

Single-Cell-Based High-Throughput Cultivation and Functional Characterization of Biosurfactant-Producing Bacteria from Soil and Oilfield-Produced Water
 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection and Preparation
2.2. Single-Cell Cultivation and Functional Identification
2.3. Phylogenetic Analysis
2.4. Surface Tension Measurement
2.5. Biosurfactant Characterization (Crude Biosurfactant Extraction and TLC)
2.6. Biosurfactant Production Optimization Using Response Surface Methodology
2.7. Statistical Analysis
3. Results and Discussion
3.1. Workflow of the SCCBB Approach
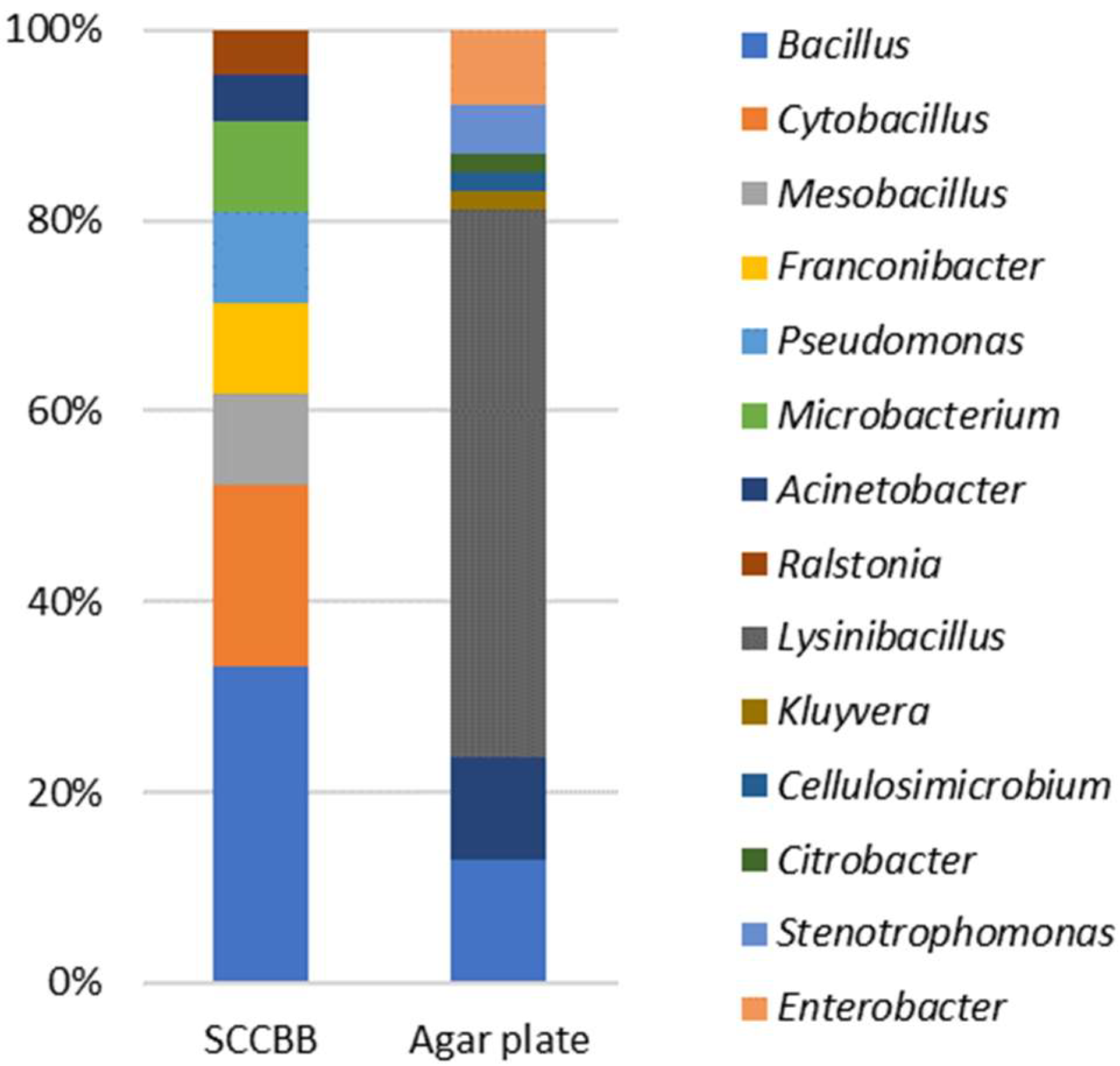
3.2. Single-Cell-Based Inoculation Reduced the Cultivation Bias
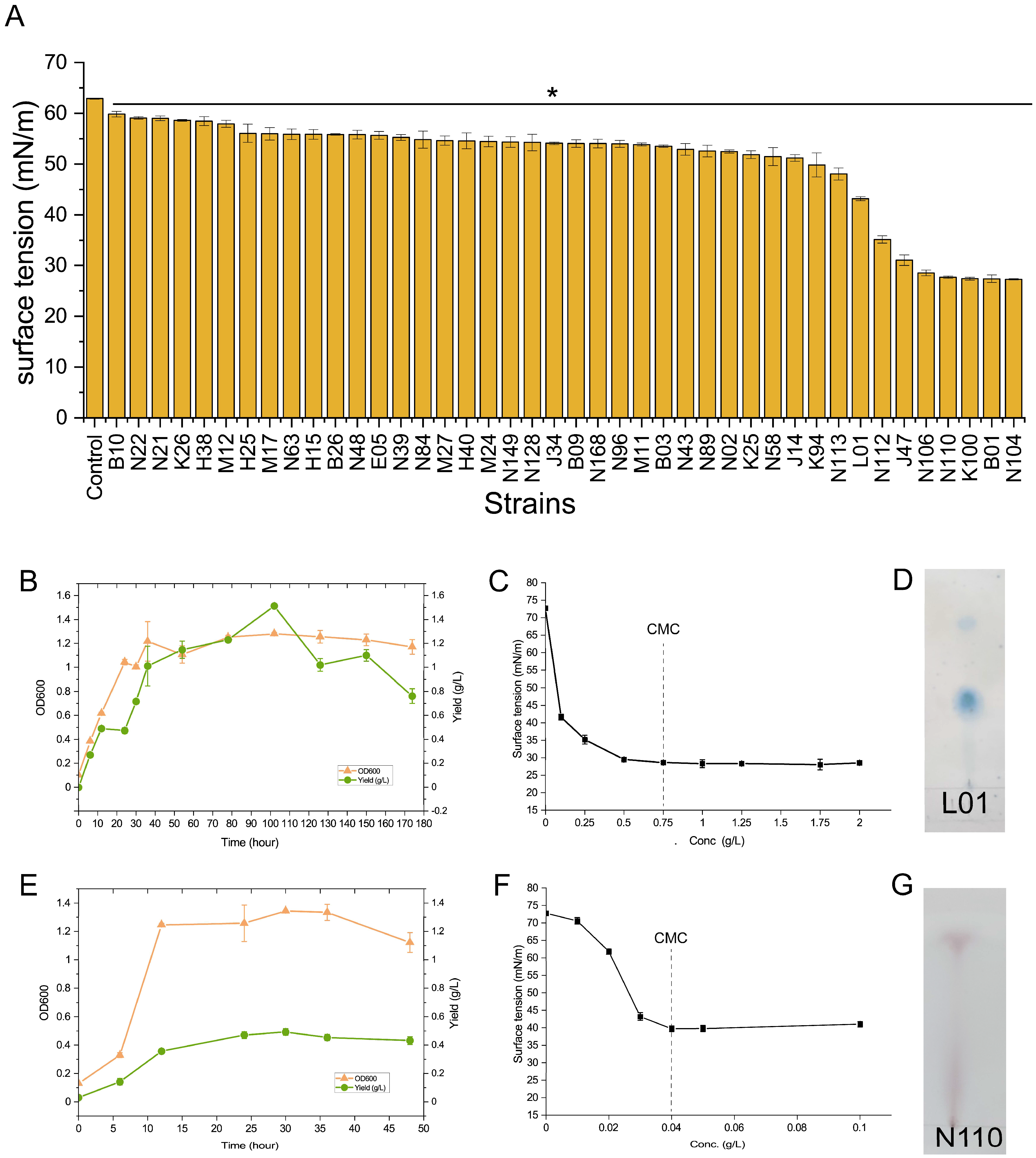
3.3. Isolation of Biosurfactant-Producing Bacterial Strains from Soil and Produced Water Samples
3.4. Functional Verification of the Isolates
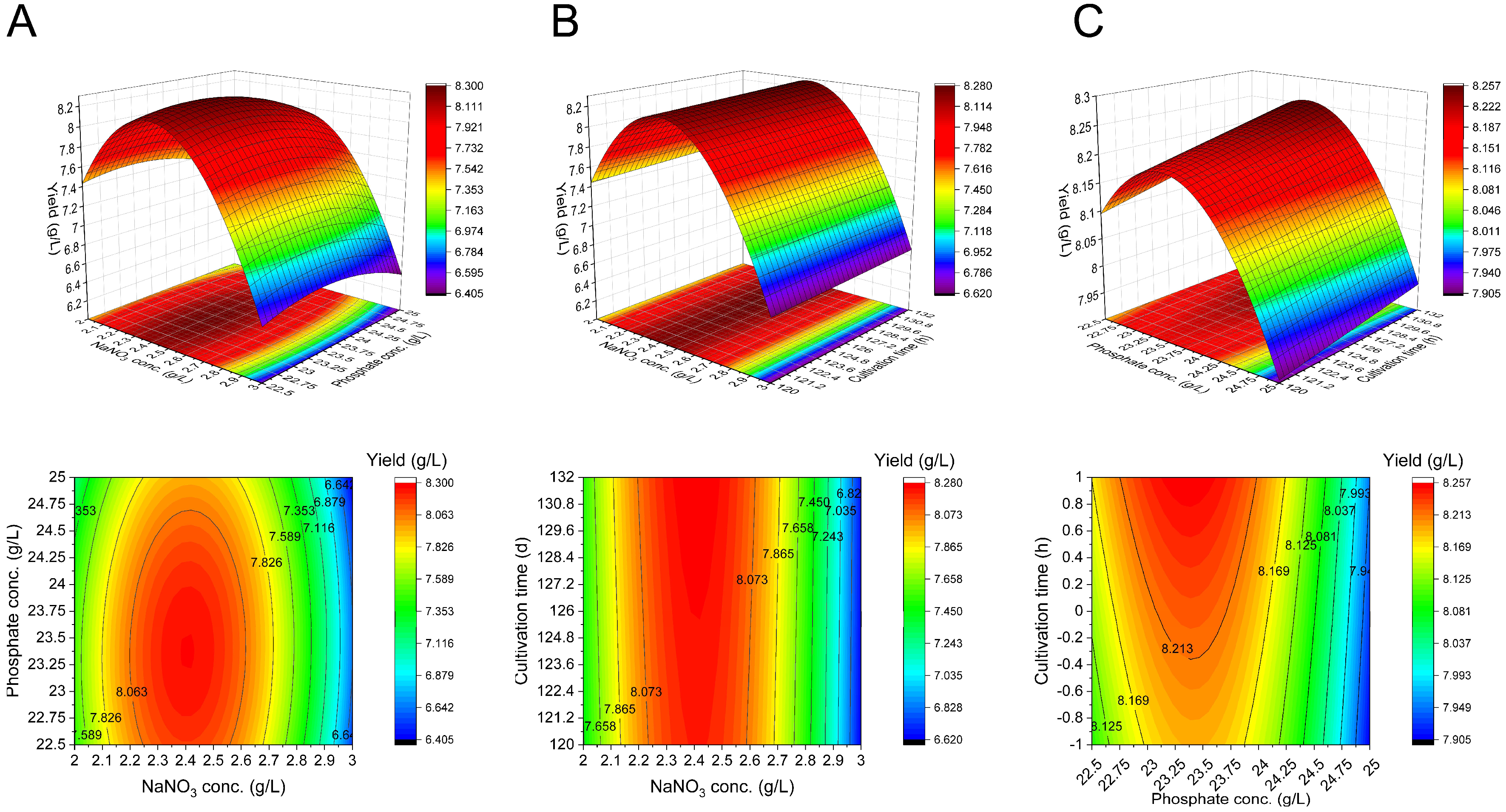
3.5. Optimization of the Glycolipids Yield of Pseudomonas sp. L01 Using Response Surface Methodology (RSM)
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Geys, R.; Soetaert, W.; Van Bogaert, I. Biotechnological opportunities in biosurfactant production. Curr. Opin. Biotechnol. 2014, 30, 66–72. [Google Scholar] [CrossRef] [PubMed]
- Jezierska, S.; Claus, S.; Van Bogaert, I. Yeast glycolipid biosurfactants. FEBS Lett. 2018, 592, 1312–1329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cooper, D.G.; Paddock, D.A. Production of a Biosurfactant from Torulopsis bombicola. Appl. Environ. Microbiol. 1984, 47, 173–176. [Google Scholar] [CrossRef] [Green Version]
- Batista, S.B.; Mounteer, A.H.; Amorim, F.R.; Totola, M.R. Isolation and characterization of biosurfactant/bioemulsifier-producing bacteria from petroleum contaminated sites. Bioresour. Technol. 2006, 97, 868–875. [Google Scholar] [CrossRef] [PubMed]
- Luft, L.; Confortin, T.C.; Todero, I.; Zabot, G.L.; Mazutti, M.A. An overview of fungal biopolymers: Bioemulsifiers and biosurfactants compounds production. Crit. Rev. Biotechnol. 2020, 40, 1059–1080. [Google Scholar] [CrossRef] [PubMed]
- Sachdev, D.P.; Cameotra, S.S. Biosurfactants in agriculture. Appl. Microbiol. Biotechnol. 2013, 97, 1005–1016. [Google Scholar] [CrossRef] [Green Version]
- Nitschke, M.; Costa, S.G.V.A.O. Biosurfactants in food industry. Trends Food Sci. Technol. 2007, 18, 252–259. [Google Scholar] [CrossRef]
- Kumar, A.; Singh, S.K.; Kant, C.; Verma, H.; Kumar, D.; Singh, P.P.; Modi, A.; Droby, S.; Kesawat, M.S.; Alavilli, H.; et al. Microbial Biosurfactant: A New Frontier for Sustainable Agriculture and Pharmaceutical Industries. Antioxidants 2021, 10, 1472. [Google Scholar] [CrossRef]
- Ron, E.Z.; Rosenberg, E. Biosurfactants and oil bioremediation. Curr. Opin. Biotechnol. 2002, 13, 249–252. [Google Scholar] [CrossRef]
- De Almeida, D.G.; Da Silva, R.D.F.S.; Luna, J.M.; Rufino, R.D.; Santos, V.A.; Banat, I.M.; Sarubbo, L.A. Biosurfactants: Promising Molecules for Petroleum Biotechnology Advances. Front. Microbiol. 2016, 7, 1718. [Google Scholar] [CrossRef]
- Rodrigues, L.; Banat, I.M.; Teixeira, J.; Oliveira, R. Biosurfactants: Potential applications in medicine. J. Antimicrob. Chemother. 2006, 57, 609–618. [Google Scholar] [CrossRef] [PubMed]
- Mgbechidinma, C.L.; Akan, O.D.; Zhang, C.; Huang, M.; Linus, N.; Zhu, H.; Wakil, S.M. Integration of Green Economy Concepts for Sustainable Biosurfactant Production—A Review. Bioresour. Technol. 2022, 364, 128021. [Google Scholar] [CrossRef] [PubMed]
- Souza, E.C.; Vessoni-Penna, T.C.; Oliveira, R.P.D. Biosurfactant-enhanced hydrocarbon bioremediation: An overview. Int. Biodeterior. Biodegrad. 2014, 89, 88–94. [Google Scholar] [CrossRef]
- Wang, X.B.; Nie, Y.; Tang, Y.Q.; Wu, G.; Wu, X.L. n-Alkane Chain Length Alters Dietzia sp Strain DQ12-45-1b Biosurfactant Production and Cell Surface Activity. Appl. Environ. Microbiol. 2013, 79, 400–402. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.B.; Chi, C.Q.; Nie, Y.; Tang, Y.Q.; Tan, Y.; Wu, G.; Wu, X.L. Degradation of petroleum hydrocarbons (C6-C40) and crude oil by a novel Dietzia strain. Bioresour. Technol. 2011, 102, 7755–7761. [Google Scholar] [CrossRef]
- Stewart, E.J. Growing Unculturable Bacteria. J. Bacteriol. 2012, 194, 4151–4160. [Google Scholar] [CrossRef] [Green Version]
- Lewis, W.H.; Tahon, G.; Geesink, P.; Sousa, D.Z.; Ettema, T.J.G. Innovations to culturing the uncultured microbial majority. Nat. Rev. Microbiol. 2021, 19, 225–240. [Google Scholar] [CrossRef]
- Kaeberlein, T.; Lewis, K.; Epstein, S.S. Isolating “uncultivable” microorganisms in pure culture in a simulated natural environment. Science 2002, 296, 1127–1129. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.Y.; Liu, Y.X.; Guo, X.X.; Qin, Y.; Garrido-Oter, R.; Schulze-Lefert, P.; Bai, Y. High-throughput cultivation and identification of bacteria from the plant root microbiota. Nat. Protoc. 2021, 16, 988–1012. [Google Scholar] [CrossRef]
- Nichols, D.; Cahoon, N.; Trakhtenberg, E.M.; Pham, L.; Mehta, A.; Belanger, A.; Kanigan, T.; Lewis, K.; Epstein, S.S. Use of Ichip for High-Throughput In Situ Cultivation of “Uncultivable” Microbial Species. Appl. Environ. Microbiol. 2010, 76, 2445–2450. [Google Scholar] [CrossRef]
- Youssef, N.H.; Duncan, K.E.; Nagle, D.P.; Savage, K.N.; Knapp, R.M.; McInerney, M.J. Comparison of methods to detect biosurfactant production by diverse microorganisms. J. Microbiol. Methods 2004, 56, 339–347. [Google Scholar] [CrossRef] [PubMed]
- Jain, D.K.; Collinsthompson, D.L.; Lee, H.; Trevors, J.T. A Drop-Collapsing Test for Screening Surfactant-Producing Microorganisms. J. Microbiol. Methods 1991, 13, 271–279. [Google Scholar] [CrossRef]
- Bodour, A.A.; Miller-Maier, R.M. Application of a modified drop-collapse technique for surfactant quantitation and screening of biosurfactant-producing microorganisms. J. Microbiol. Methods 1998, 32, 273–280. [Google Scholar] [CrossRef]
- Chen, C.Y.; Baker, S.C.; Darton, R.C. The application of a high throughput analysis method for the screening of potential biosurfactants from natural sources. J. Microbiol. Methods 2007, 70, 503–510. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.W.; Wang, M.X.; Nie, Y.; Wu, X.L. Isolation Chip Increases Culturable Bacterial Diversity and Reduces Cultivation Bias. Curr. Microbiol. 2021, 78, 2025–2032. [Google Scholar] [CrossRef] [PubMed]
- Stecher, G.; Tamura, K.; Kumar, S.; Russo, C. Molecular Evolutionary Genetics Analysis (MEGA) for macOS. Mol. Biol. Evol. 2020, 37, 1237–1239. [Google Scholar] [CrossRef]
- Saitou, N.; Nei, M. The Neighbor-Joining Method—A New Method for Reconstructing Phylogenetic Trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [CrossRef]
- Tamura, K.; Nei, M.; Kumar, S. Prospects for inferring very large phylogenies by using the neighbor-joining method. Proc. Natl. Acad. Sci. USA 2004, 101, 11030–11035. [Google Scholar] [CrossRef] [Green Version]
- Zhou, G.; Qiu, H.; Zhang, Q.; Xu, M.; Wang, J.; Wang, G. Experimental Investigation of Coal Dust Wettability Based on Surface Contact Angle. J. Chem. 2016, 2016, 9452303. [Google Scholar] [CrossRef] [Green Version]
- Gallegos-Monterrosa, R.; Kankel, S.; Gotze, S.; Barnett, R.; Stallforth, P.; Kovacs, A.T. Lysinibacillus fusiformis M5 Induces Increased Complexity in Bacillus subtilis 168 Colony Biofilms via Hypoxanthine. J. Bacteriol. 2017, 199, e00204-17. [Google Scholar] [CrossRef]
- Karmakar, K.; Bhattacharya, R.; Sharma, A.; Parmar, K.; Nath, U.; Nataraja, K.N.; Earanna, N.; Sharma, G.; Chakravortty, D. Lysinibacillus macroides-mediated control of cellulose-producing morphotype of Salmonella. J. Sci. Food Agric. 2022, 102, 6491–6501. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, I.; Yokota, A.; Yamazoe, A.; Fujiwara, T. Proposal of Lysinibacillus boronitolerans gen. nov sp nov., and transfer of Bacillus fusiformis to Lysinibacillus fusiformis comb. nov and Bacillus sphaericus to Lysinibacillus sphaericus comb. nov. Int. J. Syst. Evol. Microbiol. 2007, 57, 1117–1125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, M.; Oh, H.S.; Park, S.C.; Chun, J. Towards a taxonomic coherence between average nucleotide identity and 16S rRNA gene sequence similarity for species demarcation of prokaryotes. Int. J. Syst. Evol. Microbiol. 2014, 64, 346–351. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.-Y.; Hu, B.; Dolfing, J.; Li, Y.; Tang, Y.-Q.; Jiang, Y.; Chi, C.-Q.; Xing, J.; Nie, Y.; Wu, X.-L. Thermodynamically favorable reactions shape the archaeal community affecting bacterial community assembly in oil reservoirs. Sci. Total Environ. 2021, 781, 146506. [Google Scholar] [CrossRef]
- Ghojavand, H.; Vahabzadeh, F.; Roayaei, E.; Shahraki, A.K. Production and properties of a biosurfactant obtained from a member of the Bacillus subtilis group (PTCC 1696). J. Colloid Interface Sci. 2008, 324, 172–176. [Google Scholar] [CrossRef]
- Desai, J.D.; Banat, I.M. Microbial production of surfactants and their commercial potential. Microbiol. Mol. Biol. Rev. 1997, 61, 47–64. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | Low Level (−1) | High Level (+1) | Effect | T-Value | p-Value |
|---|---|---|---|---|---|
| glucose (g/L) | 20 | 30 | 0.1120 | 1.19 | 0.281 |
| NaNO3 (g/L) | 1 | 3 | 1.0537 | 11.15 | 0.000 |
| PO43− (g/L) | 20 | 25 | 0.1637 | 1.73 | 0.134 |
| pH | 6.8 | 7.2 | −0.1480 | −1.57 | 0.168 |
| time (h) | 108 | 132 | 0.2520 | 2.67 | 0.037 |
| Trials | NaNO3 (g/L) | PO43− (g/L) | Time (h) | Yields (g/L) |
|---|---|---|---|---|
| 1 | 1.0 | 20 | 108 | 3.55 |
| 2 | 1.5 | 21.25 | 114 | 6.09 |
| 3 | 2.0 | 22.5 | 120 | 6.97 |
| 4 | 2.5 | 23.75 | 126 | 8.23 |
| 5 | 3.0 | 25 | 132 | 6.06 |
| Source | Sum of Squares | df | Mean Square | F-Value | p-Value |
|---|---|---|---|---|---|
| Model | 4.58 | 8 | 0.5725 | 114.11 | 0.0012 * |
| B (NaNO3) | 1.39 | 1 | 1.39 | 277.96 | 0.0005 * |
| C (PO43−) | 0.0841 | 1 | 0.0841 | 16.75 | 0.0264 * |
| E (time) | 0.0072 | 1 | 0.0072 | 1.44 | 0.3169 |
| BC | 0.0042 | 1 | 0.0042 | 0.8422 | 0.4264 |
| BE | 0.0002 | 1 | 0.0002 | 0.0449 | 0.8459 |
| CE | 0.0002 | 1 | 0.0002 | 0.0449 | 0.8459 |
| B2 | 2.67 | 1 | 2.67 | 531.84 | 0.0002 * |
| C2 | 0.0648 | 1 | 0.0648 | 12.92 | 0.0369 * |
| E2 | 0.0000 | 0 | |||
| Residual | 0.0150 | 3 | 0.0050 | ||
| Cor Total | 4.59 | 11 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, Y.; Jing, Y.; Zhang, Y.; Liu, Q.; Xiu, J.; Zhang, K.; Jia, N.; Zhou, M.; Zhou, X.; Huang, J.; et al. Single-Cell-Based High-Throughput Cultivation and Functional Characterization of Biosurfactant-Producing Bacteria from Soil and Oilfield-Produced Water. Microorganisms 2022, 10, 2216. https://doi.org/10.3390/microorganisms10112216
Xu Y, Jing Y, Zhang Y, Liu Q, Xiu J, Zhang K, Jia N, Zhou M, Zhou X, Huang J, et al. Single-Cell-Based High-Throughput Cultivation and Functional Characterization of Biosurfactant-Producing Bacteria from Soil and Oilfield-Produced Water. Microorganisms. 2022; 10(11):2216. https://doi.org/10.3390/microorganisms10112216
Chicago/Turabian StyleXu, Ying, Yali Jing, Yaqian Zhang, Qingjie Liu, Jianlong Xiu, Ke Zhang, Ninghong Jia, Minghui Zhou, Xinyu Zhou, Jia Huang, and et al. 2022. "Single-Cell-Based High-Throughput Cultivation and Functional Characterization of Biosurfactant-Producing Bacteria from Soil and Oilfield-Produced Water" Microorganisms 10, no. 11: 2216. https://doi.org/10.3390/microorganisms10112216