
Protective Effect of Bifidobacterium animalis subs. lactis MG741 as Probiotics against UVB-Exposed Fibroblasts and Hairless Mice

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Apparatus and Reagents
2.2. Preparation Sample of Bi. lactis MG741
2.3. In Vitro Study
2.3.1. Cytotoxicity
2.3.2. Determination of PIP by EIA
2.4. Animals Study
2.4.1. Experimental Design
2.4.2. Determination of Wrinkle Formation
2.4.3. Histopathological and Skin Hydration Examination
2.5. Quantitative RT-PCR (qRT-PCR)
2.6. Western Blotting
2.7. Statistics
3. Results
3.1. Protective Effect of Bi. lactis MG741 on UVB-Exposed Hs68 Fibroblasts
3.2. Bi. lactis MG741 Exerts Rreduction of mRNA and Protein Expression Related to Wrinkles on UVB-Exposed Hs68 Fibroblasts
3.3. Bi. lactis MG741 Modulates Skin Changes on UVB-Exposed Dorsal Skin of HR-1 Mice
3.4. Effect of Bi. lactis MG741 on Protein Expression Related to Skin Aging on UVB-Exposed Dorsal Skin of HR-1 Mice
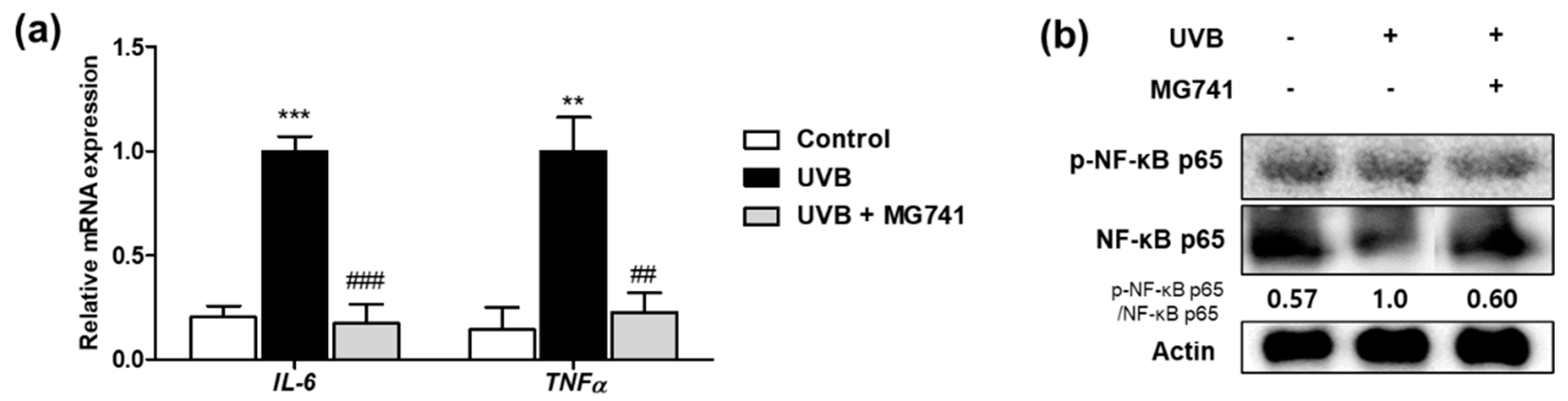
3.5. Bi. lactis MG741 Downregulates Factors Related to Inflammationon on UVB-Exposed Dorsal Skin of HR-1 Mice
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Venus, M.; Waterman, J.; McNab, I. Basic physiology of the skin. Surgery 2010, 28, 469–472. [Google Scholar] [CrossRef]
- Suh, M.G.; Bae, G.Y.; Jo, K.; Kim, J.M.; Hong, K.-B.; Suh, H.J. Photoprotective Effect of Dietary Galacto-Oligosaccharide (GOS) in Hairless Mice via Regulation of the MAPK Signaling Pathway. Molecules 2020, 25, 1679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gromkowska-Kępka, K.J.; Puścion-Jakubik, A.; Markiewicz-Żukowska, R.; Socha, K. The impact of ultraviolet radiation on skin photoaging—review of in vitro studies. J. Cosmet. Dermatol. 2021, 20, 3427–3431. [Google Scholar] [CrossRef]
- Berneburg, M.; Plettenberg, H.; Krutmann, J. Photoaging of human skin. Photodermatol. Photoimmunol. Photomed. Rev. Artic. 2000, 16, 239–244. [Google Scholar] [CrossRef]
- Ansary, T.M.; Hossain, M.R.; Kamiya, K.; Komine, M.; Ohtsuki, M. Inflammatory Molecules Associated with Ultraviolet Radiation-Mediated Skin Aging. Int. J. Mol. Sci. 2021, 22, 3974. [Google Scholar] [CrossRef] [PubMed]
- Kaddurah, H.; Braunberger, T.L.; Vellaichamy, G.; Nahhas, A.F.; Lim, H.W.; Hamzavi, I.H. The Impact of Sunlight on Skin Aging. Curr. Geriatr. Rep. 2018, 7, 228–237. [Google Scholar] [CrossRef]
- Mukherjee, P.K.; Maity, N.; Nema, N.K.; Sarkar, B.K. Bioactive compounds from natural resources against skin aging. Phytomedicine 2011, 19, 64–73. [Google Scholar] [CrossRef] [PubMed]
- Naylor, E.C.; Watson, R.E.; Sherratt, M.J. Molecular aspects of skin ageing. Maturitas 2011, 69, 249–256. [Google Scholar] [CrossRef]
- Shin, J.-W.; Kwon, S.-H.; Choi, J.-Y.; Na, J.-I.; Huh, C.-H.; Choi, H.-R.; Park, K.-C. Molecular Mechanisms of Dermal Aging and Antiaging Approaches. Int. J. Mol. Sci. 2019, 20, 2126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mack, D.R. Probiotics: Mixed messages. Can. Fam. Physician 2005, 51, 1455. [Google Scholar]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. Expert consensus document: The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef] [PubMed]
- Wilkins, T.; Sequoia, J. Probiotics for Gastrointestinal Conditions: A Summary of the Evidence. Am. Fam. Physician 2017, 96, 170–178. [Google Scholar] [PubMed]
- Lolou, V.; Panayiotidis, M.I. Functional Role of Probiotics and Prebiotics on Skin Health and Disease. Fermentation 2019, 5, 41. [Google Scholar] [CrossRef] [Green Version]
- Lew, L.-C.; Liong, M.-T. Bioactives from probiotics for dermal health: Functions and benefits. J. Appl. Microbiol. 2013, 114, 1241–1253. [Google Scholar] [CrossRef] [PubMed]
- Mishra, V.; Sharma, R.; Jaiswal, M.; Subramani, S.K.; Yadav, A.; Rana, S.; Bisen, P.S. A review on recently explored functional aspects of probiotics. Ind. J. Pure Appl. Biosci. 2020, 8, 213–226. [Google Scholar] [CrossRef]
- Wenk, J.; Brenneisen, P.; Meewes, C.; Wlaschek, M.; Peters, T.; Blaudschun, R.; Ma, W.; Kuhr, L.; Schneider, L.; Scharffetter-Kochanek, K. UV-induced oxidative stress and photoaging. Curr. Probl. Dermatol. Basel 2001, 29, 83–94. [Google Scholar]
- Hu, S.; Zhang, X.; Chen, F.; Wang, M. Dietary polyphenols as photoprotective agents against UV radiation. J. Funct. Foods 2017, 30, 108–118. [Google Scholar] [CrossRef]
- Lee, J.Y.; Kang, C.-H. Probiotics alleviate oxidative stress in H2O2-exposed hepatocytes and t-BHP-induced C57BL/6 mice. Microorganisms 2022, 10, 234. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.Y.; Kim, H.; Jeong, Y.; Kang, C.-H. Lactic Acid Bacteria Exert a Hepatoprotective Effect against Ethanol-Induced Liver Injury in HepG2 Cells. Microorganisms 2021, 9, 1844. [Google Scholar] [CrossRef]
- Silveira, J.E.P.S.; Pedroso, D.M.M. UV light and skin aging. Rev. Environ. Health 2014, 29, 243–254. [Google Scholar]
- Hachiya, A.; Sriwiriyanont, P.; Fujimura, T.; Ohuchi, A.; Kitahara, T.; Takema, Y.; Kitzmiller, W.J.; Visscher, M.O.; Tsuboi, R.; Boissy, R.E. Mechanistic Effects of Long-Term Ultraviolet B Irradiation Induce Epidermal and Dermal Changes in Human Skin Xenografts. Am. J. Pathol. 2009, 174, 401–413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ra, J.; Lee, D.E.; Kim, S.H.; Jeong, J.-W.; Ku, H.K.; Kim, T.-Y.; Choi, I.-D.; Jeung, W.; Sim, J.-H.; Ahn, Y.-T. Effect of Oral Administration of Lactobacillus plantarum HY7714 on Epidermal Hydration in Ultraviolet B-Irradiated Hairless Mice. J. Microbiol. Biotechnol. 2014, 24, 1736–1743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.M.; Lee, D.E.; Park, S.D.; Kim, Y.-T.; Kim, Y.J.; Jeong, J.W.; Jang, S.S.; Ahn, Y.-T.; Sim, J.-H.; Huh, C.-S.; et al. Oral Administration of Lactobacillus plantarum HY7714 Protects Hairless Mouse Against Ultraviolet B-Induced Photoaging. J. Microbiol. Biotechnol. 2014, 24, 1583–1591. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.E.; Huh, C.-S.; Ra, J.; Choi, I.-D.; Jeong, J.-W.; Kim, S.-H.; Ryu, J.H.; Seo, Y.K.; Koh, J.S.; Lee, J.-H.; et al. Clinical Evidence of Effects of Lactobacillus plantarum HY7714 on Skin Aging: A Randomized, Double Blind, Placebo-Controlled Study. J. Microbiol. Biotechnol. 2015, 25, 2160–2168. [Google Scholar] [CrossRef]
- Satoh, T.; Murata, M.; Iwabuchi, N.; Odamaki, T.; Wakabayashi, H.; Yamauchi, K.; Abe, F.; Xiao, J. Effect of Bifidobacterium breve B-3 on skin photoaging induced by chronic UV irradiation in mice. Benef. Microbes 2015, 6, 497–504. [Google Scholar] [CrossRef]
- Kim, D.; Lee, K.R.; Kim, N.R.; Park, S.-J.; Lee, M.; Kim, O.-K. Combination of Bifidobacterium longum and Galacto-Oligosaccharide Protects the Skin from Photoaging. J. Med. Food 2021, 24, 606–616. [Google Scholar] [CrossRef]
- Peres, P.; Terra, V.; Guarnier, F.; Cecchini, R.; Cecchini, A. Photoaging and chronological aging profile: Understanding oxidation of the skin. J. Photochem. Photobiol. B Biol. 2011, 103, 93–97. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Hong, Y.; Kim, M. Structural and Functional Changes and Possible Molecular Mechanisms in Aged Skin. Int. J. Mol. Sci. 2021, 22, 12489. [Google Scholar] [CrossRef] [PubMed]
- Jo, K.; Bae, G.Y.; Cho, K.; Park, S.S.; Suh, H.J.; Hong, K.-B. An Anthocyanin-Enriched Extract from Vaccinium uliginosum Improves Signs of Skin Aging in UVB-Induced Photodamage. Antioxidants 2020, 9, 844. [Google Scholar] [CrossRef] [PubMed]
- Nilforoushzadeh, M.A.; Ashtiani, H.R.A.; Jaffary, F.; Jahangiri, F.; Nikkhah, N.; Mahmoudbeyk, M.; Fard, M.; Ansari, Z.; Zare, S. Dermal Fibroblast Cells: Biology and Function in Skin Regeneration. J. Ski. Stem Cell 2017, 4, e69080. [Google Scholar] [CrossRef] [Green Version]
- Kang, M.C.; Yumnam, S.; Kim, S.Y. Oral Intake of Collagen Peptide Attenuates Ultraviolet B Irradiation-Induced Skin Dehydration In Vivo by Regulating Hyaluronic Acid Synthesis. Int. J. Mol. Sci. 2018, 19, 3551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oba, C.; Ohara, H.; Morifuji, M.; Ito, K.; Ichikawa, S.; Kawahata, K.; Koga, J. Collagen hydrolysate intake improves the loss of epidermal barrier function and skin elasticity induced by UVB irradiation in hairless mice. Photodermatol. Photoimmunol. Photomed. 2013, 29, 204–211. [Google Scholar] [CrossRef] [PubMed]
- Edwards, C.; Pearse, A.; Marks, R.; Nishimori, Y.; Matsumoto, K.; Kawai, M. Degenerative Alterations of Dermal Collagen Fiber Bundles in Photodamaged Human Skin and UV-Irradiated Hairless Mouse Skin: Possible Effect on Decreasing Skin Mechanical Properties and Appearance of Wrinkles. J. Investig. Dermatol. 2001, 117, 1458–1463. [Google Scholar] [CrossRef] [PubMed]
- Hideaki, N. Matrix metalloproteinases. J. Biol. Chem. 1999, 274, 21491–21494. [Google Scholar]
- Brenneisen, P.; Sies, H.; Scharffetter-Kochanek, K. Ultraviolet-B irradiation and matrix metalloproteinases: From induction via signaling to initial events. Ann. N. Y. Acad. Sci. 2002, 973, 31–43. [Google Scholar] [CrossRef]
- Pittayapruek, P.; Meephansan, J.; Prapapan, O.; Komine, M.; Ohtsuki, M. Role of Matrix Metalloproteinases in Photoaging and Photocarcinogenesis. Int. J. Mol. Sci. 2016, 17, 868. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiang, H.-M.; Chen, H.-C.; Chiu, H.-H.; Chen, C.-W.; Wang, S.-M.; Wen, K.-C. Neonauclea reticulata(Havil.) Merr Stimulates Skin Regeneration after UVB Exposure via ROS Scavenging and Modulation of the MAPK/MMPs/Collagen Pathway. Evid.-Based Complement. Altern. Med. 2013, 2013, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Capece, D.; Verzella, D.; Flati, I.; Arboretto, P.; Cornice, J.; Franzoso, G. NF-κB: Blending metabolism, immunity, and inflammation. Trends Immunol. 2022, 43, 757–775. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, L.; Wen, X.; Hao, D.; Zhang, N.; He, G.; Jiang, X. NF-κB signaling in skin aging. Mech. Ageing Dev. 2019, 184, 111160. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, J.Y.; Park, J.-Y.; Kim, Y.; Kang, C.-H. Protective Effect of Bifidobacterium animalis subs. lactis MG741 as Probiotics against UVB-Exposed Fibroblasts and Hairless Mice. Microorganisms 2022, 10, 2343. https://doi.org/10.3390/microorganisms10122343
Lee JY, Park J-Y, Kim Y, Kang C-H. Protective Effect of Bifidobacterium animalis subs. lactis MG741 as Probiotics against UVB-Exposed Fibroblasts and Hairless Mice. Microorganisms. 2022; 10(12):2343. https://doi.org/10.3390/microorganisms10122343
Chicago/Turabian StyleLee, Ji Yeon, Jeong-Yong Park, YongGyeong Kim, and Chang-Ho Kang. 2022. "Protective Effect of Bifidobacterium animalis subs. lactis MG741 as Probiotics against UVB-Exposed Fibroblasts and Hairless Mice" Microorganisms 10, no. 12: 2343. https://doi.org/10.3390/microorganisms10122343