

Oral Administration of Recombinant Lactoferrin-Expressing Probiotics Ameliorates Diet-Induced Lipid Accumulation and Inflammation in Non-Alcoholic Fatty Liver Disease in Mice

Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of Probiotics and Paraprobiotics
2.2. Animals and High-Fat Diet-Induced Obesity/NAFLD Model
2.3. Probiotic Treatment
2.4. Determination of Body Weight and Food Intake, and Biochemical Analyses
2.5. Histologic Analysis and Severity Scoring
2.6. Statistical Analysis
3. Results
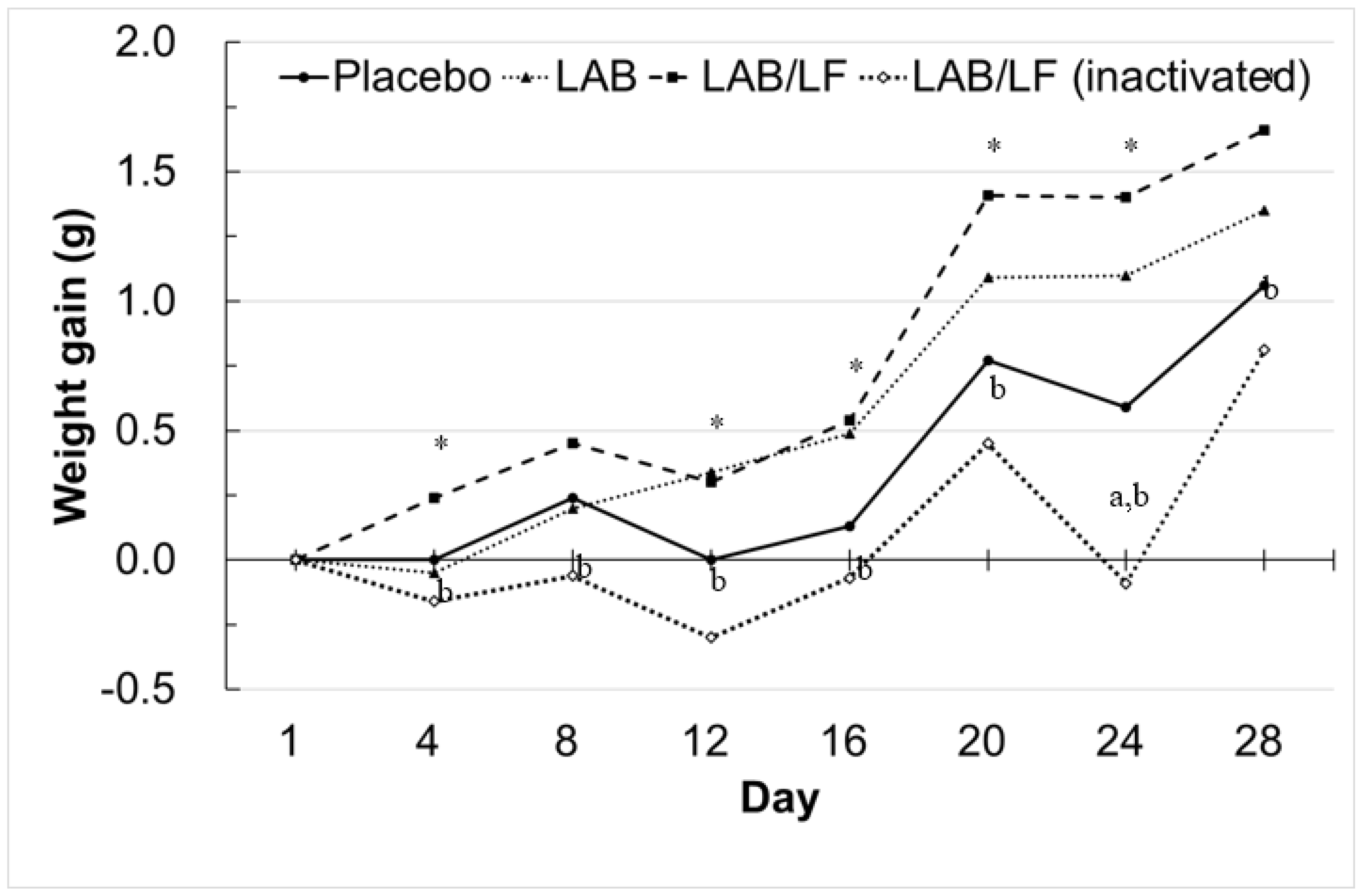
3.1. Effects of 4 Weeks of Administering a Probiotic Mixture on the Body Parameters and Organ Weights of Obese Mice
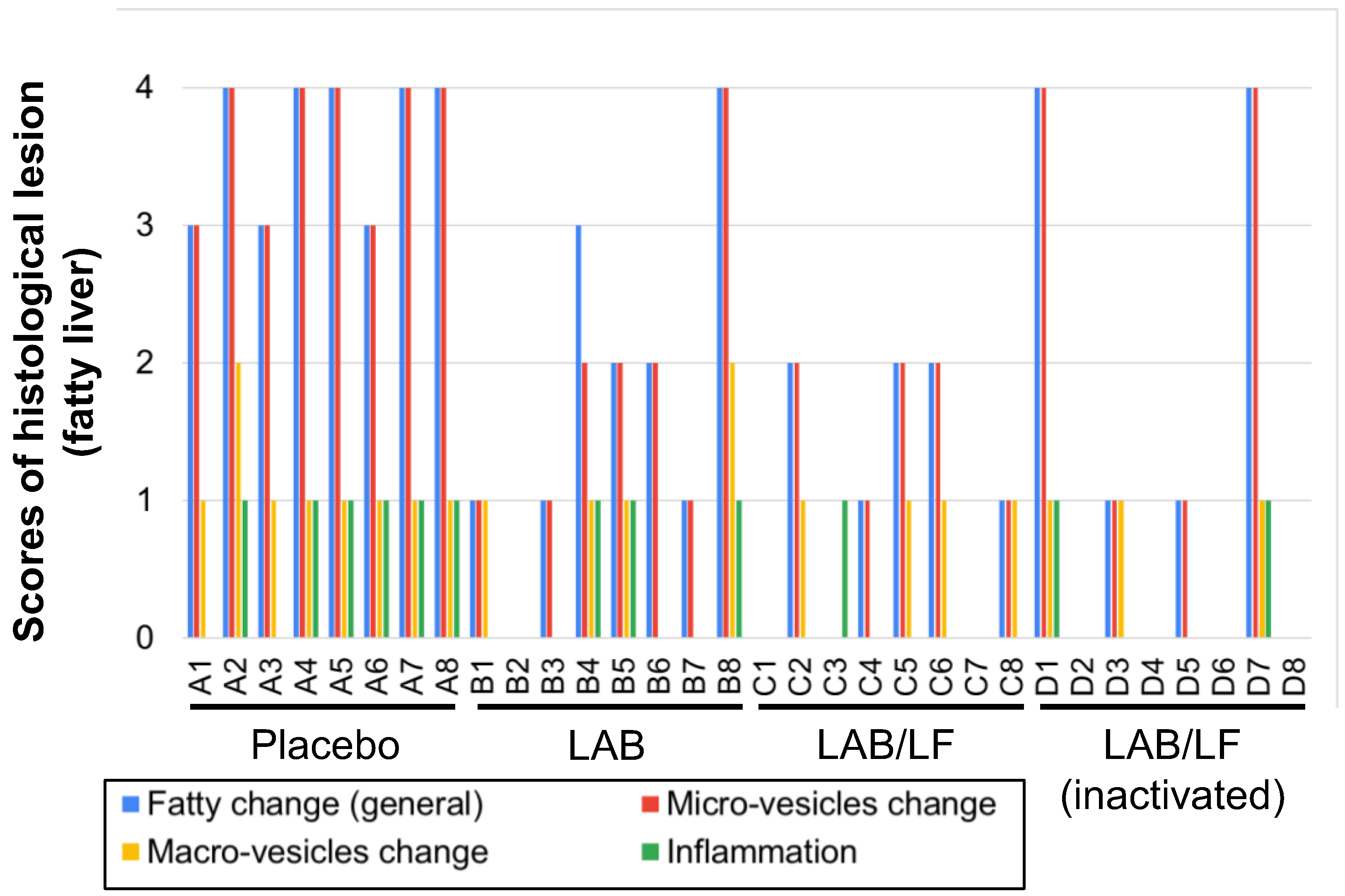
3.2. Effects of the Administration of a Probiotic Mixture on Fatty Accumulation and Inflammation in the Tissues of DIO Mice
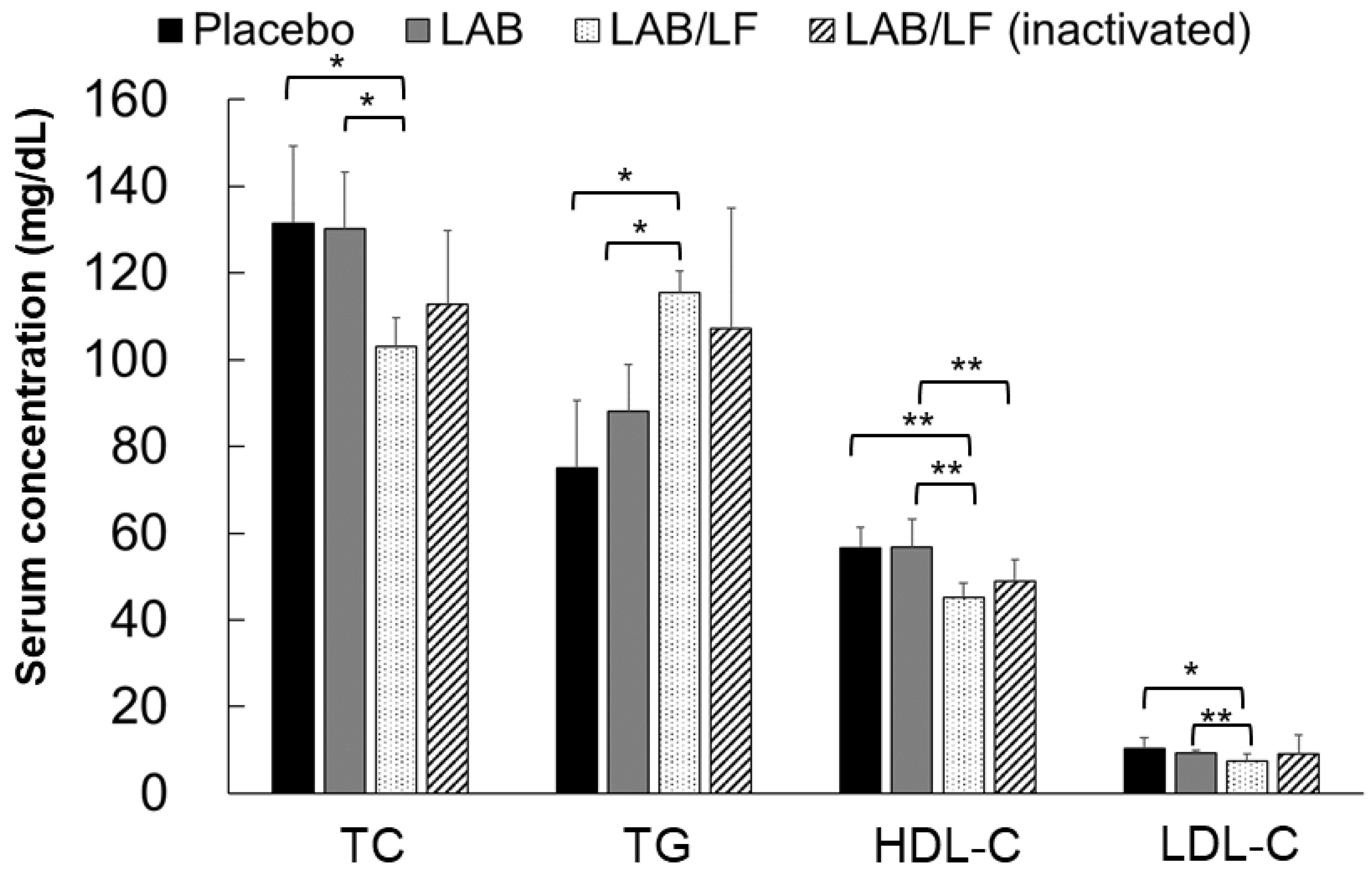
3.3. Effects of the Administration of Probiotics on the Serum Lipid Profiles of Obese Mice
3.4. Effects of Administration of Probiotics on the Biochemical Parameters of Obese Mice
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ludwig, J.; Viggiano, T.R.; McGill, D.B.; Oh, B.J. Nonalcoholic steatohepatitis: Mayo Clinic experiences with a hitherto unnamed disease. Mayo Clin. Proc. 1980, 55, 434–438. [Google Scholar]
- Powell, E.E.; Wong, V.W.-S.; Rinella, M.J.T.L. Non-alcoholic fatty liver disease. Lancet 2021, 397, 2212–2224. [Google Scholar] [CrossRef]
- Schwenger, K.; Allard, J.P. Clinical approaches to non-alcoholic fatty liver disease. World J. Gastroenterol. 2014, 20, 1712–1723. [Google Scholar] [CrossRef]
- Farrell, G.C.; Wong, V.W.-S.; Chitturi, S. NAFLD in Asia—As common and important as in the West. Nat. Rev. Gastroenterol. Hepatol. 2013, 10, 307–318. [Google Scholar] [CrossRef]
- Marchesini, G.; Bugianesi, E.; Forlani, G.; Cerrelli, F.; Lenzi, M.; Manini, R.; Natale, S.; Vanni, E.; Villanova, N.; Melchionda, N.; et al. Nonalcoholic fatty liver, steatohepatitis, and the metabolic syndrome. Hepatology 2003, 37, 917–923. [Google Scholar] [CrossRef]
- Machado, M.; Marques-Vidal, P.; Cortez-Pinto, H. Hepatic histology in obese patients undergoing bariatric surgery. J. Hepatol. 2006, 45, 600–606. [Google Scholar] [CrossRef]
- Yki-Järvinen, H. Non-alcoholic fatty liver disease as a cause and a consequence of metabolic syndrome. Lancet Diabetes Endocrinol. 2014, 2, 901–910. [Google Scholar] [CrossRef]
- Leoni, S.; Tovoli, F.; Napoli, L.; Serio, I.; Ferri, S.; Bolondi, L. Current guidelines for the management of non-alcoholic fatty liver disease: A systematic review with comparative analysis. World J. Gastroenterol. 2018, 24, 3361–3373. [Google Scholar] [CrossRef]
- Paolella, G.; Mandato, C.; Pierri, L.; Poeta, M.; Di Stasi, M.; Vajro, P. Gut-liver axis and probiotics: Their role in non-alcoholic fatty liver disease. World J. Gastroenterol. 2014, 20, 15518–15531. [Google Scholar] [CrossRef]
- Xiao, M.-W.; Lin, S.-X.; Shen, Z.-H.; Luo, W.-W.; Wang, X.-Y. Systematic Review with Meta-Analysis: The Effects of Probiotics in Nonalcoholic Fatty Liver Disease. Gastroenterol. Res. Pract. 2019, 2019, 1484598. [Google Scholar] [CrossRef]
- Koopman, N.; Molinaro, A.; Nieuwdorp, M.; Holleboom, A.G. Review article: Can bugs be drugs? The potential of probiotics and prebiotics as treatment for non-alcoholic fatty liver disease. Aliment. Pharmacol. Ther. 2019, 50, 628–639. [Google Scholar] [CrossRef]
- de Almada, C.N.; Almada, C.N.; Martinez, R.C.; Sant’Ana, A.S. Paraprobiotics: Evidences on their ability to modify biological responses, inactivation methods and perspectives on their application in foods. Trends Food Sci. Technol. 2016, 58, 96–114. [Google Scholar] [CrossRef]
- Aguilar-Toalá, J.E.; Garcia-Varela, R.; Garcia, H.S.; Mata-Haro, V.; González-Córdova, A.F.; Vallejo-Cordoba, B.; Hernández-Mendoza, A. Postbiotics: An evolving term within the functional foods field. Trends Food Sci. Technol. 2018, 75, 105–114. [Google Scholar] [CrossRef]
- Wang, Y.-C.; Hu, S.-Y.; Chiu, C.-S.; Liu, C.-H. Multiple-strain probiotics appear to be more effective in improving the growth performance and health status of white shrimp, Litopenaeus vannamei, than single probiotic strains. Fish Shellfish Immunol. 2018, 84, 1050–1058. [Google Scholar] [CrossRef]
- Actor, J.K.; Hwang, S.-A.; Kruzel, M.L. Lactoferrin as a natural immune modulator. Curr. Pharm. Des. 2009, 15, 1956–1973. [Google Scholar] [CrossRef] [Green Version]
- Kang, X.; Liang, H.; Luo, Y.; Li, Z.; He, F.; Han, X. Anti-adipogenesis and metabolism-regulating effects of heat-inactivated Streptococcus thermophilus MN-ZLW-002. Lett. Appl. Microbiol. 2021, 72, 677–687. [Google Scholar] [CrossRef]
- Tanaka, Y.; Hirose, Y.; Yamamoto, Y.; Yoshikai, Y.; Murosaki, S. Daily intake of heat-killed Lactobacillus plantarum L-137 improves inflammation and lipid metabolism in overweight healthy adults: A randomized-controlled trial. Randomized Control. Trial 2019, 59, 2641–2649. [Google Scholar] [CrossRef] [Green Version]
- Lim, J.-J.; Jung, A.-H.; Suh, H.J.; Choi, H.-S.; Kim, H. Lactiplantibacillus plantarum K8-based paraprobiotics prevents obesity and obesity-induced inflammatory responses in high fat diet-fed mice. Food Res. Int. 2022, 155, 111066. [Google Scholar] [CrossRef]
- Nataraj, B.H.; Ali, S.A.; Behare, P.V.; Yadav, H. Postbiotics-parabiotics: The new horizons in microbial biotherapy and functional foods. Microb. Cell Factories 2020, 19, 1–22. [Google Scholar] [CrossRef]
- Frioni, A.; Conte, M.P.; Cutone, A.; Longhi, C.; Musci, G.; Di Patti, M.C.B.; Natalizi, T.; Marazzato, M.; Lepanto, M.S.; Puddu, P.; et al. Lactoferrin differently modulates the inflammatory response in epithelial models mimicking human inflammatory and infectious diseases. BioMetals 2014, 27, 843–856. [Google Scholar] [CrossRef]
- Legrand, D. Overview of Lactoferrin as a Natural Immune Modulator. J. Pediatr. 2016, 173, S10–S15. [Google Scholar] [CrossRef]
- Valenti, P.; Antonini, G. Lactoferrin: An important host defence against microbial and viral attack. Cell Mol. Life Sci. 2005, 62, 2576–2587. [Google Scholar] [CrossRef] [PubMed]
- Mayeur, S.; Spahis, S.; Pouliot, Y.; Levy, E. Lactoferrin, a Pleiotropic Protein in Health and Disease. Antioxid. Redox Signal. 2016, 24, 813–836. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brimelow, R.; West, N.P.; Williams, L.; Cripps, A.W.; Cox, A.J. A role for whey-derived lactoferrin and immunoglobulins in the attenuation of obesity-related inflammation and disease. Crit. Rev. Food Sci. Nutr. 2015, 57, 1593–1602. [Google Scholar] [CrossRef] [PubMed]
- Artym, J. A remedy against obesity? The role of lactoferrin in the metabolism of glucose and lipids. Postepy Hig. Med. Dosw. 2012, 66, 937–953. [Google Scholar] [CrossRef]
- Sun, J.; Ren, F.; Xiong, L.; Zhao, L.; Guo, H. Bovine lactoferrin suppresses high-fat diet induced obesity and modulates gut microbiota in C57BL/6J mice. J. Funct. Foods 2016, 22, 189–200. [Google Scholar] [CrossRef]
- Li, Y.-C.; Hsieh, C.-C. Lactoferrin dampens high-fructose corn syrup-induced hepatic manifestations of the metabolic syndrome in a murine model. PLoS ONE 2014, 9, e97341. [Google Scholar] [CrossRef] [Green Version]
- Pilvi, T.K.; Harala, S.; Korpela, R.; Mervaala, E.M. Effects of high-calcium diets with different whey proteins on weight loss and weight regain in high-fat-fed C57BL/6J mice. Br. J. Nutr. 2009, 102, 337–341. [Google Scholar] [CrossRef]
- Shi, J.; Finckenberg, P.; Martonen, E.; Ahlroos-Lehmus, A.; Pilvi, T.K.; Korpela, R.; Mervaala, E.M. Metabolic effects of lactoferrin during energy restriction and weight regain in diet-induced obese mice. J. Funct. Foods 2012, 4, 66–78. [Google Scholar] [CrossRef]
- Takeuchi, T.; Shimizu, H.; Ando, K.; Harada, E. Bovine lactoferrin reduces plasma triacylglycerol and NEFA accompanied by decreased hepatic cholesterol and triacylglycerol contents in rodents. Br. J. Nutr. 2004, 91, 533–538. [Google Scholar] [CrossRef]
- McManus, B.; Korpela, R.; O’Connor, P.; Schellekens, H.; Cryan, J.F.; Cotter, P.D.; Nilaweera, K.N. Compared to casein, bovine lactoferrin reduces plasma leptin and corticosterone and affects hypothalamic gene expression without altering weight gain or fat mass in high fat diet fed C57/BL6J mice. Nutr. Metab. 2015, 12, 53. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.-S.; Lin, C.-F.; Lee, C.-P.; Hsieh, M.-C.; Lu, H.-F.; Chen, Y.-F.; Ku, Y.-W.; Chen, P.-W. A Single Plasmid of Nisin-Controlled Bovine and Human Lactoferrin Expressing Elevated Antibacterial Activity of Lactoferrin-Resistant Probiotic Strains. Antibiotics 2021, 10, 120. [Google Scholar] [CrossRef] [PubMed]
- Chapman, C.M.C.; Gibson, G.R.; Rowland, I. Health benefits of probiotics: Are mixtures more effective than single strains? Eur. J. Nutr. 2011, 50, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Mohsin, M.; Abbas, R.Z.; Yin, G.; Sindhu, Z.-U.; Abbas, A.; Huang, Z.; Aleem, M.T.; Saeed, Z.; Afzal, M.Z.; Ejaz, A.; et al. Probiotics as therapeutic, antioxidant and immunomodulatory agents against poultry coccidiosis. World’s Poult. Sci. J. 2021, 77, 331–345. [Google Scholar] [CrossRef]
- Gu, Q.; Yin, Y.; Yan, X.; Liu, X.; Liu, F.; McClements, D.J. Encapsulation of multiple probiotics, synbiotics, or nutrabiotics for improved health effects: A review. Adv. Colloid Interface Sci. 2022, 102781. [Google Scholar] [CrossRef]
- Zheng, J.; Wittouck, S.; Salvetti, E.; Franz, C.M.A.P.; Harris, H.M.B.; Mattarelli, P.; O’Toole, P.W.; Pot, B.; Vandamme, P.; Walter, J.; et al. A taxonomic note on the genus Lactobacillus: Description of 23 novel genera, emended description of the genus Lactobacillus Beijerinck 1901, and union of Lactobacillaceae and Leuconostocaceae. Int. J. Syst. Evol. Microbiol. 2020, 70, 2782–2858. [Google Scholar] [CrossRef]
- Oligschlaeger, Y.; Shiri-Sverdlov, R. NAFLD Preclinical Models: More than a Handful, Less of a Concern? Biomedicines 2020, 8, 28. [Google Scholar] [CrossRef] [Green Version]
- Oseini, A.M.; Cole, B.K.; Issa, D.; Feaver, R.E.; Sanyal, A.J. Translating scientific discovery: The need for preclinical models of nonalcoholic steatohepatitis. Hepatol. Int. 2018, 12, 6–16. [Google Scholar] [CrossRef] [Green Version]
- Recena Aydos, L.; Aparecida do Amaral, L.; Serafim de Souza, R.; Jacobowski, A.C.; Freitas Dos Santos, E.; Rodrigues Macedo, M.L. Nonalcoholic Fatty Liver Disease Induced by High-Fat Diet in C57bL/6 Models. Nutrients 2019, 11, 3067. [Google Scholar] [CrossRef] [Green Version]
- Van Herck, M.A.; Vonghia, L.; Francque, S.M. Animal Models of Nonalcoholic Fatty Liver Disease—A Starter’s Guide. Nutrients 2017, 9, 1072. [Google Scholar] [CrossRef] [Green Version]
- Beppu, F.; Li, H.; Yoshinaga, K.; Nagai, T.; Yoshinda, A.; Kubo, A.; Kanda, J.; Gotoh, N. Dietary Starfish Oil Prevents Hepatic Steatosis and Hyperlipidemia in C57BL/6N Mice Fed High-fat Diet. J. Oleo Sci. 2017, 66, 761–769. [Google Scholar] [CrossRef] [PubMed]
- de Moura, E.D.M.; Dos Reis, S.A.; da Conceicao, L.L.; Sediyama, C.; Pereira, S.S.; de Oliveira, L.L.; Peluzio, M.d.C.G.; Martinez, J.A.; Milagro, F.I. Diet-induced obesity in animal models: Points to consider and influence on metabolic markers. Diabetol. Metab. Syndr. 2021, 13, 32. [Google Scholar] [CrossRef]
- Hashimoto, E.; Taniai, M.; Tokushige, K. Characteristics and diagnosis of NAFLD/NASH. J. Gastroenterol. Hepatol. 2013, 28, 64–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brunt, E.M.; Kleiner, D.E.; Wilson, L.A.; Belt, P.; Neuschwander-Tetri, B.A.; NASH Clinical Research Network (CRN). Nonalcoholic fatty liver disease (NAFLD) activity score and the histopathologic diagnosis in NAFLD: Distinct clinicopathologic meanings. Hepatology 2011, 53, 810–820. [Google Scholar] [CrossRef] [Green Version]
- Piazzolla, V.A.; Mangia, A. Noninvasive Diagnosis of NAFLD and NASH. Cells 2020, 9, 1005. [Google Scholar] [CrossRef] [PubMed]
- Brunt, E.M.; Janney, C.G.; Di Bisceglie, A.M.; Neuschwander-Tetri, B.A.; Bacon, B.R. Nonalcoholic steatohepatitis: A proposal for grading and staging the histological lesions. Am. J. Gastroenterol. 1999, 94, 2467–2474. [Google Scholar] [CrossRef]
- Mazzaccara, C.; Labruna, G.; Cito, G.; Scarfò, M.; De Felice, M.; Pastore, L.; Sacchetti, L. Age-Related Reference Intervals of the Main Biochemical and Hematological Parameters in C57BL/6J, 129SV/EV and C3H/HeJ Mouse Strains. PLoS ONE 2008, 3, e3772. [Google Scholar] [CrossRef] [PubMed]
- Leung, C.; Rivera, L.; Furness, J.B.; Angus, C.L.P.W. The role of the gut microbiota in NAFLD. Nat. Rev. Gastroenterol. Hepatol. 2016, 13, 412–425. [Google Scholar] [CrossRef]
- Aron-Wisnewsky, J.; Vigliotti, C.; Witjes, J.; Le, P.; Holleboom, A.G.; Verheij, J.; Nieuwdorp, M.; Clément, K. Gut microbiota and human NAFLD: Disentangling microbial signatures from metabolic disorders. Nat. Rev. Gastroenterol. Hepatol. 2020, 17, 279–297. [Google Scholar] [CrossRef]
- Lau, E.; Carvalho, D.; Freitas, P. Gut Microbiota: Association with NAFLD and Metabolic Disturbances. BioMed. Res. Int. 2015, 2015, 979515. [Google Scholar] [CrossRef]
- Ferolla, S.M.; Armiliato, G.N.; Couto, C.A.; Ferrari, T.C. Probiotics as a complementary therapeutic approach in nonalcoholic fatty liver disease. World J. Hepatol. 2015, 7, 559–565. [Google Scholar] [CrossRef]
- Meroni, M.; Longo, M.; Dongiovanni, P. The Role of Probiotics in Nonalcoholic Fatty Liver Disease: A New Insight into Therapeutic Strategies. Nutrients 2019, 11, 2642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bessone, F.; Razori, M.V.; Roma, M.G. Molecular pathways of nonalcoholic fatty liver disease development and progression. Cell. Mol. Life Sci. 2019, 76, 99–128. [Google Scholar] [CrossRef] [PubMed]
- Bugianesi, E.; Manzini, P.; D’Antico, S.; Vanni, E.; Longo, F.; Leone, N.; Massarenti, P.; Piga, A.; Marchesini, G.; Rizzetto, M. Relative contribution of iron burden, HFE mutations, and insulin resistance to fibrosis in nonalcoholic fatty liver. Hepatology 2004, 39, 179–187. [Google Scholar] [CrossRef] [Green Version]
- Fernández-Real, J.M.; Manco, M. Effects of iron overload on chronic metabolic diseases. Lancet Diabetes Endocrinol. 2014, 2, 513–526. [Google Scholar] [CrossRef]
- Nelson, J.E.; Wilson, L.; Brunt, E.M.; Yeh, M.M.; Kleiner, D.E.; Unalp-Arida, A.; Kowdley, K.V. Relationship between the pattern of hepatic iron deposition and histological severity in nonalcoholic fatty liver disease. Hepatology 2011, 53, 448–457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valenti, L.; Fracanzani, A.L.; Bugianesi, E.; Dongiovanni, P.; Galmozzi, E.; Vanni, E.; Canavesi, E.; Lattuada, E.; Roviaro, G.; Marchesini, G.; et al. HFE Genotype, Parenchymal Iron Accumulation, and Liver Fibrosis in Patients with Nonalcoholic Fatty Liver Disease. Gastroenterology 2010, 138, 905–912. [Google Scholar] [CrossRef] [Green Version]
- Guo, C.; Xue, H.; Guo, T.; Zhang, W.; Xuan, W.-Q.; Ren, Y.-T.; Wang, D.; Chen, Y.-H.; Meng, Y.-H.; Gao, H.-L.; et al. Recombinant human lactoferrin attenuates the progression of hepatosteatosis and hepatocellular death by regulating iron and lipid homeostasis in ob/ob mice. Food Funct. 2020, 11, 7183–7196. [Google Scholar] [CrossRef]
- Torres, S.; Fabersani, E.; Marquez, A.; Gauffin-Cano, P. Adipose tissue inflammation and metabolic syndrome. The proactive role of probiotics. Eur. J. Nutr. 2019, 58, 27–43. [Google Scholar] [CrossRef]
- Siciliano, R.; Reale, A.; Mazzeo, M.; Morandi, S.; Silvetti, T.; Brasca, M. Paraprobiotics: A New Perspective for Functional Foods and Nutraceuticals. Nutrients 2021, 13, 1225. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Placebo 1,2 | LAB 3 | LAB/LF 4 | LAB/LF (Inactivated) 5 | |
|---|---|---|---|---|
| Weight gain over study (g) | 1.06 ± 0.66 | 1.35 ± 0.94 | 1.66 ± 0.57 *,a | 0.81 ± 1.23 b |
| Weight gain (g/day) | 0.03 ± 0.02 | 0.04 ± 0.03 | 0.05 ± 0.02 *,a | 0.03 ± 0.04 b |
| Food intake (g/day) | 5.39 ± 0.14 | 6.13 ± 0.93 | 5.26 ± 0.11 | 5.6 ± 0.04 |
| Food efficiency ratio (%) 6 | 0.63 ± 0.38 | 0.72 ± 0.51 | 1.02 ± 0.34 *,a | 0.47 ± 0.7 b |
| Liver | Placebo 1 | LAB 1 | LAB/LF 1 | LAB/LF 1 (Inactivated) |
|---|---|---|---|---|
| Fatty change, general | 3.63 a | 1.75 b | 1.14 b | 1.25 b |
| Fatty change, microvesicles | 3.63 a | 1.63 b | 1.14 b | 1.25 b |
| Fatty change, macrovesicles | 1.13 a | 0.63 a | 0.50 b | 0.38 b |
| Inflammation | 0.75 a | 0.38 a | 0.13 b | 0.25 b |
| Kidney | ||||
| Fatty change, proximal tubule, multifocal | 1.63 a | 1.38 | 1.13 | 0.75 b |
| Epididymal fat | ||||
| Inflammation, multifocal | 0.88 a | 0.63 | 0.50 | 0.38 b |
| Placebo 1 | LAB 1 | LAB/LF 1 | LAB/LF (Inactivated) 1 | Normal Range 1 | |
|---|---|---|---|---|---|
| ALT (U/L) | 63.5 ± 9.2 | 53 ± 22.3 | 66.3 ± 23.7 | 48.2 ± 16.9 | 46–70 |
| AST (U/L) | 215 ± 148.49 | 113 ± 32.6 | 159.3 ± 74.4 | 124.2 ± 46.1 | 55–91 |
| UA (mg/dL) | 3.4 ± 0.85 | 3.13 ± 0.37 | 4.14 ± 1.10 | 3.05 ± 1.08 | 1.69–10.17 |
| CRE (mg/dL) | 0.25 ± 0.07 | 0.20 ± 0 a | 0.31 ± 0.07 b | 0.25 ± 0.05 a | 0.1–0.15 |
| GLU (mg/dL) | 179.5 ± 19.09 | 233.5 ± 84.04 a | 161 ± 22.55 b | 180.50 ± 33 | 100.9–163.96 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Z.-S.; Li, P.-L.; Ku, Y.-W.; Chen, P.-W. Oral Administration of Recombinant Lactoferrin-Expressing Probiotics Ameliorates Diet-Induced Lipid Accumulation and Inflammation in Non-Alcoholic Fatty Liver Disease in Mice. Microorganisms 2022, 10, 2215. https://doi.org/10.3390/microorganisms10112215
Liu Z-S, Li P-L, Ku Y-W, Chen P-W. Oral Administration of Recombinant Lactoferrin-Expressing Probiotics Ameliorates Diet-Induced Lipid Accumulation and Inflammation in Non-Alcoholic Fatty Liver Disease in Mice. Microorganisms. 2022; 10(11):2215. https://doi.org/10.3390/microorganisms10112215
Chicago/Turabian StyleLiu, Zhen-Shu, Pei-Lin Li, Yu-We Ku, and Po-Wen Chen. 2022. "Oral Administration of Recombinant Lactoferrin-Expressing Probiotics Ameliorates Diet-Induced Lipid Accumulation and Inflammation in Non-Alcoholic Fatty Liver Disease in Mice" Microorganisms 10, no. 11: 2215. https://doi.org/10.3390/microorganisms10112215