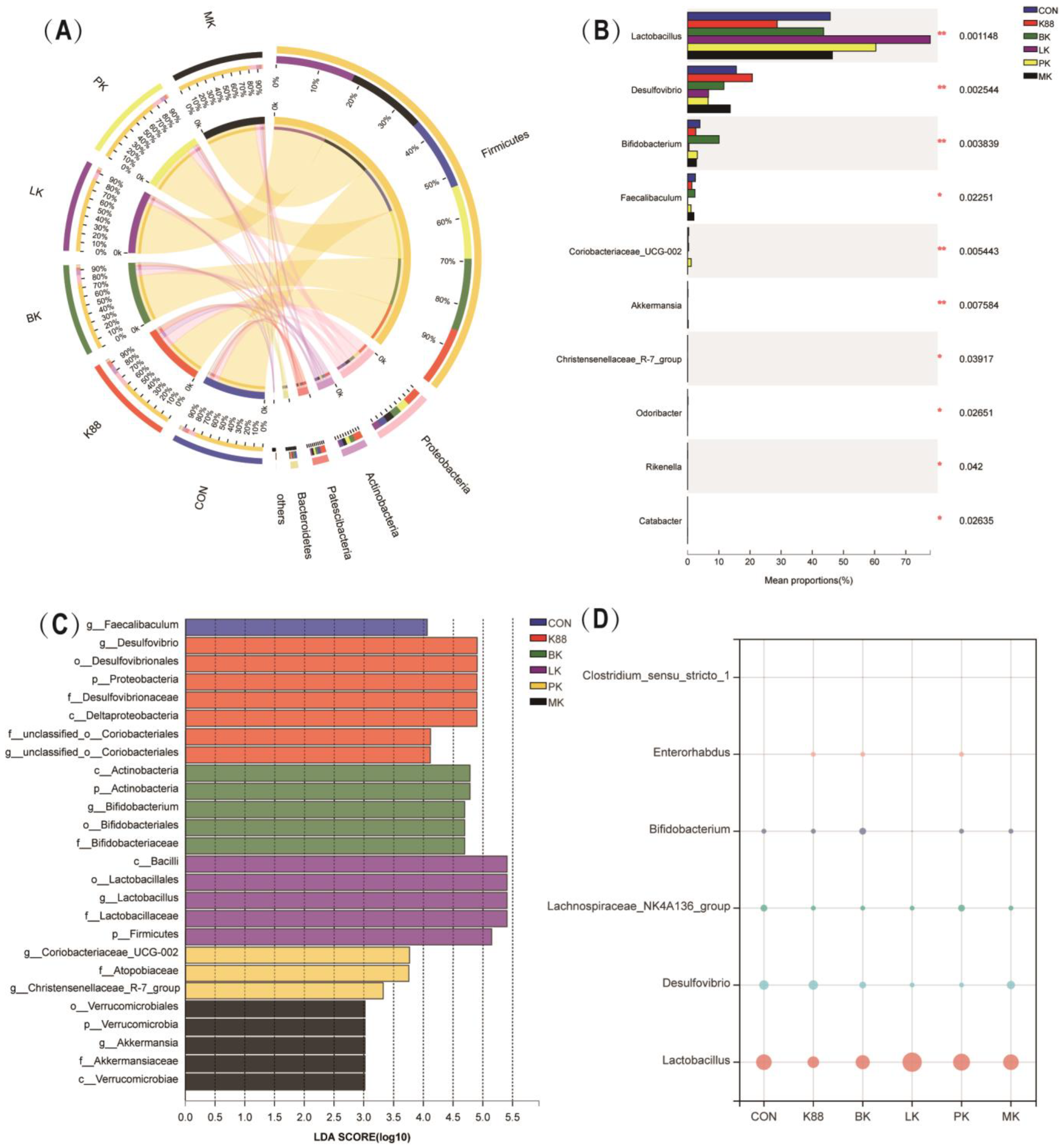
3.5. Probiotics Modulated Intestinal Microbiota Composition Wrecked by K88
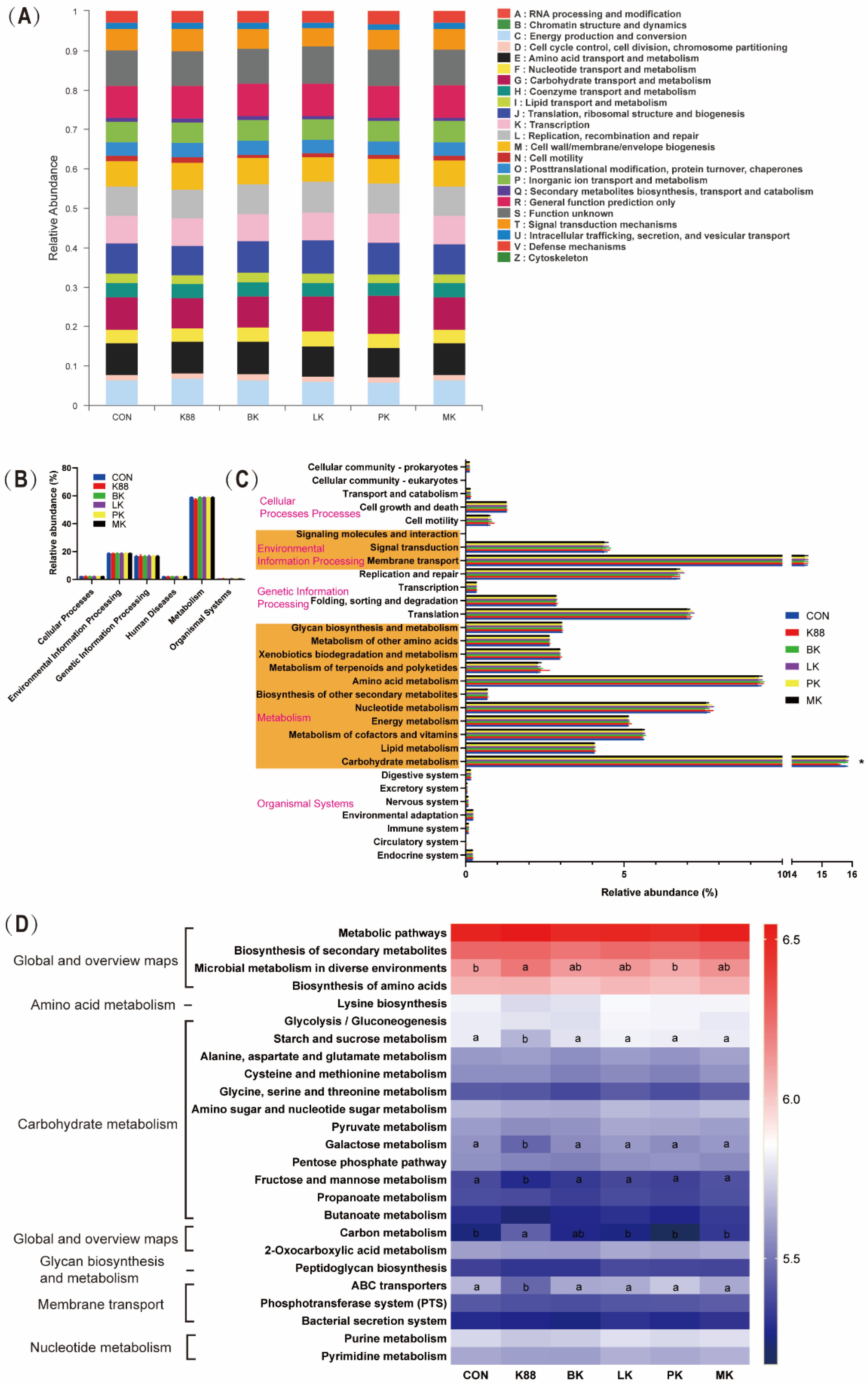
3.6. Bacterial Metabolism of Probiotics Pre-Treatment
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Lozano, R.; Naghavi, M.; Foreman, K.; Lim, S.; Shibuya, K.; Aboyans, V. Global and regional mortality from 235 causes of death for 20 age groups in 1990 and 2010: A systematic analysis for the Global Burden of Disease Study 2010. Lancet 2013, 380, 2095–2128. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.; DuPont, H.L. Etiology of travellers’ diarrhea. J. Travel Med. 2017, 24, S13–S16. [Google Scholar] [CrossRef] [PubMed]
- GBD 2016 Diarrhoeal Disease Collaborators. Estimates of the global, regional, and national morbidity, mortality, and aetiologies of diarrhoea in 195 countries: A systematic analysis for the Global Burden of Disease Study 2016. Lancet Infect Dis. 2018, 18, 1211–1228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gresse, R.; Chaucheyras-Durand, F.; Fleury, M.A.; Wiele, T.V.D.; Forano, E.; Blanquet-Diot, S. Intestinal Microbiota Dysbiosis in Postweaning Piglets: Understanding the Keys to Health. Trends Microbiol. 2017, 25, 851–873. [Google Scholar] [CrossRef]
- Zhu, Y.; Luo, Q.; Davis, S.M.; Westra, C.; Vickers, T.J.; Fleckenstein, J.M. Molecular Determinants of Enterotoxigenic Escherichia coli Heat-Stable Toxin Secretion and Delivery. Infect. Immun. 2018, 86, e00526-e18. [Google Scholar] [CrossRef] [Green Version]
- Alison, L.S.; Timo, V.S.; Tom, J.B.; John, W.M. Effect of weaning age on nursery pig and sow reproductive performance. J. Swine Health Prod. 2008, 16, 131–137. [Google Scholar]
- Sterndale, S.O.; Evans, D.J.; Mansfield, J.P.; Clarke, J.; Sahibzada, S.; Abraham, S.; Miller, D.W.; Kim, J.C.; Pluske, J.R. Effect of mucin 4 allele on susceptibility to experimental infection with enterotoxigenic F4 Escherichia coli in pigs fed experimental diets. J. Anim. Sci. Biotechnol. 2019, 10, 56. [Google Scholar] [CrossRef] [Green Version]
- Zhou, H.; Zhu, J.; Zhu, G. Fimbriae of animal-originated enterotoxigenic Escherichia coli—A review. Acta Microbiol. Sin. 2012, 52, 679–686. [Google Scholar]
- Xia, P.; Song, Y.; Zou, Y.; Yang, Y.; Zhu, G. F4+ enterotoxigenic Escherichia coli (ETEC) adhesion mediated by the major fimbrial subunit FaeG. J. Basic Microbiol. 2015, 55, 1118–1124. [Google Scholar] [CrossRef]
- Broeck, W.V.D.; Cox, E.; Oudega, B.; Goddeeris, B.M. The F4 fimbrial antigen of Escherichia coli and its receptors. Vet. Microbiol. 2000, 71, 223–244. [Google Scholar] [CrossRef]
- Goldenberg, J.Z.; Yap, C.; Lytvyn, L.; Lo, C.K.; Beardsley, J.; Mertz, D.; Johnston, B.C. Probiotics for the prevention of Clostridium difficile-associated diarrhea in adults and children. Cochrane Database Syst. Rev. 2017, 12, CD006095. [Google Scholar] [CrossRef]
- Maltz, C.; Miskovitz, P.F.; Hajifathalian, K. Lactulose may reduce Clostridium difficile-related diarrhea among patients receiving antibiotics. JGH Open 2020, 4, 1088–1090. [Google Scholar] [CrossRef]
- Huang, G.; Li, X.; Lu, D.; Liu, S.; Suo, X.; Li, Q.; Li, N. Lysozyme improves intestinal performance and protects against enterotoxigenic Escherichia coli infection in neonatal piglets. Vet. Res. 2018, 49, 20. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, S.T.; Hwang, J.A.; Hoon, J.; Mun, H.S.; Yang, C.J. Comparison of single and blend acidifiers as alternative to antibiotics on growth performance, fecal microflora, and humoral immunity in weaned piglets. Asian Austral. J. Anim. 2014, 27, 93–100. [Google Scholar] [CrossRef]
- Abiala, M.; Olayiwola, J.; Babatunde, O.; Aiyelaagbe, O.; Akinyemi, S. Evaluation of therapeutic potentials of plant extracts against poultry bacteria threatening public health. BMC Complem. Altern. Med. 2016, 16, 417. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Dou, X.; Song, J.; Lyu, Y.; Zhu, X.; Xu, L.; Li, W.; Shan, A.-S. Antimicrobial peptides: Promising alternatives in the post feeding antibiotic era. Med. Res. Rev. 2019, 39, 831–859. [Google Scholar] [CrossRef]
- Fukuda, S.; Toh, H.; Hase, K.; Oshima, K.; Nakanishi, Y.; Yoshimura, K.; Tobe, T.; Clarke, J.M.; Topping, D.L.; Suzuki, T.; et al. Bifidobacteria can protect from enteropathogenic infection through production of acetate. Nature 2011, 469, 543–547. [Google Scholar] [CrossRef]
- Hrala, M.; Bosák, J.; Micenková, L.; Křenová, J.; Lexa, M.; Pirková, V.; Tomáštíková, Z.; Koláčková, I.; Šmajs, D. Escherichia coli Strains Producing Selected Bacteriocins Inhibit Porcine Enterotoxigenic Escherichia coli (ETEC) under both In Vitro and In Vivo Conditions. Appl. Environ. Microb. 2021, 87, e0312120. [Google Scholar] [CrossRef]
- Bermudez-Brito, M.; Plaza-Díaz, J.; Muñoz-Quezada, S.; Gómez-Llorente, C.; Gil, A. Probiotic mechanisms of action. Ann. Nutr. Metab. 2012, 61, 160–174. [Google Scholar] [CrossRef]
- van Zyl, W.F.; Deane, S.M.; Dicks, L. Molecular insights into probiotic mechanisms of action employed against intestinal pathogenic bacteria. Intest. Microbes 2020, 12, 1831339. [Google Scholar] [CrossRef]
- Ni, X. Application Progress of Autochthonous Microbiota in Broiler Feed. Feed Indus. 2021, 5, 155300. [Google Scholar]
- Duranti, S.; Lugli, G.A.; Milani, C.; James, K.; Mancabelli, L.; Turroni, F.; Alessandri, G.; Mangifesta, M.; Mancino, W.; Ossiprandi, M.C.; et al. Bifidobacterium bifidum and the infant intestinal microbiota: An intriguing case of microbe-host co-evolution. Environ. Microbiol. 2019, 21, 3683–3695. [Google Scholar] [CrossRef]
- O’Callaghan, A.; van Sinderen, D. Bifidobacteria and their role as members of the human intestinal microbiota. Front. Microbiol. 2016, 7, 925. [Google Scholar]
- Wu, T.; Shi, Y.; Zhang, Y.; Zhang, M.; Zhang, L.; Ma, Z.; Zhao, D.; Wang, L.; Yu, H.; Hou, Y.; et al. Lactobacillus rhamnosus LB1 Alleviates Enterotoxigenic Escherichia coli-Induced Adverse Effects in Piglets by Improving Host Immune Response and Anti-Oxidation Stress and Restoring Intestinal Integrity. Front. Cell. Infect. Microbiol. 2021, 11, 724401. [Google Scholar] [CrossRef]
- Yang, B.; Yue, Y.; Chen, Y.; Ding, M.; Li, B.; Wang, L.; Wang, Q.; Stanton, C.; Ross, R.P.; Zhao, J.; et al. Lactobacillus plantarum CCFM1143 Alleviates Chronic Diarrhea via Inflammation Regulation and Intestinal Microbiota Modulation: A Double-Blind, Randomized, Placebo-Controlled Study. Front. Immunol. 2021, 12, 746585. [Google Scholar] [CrossRef] [PubMed]
- Tan, K.; Deng, D.; Ma, X.; Cui, Y.; Tian, Z. Pediococcus acidilactici P25 Protected Caenorhabditis elegans against Enterotoxigenic Escherichia coli K88 Infection and Transcriptomic Analysis of Its Potential Mechanisms. Biomed. Res. Int. 2020, 2020, 7340312. [Google Scholar] [CrossRef] [Green Version]
- Mao, X.; Gu, C.; Hu, H.; Tang, J.; Chen, D.; Yu, B.; He, J.; Yu, J.; Luo, J.; Tian, G. Dietary Lactobacillus rhamnosus GG Supplementation Improves the Mucosal Barrier Function in the Intestine of Weaned Piglets Challenged by Porcine Rotavirus. PLoS ONE 2016, 11, e0146312. [Google Scholar] [CrossRef] [PubMed]
- Xue, C.; Li, Y.; Lv, H.; Zhang, L.; Bi, C.; Dong, N.; Shan, A.; Wang, J. Oleanolic Acid Targets the Intestinal-Liver Axis to Alleviate Metabolic Disorders and Hepatic Steatosis. J. Agric. Food Chem. 2021, 69, 7884–7897. [Google Scholar] [CrossRef]
- Xue, C.; Lv, H.; Li, Y.; Dong, N.; Wang, Y.; Zhou, J.; Shi, B.; Shan, A. Oleanolic acid reshapes the intestinal microbiota and alters immune-related gene expression of intestinal epithelial cells. J. Sci. Food Agric. 2022, 102, 764–773. [Google Scholar] [CrossRef]
- Aluri, J.; Cooper, M.A.; Schuettpelz, L.G. Toll-Like Receptor Signaling in the Establishment and Function of the Immune System. Cells 2021, 10, 1374. [Google Scholar] [CrossRef]
- Damien, P.; Cognasse, F.; Payrastre, B.; Spinelli, S.L.; Blumberg, N.; Arthaud, C.A.; Eyraud, M.A.; Phipps, R.P.; McNicol, A.; Pozzetto, B.; et al. NF-κB Links TLR2 and PAR1 to Soluble Immunomodulator Factor Secretion in Human Platelets. Front. Immunol. 2017, 8, 85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peters, V.B.M.; van de Steeg, E.; van Bilsen, J.; Meijerink, M. Mechanisms and immunomodulatory properties of pre- and probiotics. Benef. Microbes 2019, 10, 225–236. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Ni, X.-Q.; Wang, Q.; Peng, Z.; Niu, L.; Wang, H.; Zhou, Y.; Sun, H.; Pan, K.; Jing, B.; et al. Lactobacillus plantarum BSGP201683 Isolated from Giant Panda Feces Attenuated Inflammation and Improved Intestinal Microflora in Mice Challenged with Enterotoxigenic Escherichia coli. Front. Microbiol. 2017, 8, 1885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bertin, A. Virulence factors of enterotoxigenic E. coli studied in the infant mouse model. Ann. Rech. Vet. 1983, 14, 169–182. [Google Scholar]
- Yu, Q.; Liu, X.; Liu, Y.; Riederer, B.; Li, T.; Tian, D.; Tuo, B.; Shull, G.; Seidler, U. Defective small intestinal anion secretion, dipeptide absorption, and intestinal failure in suckling NBCe1-deficient mice. Pflugers Arch. 2016, 468, 1419–1432. [Google Scholar] [CrossRef]
- Lin, Q.; Fu, Q.; Li, X.; Luo, Y.; Luo, J.; Chen, D.; Mao, X.; Yu, B.; Zheng, P.; Huang, Z.; et al. Human β-Defensin 118 Attenuates Escherichia coli K88-Induced Inflammation and Intestinal Injury in Mice. Probiotics Antimicrob. Proteins 2021, 13, 586–597. [Google Scholar] [CrossRef]
- Han, X.; Ding, S.; Ma, Y.; Fang, J.; Jiang, H.; Li, Y.; Liu, G. Lactobacillus plantarum and Lactobacillus brevis Alleviate Intestinal Inflammation and Microbial Disorder Induced by ETEC in a Murine Model. Oxid. Med. Cell. Longev. 2021, 2021, 6867962. [Google Scholar] [CrossRef]
- Ren, W.; Yin, J.; Duan, J.; Liu, G.; Zhu, X.; Chen, S.; Li, T.; Wang, S.; Tang, Y.; Hardwidge, P.R. Mouse intestinal innate immune responses altered by Enterotoxigenic Escherichia coli (ETEC) infection. Microbes Infect. 2014, 16, 954–961. [Google Scholar] [CrossRef]
- Sanchez-Villamil, J.; Navarro-Garcia, F. Role of virulence factors on host inflammatory response induced by diarrheagenic Escherichia coli pathotypes. Future Microbiol. 2015, 10, 1009–1033. [Google Scholar] [CrossRef]
- Liu, B.; Liu, Q.; Li, G.; Sun, L.; Gao, Y.; Zhang, Y.; Liu, H.; Cao, M.; Liu, G. The anti-diarrhea activity of red algae-originated sulphated polysaccharides on ETEC-K88 infected mice. RSC Adv. 2019, 9, 2360–2370. [Google Scholar] [CrossRef] [Green Version]
- Pupa, P.; Apiwatsiri, P.; Sirichokchatchawan, W.; Pirarat, N.; Nedumpun, T.; Hampson, D.; Muangsin, N.; Prapasarakul, N. Microencapsulated probiotic Lactiplantibacillus plantarum and/or Pediococcus acidilactici strains ameliorate diarrhoea in piglets challenged with Enterotoxigenic Escherichia coli. Sci. Rep. 2022, 12, 7210. [Google Scholar] [CrossRef] [PubMed]
- Megha, K.; Mohanan, P. Role of immunoglobulin and antibodies in disease management. Int. J. Bio. Macromol. 2021, 169, 28–38. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Teng, K.; Liu, G.; Liu, Y.; Zhang, J.; Zhang, X.; Zhang, M.; Tao, Y.; Zhong, J. Lactobacillus reuteri HCM2 protects mice against Enterotoxigenic Escherichia coli through modulation of intestinal microbiota. Sci. Rep. 2018, 8, 17485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fairbrother, J.; Nadeau, E.; Gyles, C. Escherichia coli in postweaning diarrhea in pigs: An update on bacterial types, pathogenesis, and prevention strategies. Anim. Health Res. Rev. 2005, 6, 17–39. [Google Scholar] [CrossRef] [Green Version]
- Yin, H.; Ye, P.; Lei, Q.; Cheng, Y.; Yu, H.; Du, J.; Pan, H.; Cao, Z. In Vitro probiotic properties of Pediococcus pentosaceus L1 and its effects on enterotoxigenic Escherichia coli-induced inflammatory responses in porcine intestinal epithelial cells. Microb. Pathog. 2020, 144, 104163. [Google Scholar] [CrossRef]
- Zeytun, A.; Chaudhary, A.; Pardington, P.; Cary, R.; Gupta, G. Induction of cytokines and chemokines by Toll-like receptor signaling: Strategies for control of inflammation. Crit. Rev. Immunol. 2010, 30, 53–67. [Google Scholar] [CrossRef]
- Zughaier, S.; Zimmer, S.; Datta, A.; Carlson, R.; Stephens, D. Differential induction of the toll-like receptor 4-MyD88-dependent and -independent signaling pathways by endotoxins. Infect Immun. 2005, 73, 2940–2950. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Liu, X.; Shang, Z.; Qiao, J. Clostridium butyricum helps to alleviate inflammation in weaned piglets challenged with Enterotoxigenic Escherichia coli K88. Front. Vet. Sci. 2021, 8, 683863. [Google Scholar] [CrossRef]
- Yang, X.; Xiao, Z.; Liu, F.; Chen, S.; Tang, W.; Zhang, D.; Liu, S. Enterotoxigenic Escherichia coli infection alters intestinal immunity in mice. Mol. Med. Rep. 2016, 14, 825–830. [Google Scholar] [CrossRef] [Green Version]
- Kataoka, K. The intestinal microbiota and its role in human health and disease. J. Med. Investig. 2016, 63, 27–37. [Google Scholar] [CrossRef] [Green Version]
- Yue, Y.; He, Z.; Zhou, Y.; Ross, R.P.; Stanton, C.; Zhao, J.; Zhang, H.; Yang, B.; Chen, W. Lactobacillus plantarum relieves diarrhea caused by enterotoxin-producing Escherichia coli through inflammation modulation and intestinal microbiota regulation. Food Funct. 2020, 11, 10362–10374. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Zhao, M.; Lu, Z.; Lv, F.; Zhao, H.; Bie, X.; Johnsonii, L. L. plantarum, and L. rhamnosus alleviated Enterohaemorrhagic Escherichia coli-induced diarrhoea in mice by regulating intestinal microbiota. Microb. Pathog. 2021, 154, 104856. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Liao, Q.; Lin, M.; Zhong, D.; Wei, L.; Han, B.; Miao, H.; Yao, M.; Xie, Z. An integrated metabonomics and microbiology analysis of host-microbiota metabolic interactions in rats with coptis chinensis-induced diarrhea. RSC Adv. 2015, 5, 79329–79341. [Google Scholar] [CrossRef]
- Sun, X.; Gao, Y.; Wang, X.; Hu, G.; Wang, Y.; Feng, B.; Hu, Y.; Mu, X.; Zhang, Y.; Dong, H. Escherichia coli O101-induced diarrhea develops intestinal microbial dysbiosis in rats. Exp. Ther. Med. 2019, 17, 824–834. [Google Scholar]
- Rizzatti, G.; Lopetuso, L.R.; Gibiino, G.; Binda, C.; Gasbarrini, A. Proteobacteria: A Common Factor in Human Diseases. Biomed. Res. Int. 2017, 2017, 9351507. [Google Scholar] [CrossRef] [Green Version]
- Shin, N.R.; Whon, T.W.; Bae, J.W. Proteobacteria: Microbial signature of dysbiosis in intestinal microbiota. Trends Biotechnol. 2015, 33, 496–503. [Google Scholar] [CrossRef]
- Kushkevych, I.; Dordević, D.; Kollár, P. Analysis of Physiological Parameters of Desulfovibrio Strains from Individuals with Colitis. Open Life Sci. 2019, 13, 481–488. [Google Scholar] [CrossRef]
- Coutinho, C.; Coutinho-Silva, R.; Zinkevich, V.; Pearce, C.B.; Ojcius, D.M.; Beech, I. Sulphate-reducing bacteria from ulcerative colitis patients induce apoptosis of gastrointestinal epithelial cells. Microb. Pathog. 2017, 112, 126–134. [Google Scholar] [CrossRef] [Green Version]
- Guo, W.; Mao, B.; Cui, S.; Tang, X.; Zhang, Q.; Zhao, J.; Zhang, H. Protective effects of a novel probiotic Bifidobacterium pseudolongum on the intestinal barrier of colitis mice via modulating the Pparγ/STAT3 pathway and intestinal microbiota. Foods 2022, 11, 1551. [Google Scholar] [CrossRef]
- Van der Beek, C.M.; Dejong, C.H.C.; Troost, F.J.; Masclee, A.A.M.; Lenaerts, K. Role of short-chain fatty acids in colonic inflammation, carcinogenesis, and mucosal protection and healing. Nutr. Rev. 2017, 75, 286–305. [Google Scholar] [CrossRef] [Green Version]
- Waters, J.L.; Ley, R.E. The human intestinal bacteria Christensenellaceae are widespread, heritable, and associated with health. BMC Biol. 2019, 17, 83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mancabelli, L.; Milani, C.; Lugli, G.A.; Turroni, F.; Cocconi, D.; van Sinderen, D.; Ventura, M. Identification of universal intestinal microbial biomarkers of common human intestinal diseases by meta-analysis. FEMS Microbiol. Ecol. 2017, 93, 153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Depommier, C.; Everard, A.; Druart, C.; Plovier, H.; Van Hul, M.; Vieira-Silva, S.; Falony, G.; Raes, J.; Maiter, D.; Delzenne, N.M.; et al. Supplementation with Akkermansia muciniphila in overweight and obese human volunteers: A proof-of-concept exploratory study. Nat. Med. 2019, 25, 1096–1103. [Google Scholar] [CrossRef] [PubMed]
- Macchione, I.G.; Lopetuso, L.R.; Ianiro, G.; Napoli, M.; Gibiino, G.; Rizzatti, G.; Petito, V.; Gasbarrini, A.; Scaldaferri, F. Akkermansia muciniphila: Key player in metabolic and gastrointestinal disorders. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 8075–8083. [Google Scholar] [PubMed]
- Wang, L.; Tang, L.; Feng, Y.; Zhao, S.; Han, M.; Zhang, C.; Yuan, G.; Zhu, J.; Cao, S.; Wu, Q.; et al. A purified membrane protein from Akkermansia muciniphila or the pasteurised bacterium blunts colitis associated tumourigenesis by modulation of CD8(+) T cells in mice. Intestinal 2020, 69, 1988–1997. [Google Scholar] [CrossRef] [Green Version]
- Frese, S.A.; Hutton, A.A.; Contreras, L.N.; Shaw, C.A.; Palumbo, M.C.; Casaburi, G.; Xu, G.; Davis, J.C.C.; Lebrilla, C.B.; Henrick, B.M.; et al. Persistence of Supplemented Bifidobacterium longum subsp. infantis EVC001 in Breastfed Infants. mSphere 2017, 2, e00501–e00517. [Google Scholar] [CrossRef] [PubMed]
- Ehrlich, A.M.; Pacheco, A.R.; Henrick, B.M.; Taft, D.; Xu, G.; Huda, M.N.; Mishchuk, D.; Goodson, M.L.; Slupsky, C.; Barile, D.; et al. Indole-3-lactic acid associated with Bifidobacterium-dominated microbiota significantly decreases inflammation in intestinal epithelial cells. BMC Microbiol. 2020, 20, 357. [Google Scholar] [CrossRef]
- Jeckelmann, J.M.; Erni, B. Transporters of glucose and other carbohydrates in bacteria. Pflugers Arch. 2020, 472, 1129–1153. [Google Scholar] [CrossRef]
- Smid, E.J.; Lacroix, C. Microbe-microbe interactions in mixed culture food fermentations. Curr. Opin. Biotechnol. 2013, 24, 148–154. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer Sequence (5′ to 3′) | Reverse Primer Sequence (5′ to 3′) | Annealing Temperature (°C) |
|---|---|---|---|
| β-actin | CTAGGCGGACTGTTACTGAGC | CGCCTTCACCGTTCCAGTTT | 60 |
| IL-1β | GCCACCTTTTGACAGTGATGAG | GACAGCCCAGGTCAAAGGTT | 60 |
| IL-6 | GACAAAGCCAGAGTCCTTCAGA | TGTGACTCCAGCTTATCTCTTGG | 60 |
| TNF-α | ATGGCCTCCCTCTCATCAGT | TTTGCTACGACGTGGGCTAC | 60 |
| TLR4 | TTCAGAACTTCAGTGGCTGGATT | CCATGCCTTGTCTTCAATTGTTT | 60 |
| MyD88 | GCATGGTGGTGGTTGTTTCTG | GAATCAGTCGCTTCTGTTGG | 60 |
| NF-κB | CTGAGCGCCCCTCGCATTTA | CCAGCCCATCTTTCTCAGCA | 60 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, W.; Kai, L.; Jiang, Z.; He, H.; Yang, M.; Su, W.; Wang, Y.; Jin, M.; Lu, Z. Bifidobacterium longum, Lactobacillus plantarum and Pediococcus acidilactici Reversed ETEC-Inducing Intestinal Inflammation in Mice. Microorganisms 2022, 10, 2350. https://doi.org/10.3390/microorganisms10122350
Li W, Kai L, Jiang Z, He H, Yang M, Su W, Wang Y, Jin M, Lu Z. Bifidobacterium longum, Lactobacillus plantarum and Pediococcus acidilactici Reversed ETEC-Inducing Intestinal Inflammation in Mice. Microorganisms. 2022; 10(12):2350. https://doi.org/10.3390/microorganisms10122350
Chicago/Turabian StyleLi, Wentao, Lixia Kai, Zipeng Jiang, Huan He, Mingzhi Yang, Weifa Su, Yizhen Wang, Mingliang Jin, and Zeqing Lu. 2022. "Bifidobacterium longum, Lactobacillus plantarum and Pediococcus acidilactici Reversed ETEC-Inducing Intestinal Inflammation in Mice" Microorganisms 10, no. 12: 2350. https://doi.org/10.3390/microorganisms10122350