
Influence of Biochar on Soil Nutrients and Associated Rhizobacterial Communities of Mountainous Apple Trees in Northern Loess Plateau China




Abstract
:1. Introduction
2. Materials and Methods
2.1. Soil and Biochar
2.2. Experimental Design
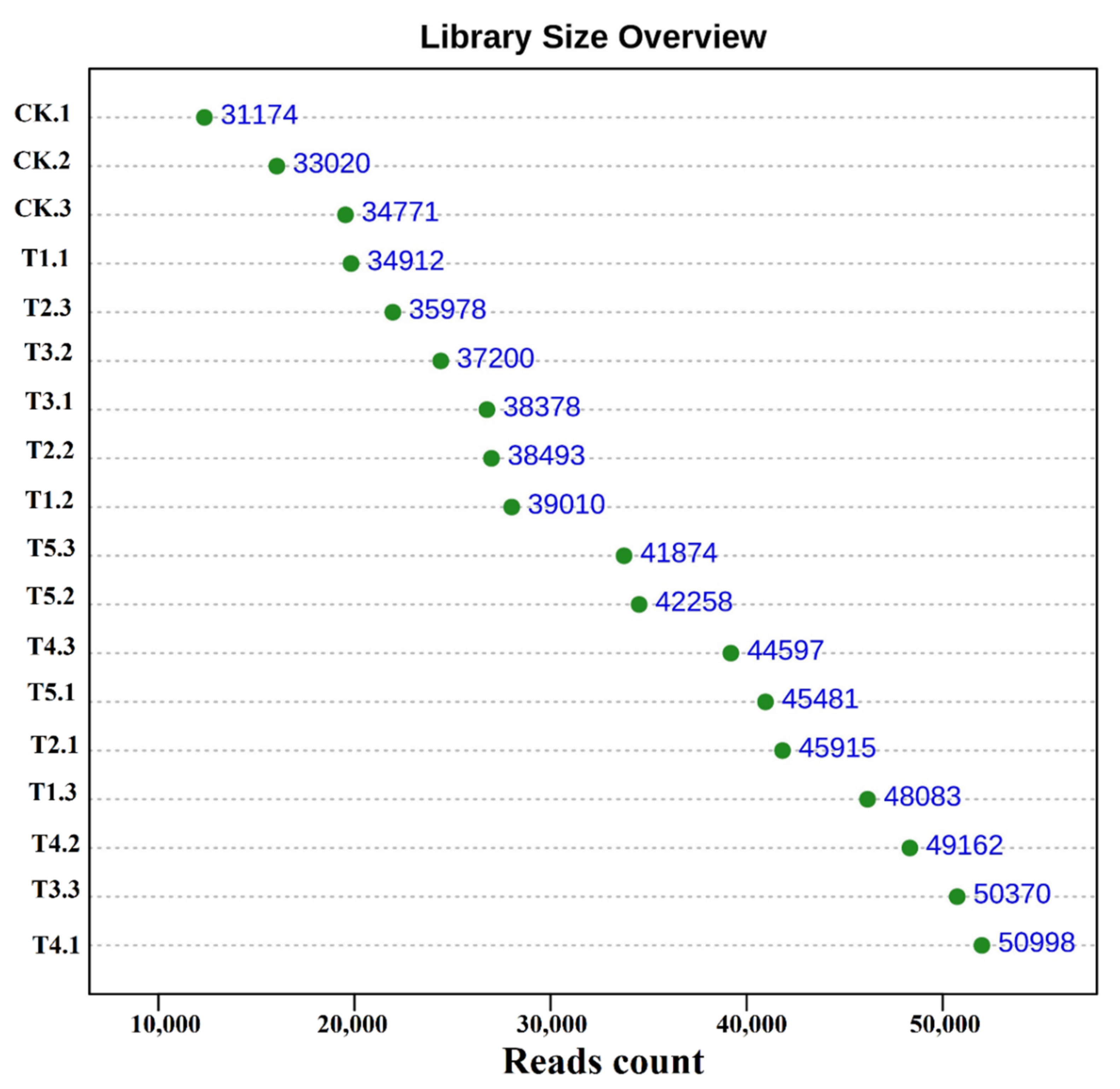
2.3. Sample Collection and Laboratory Analysis
2.4. Soil pH, Organic C, and N Analysis
2.5. Measurement of Soil Enzymes’ Activities
2.6. DNA Extraction and Sequencing
2.7. Processing of Illumina Sequencing Data
2.8. Alpha and Beta Diversity Analysis
2.9. Statistical Analysis
3. Results
3.1. Soil Physiochemical Properties and Enzymatic Activities
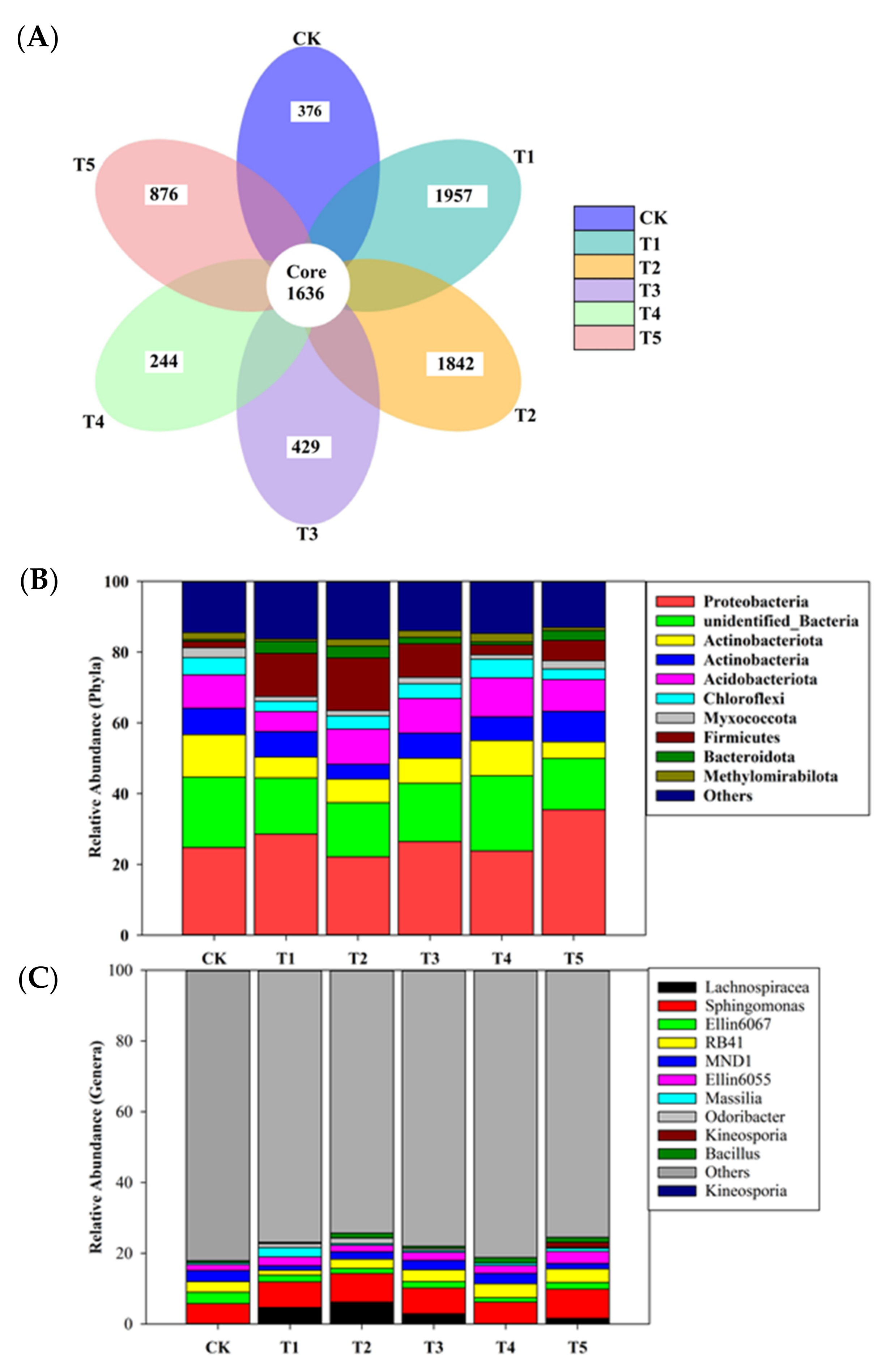
3.2. Composition and Community Structure of Rhizobacterial Microbiome
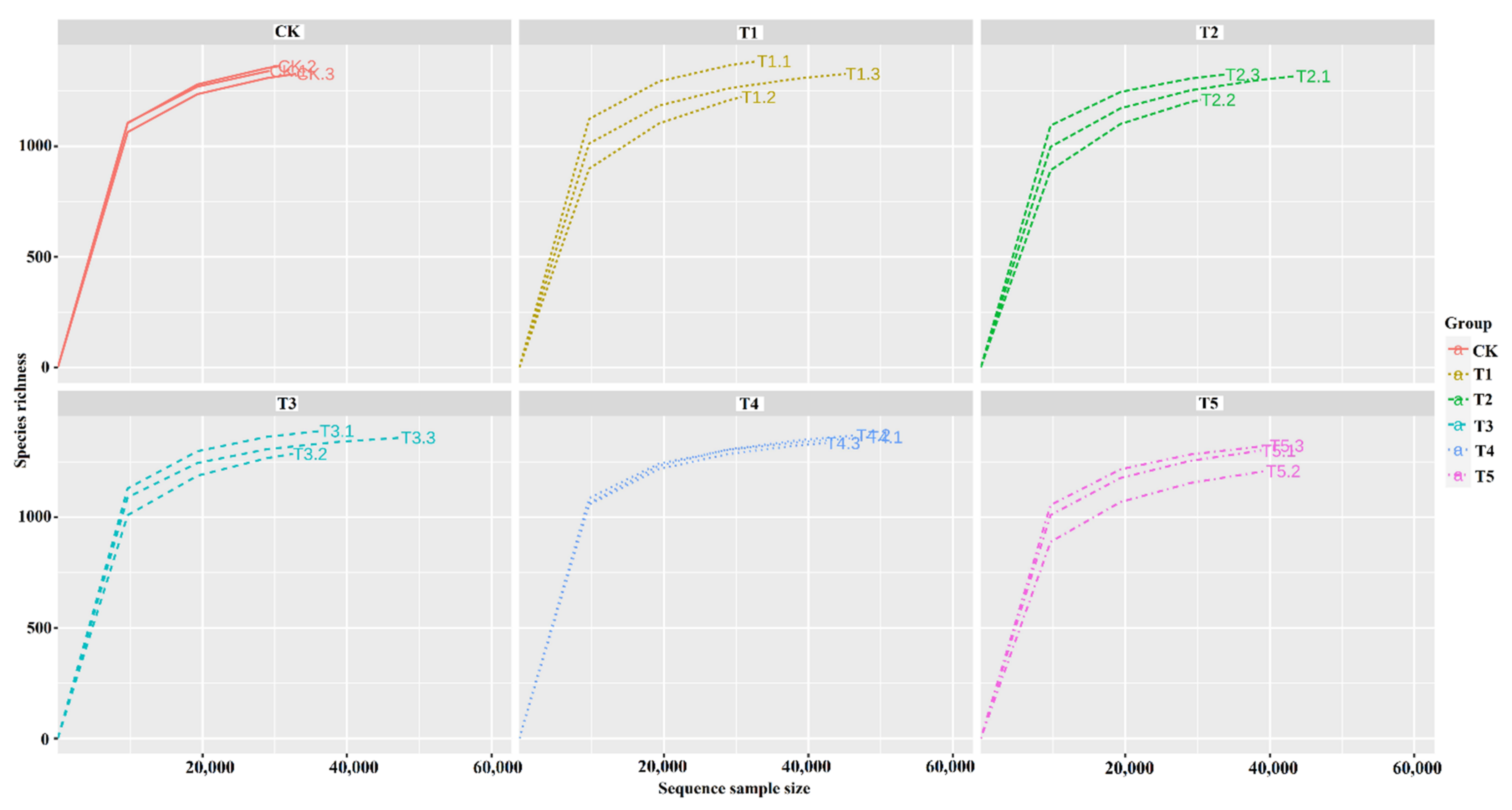
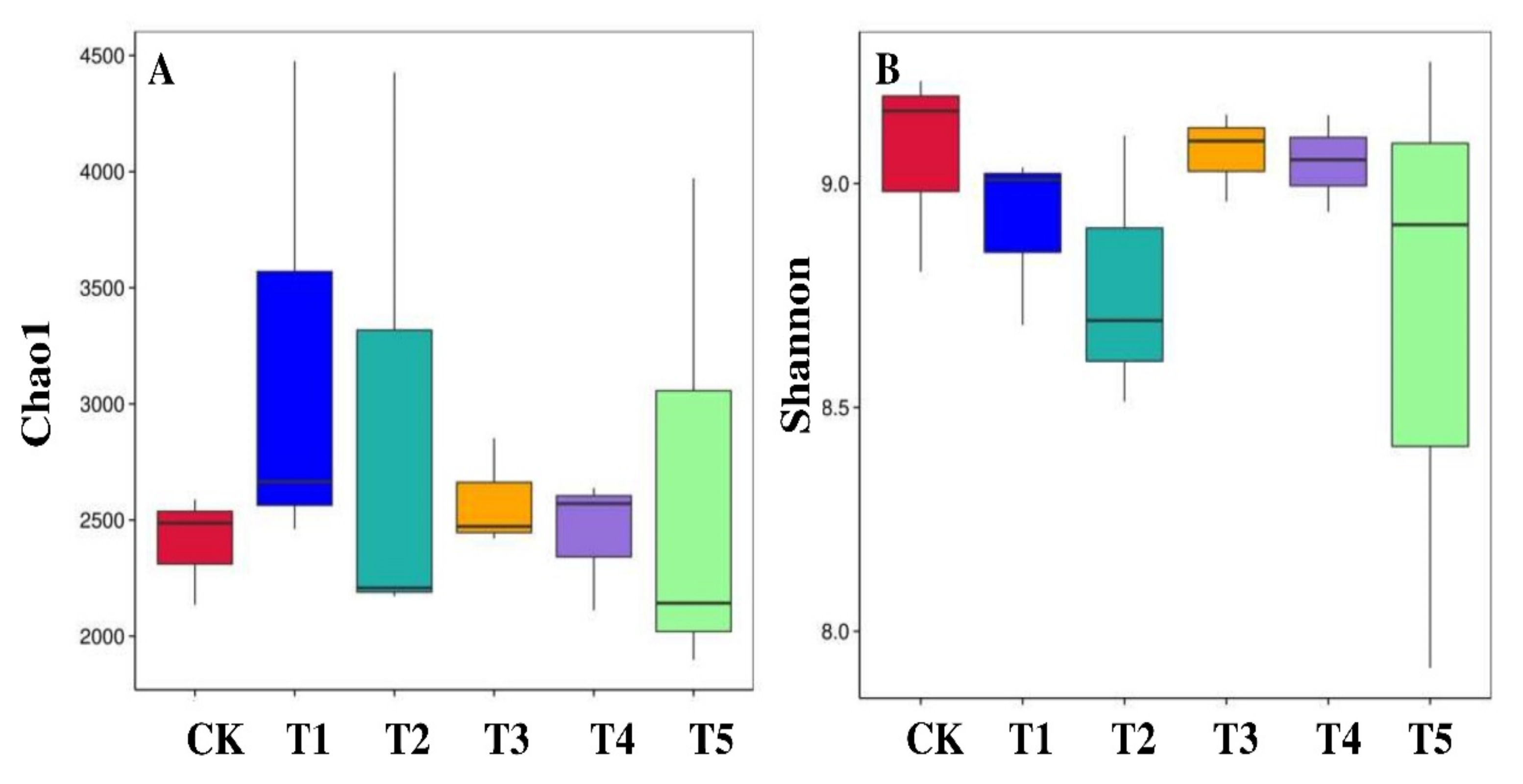
3.3. Diversity and Species Richness of Rhizobacterial Microbiome
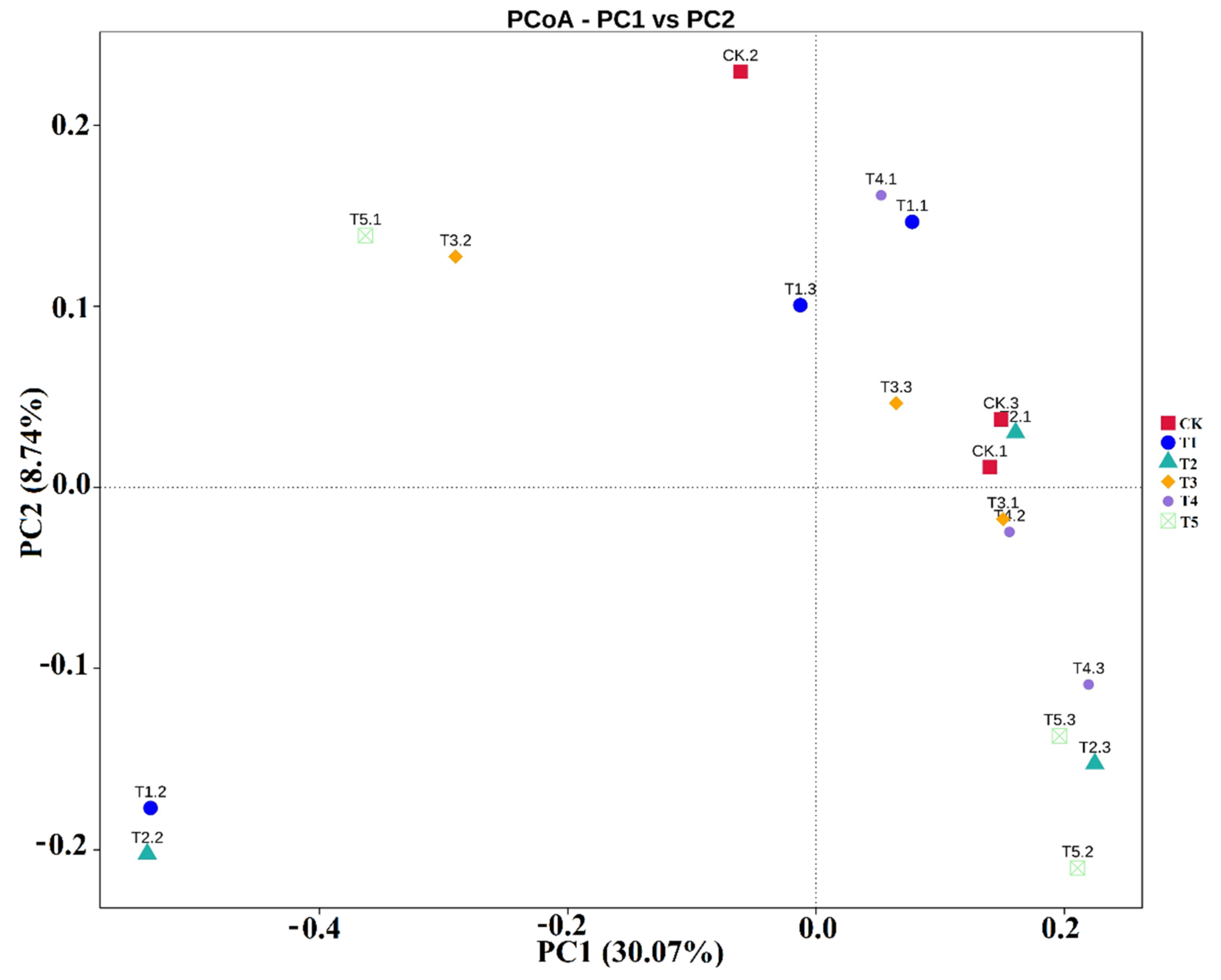
3.4. The Similarity of the Rhizobacterial Microbiomes
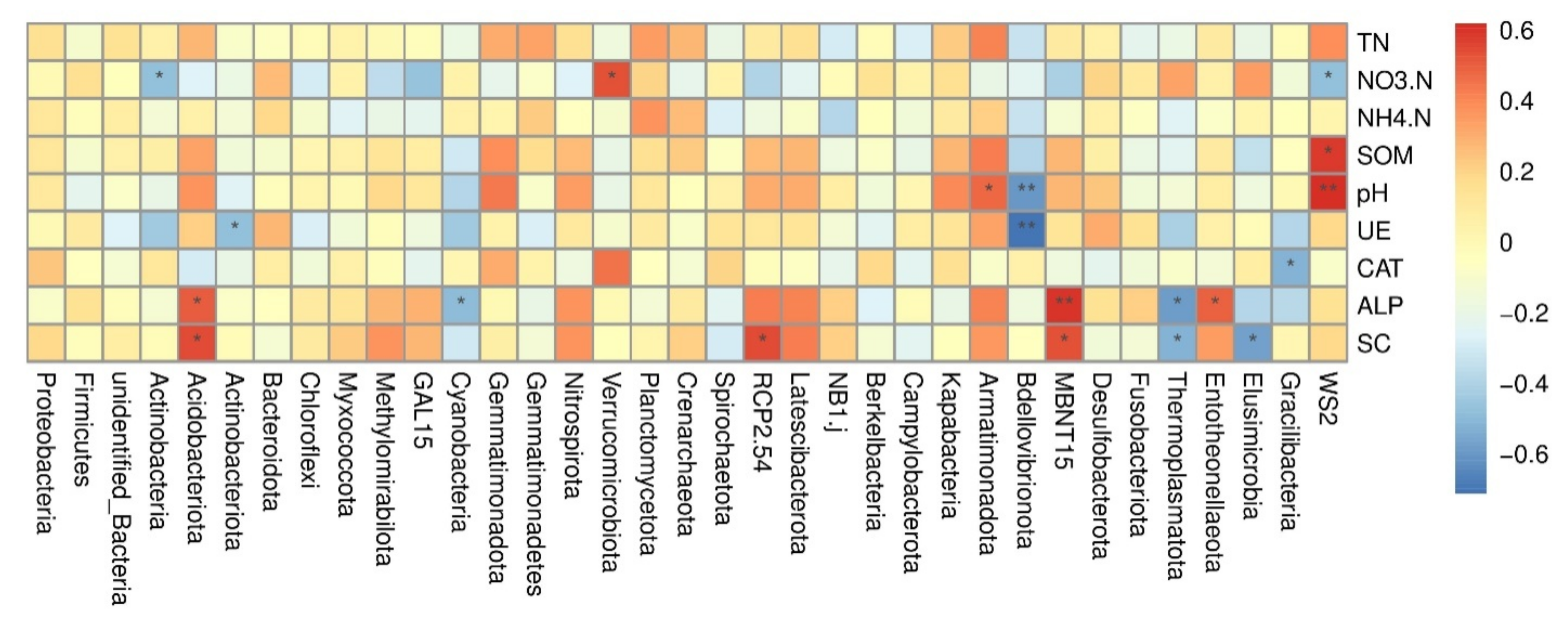
3.5. Spearmen Correlation Analysis among Major Bacterial Phyla and Environmental Factors
4. Discussion
4.1. Soil Physio-Biochemical Properties
4.2. Impacts of Biochar on Composition and Community Structure
4.3. Impact of Biochar on the Soil Bacterial Alpha and Beta Diversity
4.4. Correlation of Bacterial Communities and Environmental Factors
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hua, L.; Gao, J.; Zhou, M.; Bai, S. Impacts of Relative Elevation on Soil Nutrients and Apple Quality in the Hilly-Gully Region of the Loess Plateau, China. Sustainability 2021, 13, 1293. [Google Scholar] [CrossRef]
- Zhao, Z.; Zheng, W.; Ma, Y.; Wang, X.; Li, Z.; Zhai, B.; Wang, Z. Responses of soil water, nitrate and yield of apple orchard to integrated soil management in Loess Plateau, China. Agric. Water Manag. 2020, 240, 106325. [Google Scholar] [CrossRef]
- Zhu, Z.; Jia, Z.; Peng, L.; Chen, Q.; He, L.; Jiang, Y.; Ge, S. Life cycle assessment of conventional and organic apple production systems in China. J. Clean. Prod. 2018, 201, 156–168. [Google Scholar] [CrossRef]
- Wang, D.; Wang, L. Soil water dynamics in apple orchards of different ages on the Loess Plateau of China. Vadose Zone 2018, 17, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Chen, R.J. Spatial distribution and development tendency of China’s apple production. Food Nutr. China 2013, 19, 23–36. [Google Scholar]
- Wang, L.; Yang, F.; E, Y.; Yuan, J.; Raza, W.; Huang, Q.; Shen, Q. Long-term application of bioorganic fertilizers improved soil biochemical properties and microbial communities of an apple orchard soil. Front. Microbiol. 2016, 7, 1893. [Google Scholar] [CrossRef]
- Zoppolo, R.J.; Stefanelli, D.; Bird, G.W.; Perry, R.L. Soil properties under different orchard floor management systems for organic apple production. Org. Agric. 2011, 1, 231–246. [Google Scholar] [CrossRef]
- Wu, G.L.; Jia, C.; Huang, Z.; Lopez-Vicente, M.; Liu, Y. Plant litter crust appear as a promising measure to combat desertification in sandy land ecosystem. Catena 2021, 206, 105573. [Google Scholar] [CrossRef]
- Neilsen, G.; Forge, T.; Angers, D.; Neilsen, D.; Hogue, E. Suitable orchard floor management strategies in organic apple orchards that augment soil organic matter and maintain tree performance. Plant Soil 2014, 378, 325–335. [Google Scholar] [CrossRef]
- Wang, F.G.; Song, L.; Feng, Y.; Hong, Y.C.; Cui, D.J.; Yuan, Y.B. Characteristics of soil microbiology in different planting-life orchard acid soils. Chin. J. Soil Sci. 2011, 42, 46–50. [Google Scholar]
- Olego, M.Á.; Quiroga, M.J.; Mendaña-Cuervo, C.; Cara-Jiménez, J.; López, R.; Garzón-Jimeno, E. Long-term effects of calcium-based liming materials on soil fertility sustainability and rye production as soil quality indicators on a typic palexerult. Processes 2021, 9, 1181. [Google Scholar] [CrossRef]
- Torsvik, V.; Øvreås, L. Microbial diversity and function in soil: From genes to ecosystems. Curr. Opin. Microbiol. 2002, 5, 240–245. [Google Scholar] [CrossRef]
- Delgado-Baquerizo, M.; Reich, P.B.; Trivedi, C.; Eldridge, D.J.; Abades, S.; Alfaro, F.D.; Singh, B.K. Multiple elements of soil biodiversity drive ecosystem functions across biomes. Nat. Ecol. Evol. 2020, 4, 210–220. [Google Scholar] [CrossRef]
- Peck, G.M.; Merwin, I.A.; Thies, J.E.; Schindelbeck, R.R.; Brown, M.G. Soil properties change during the transition to integrated and organic apple production in a New York orchard. Appl. Soil. Ecol. 2011, 48, 18–30. [Google Scholar] [CrossRef]
- Rampelotto, P.H.; De Siqueira Ferreira, A.; Barboza, A.D.M.; Roesch, L.F.W. Changes in diversity, abundance, and structure of soil bacterial communities in Brazilian Savanna under different land use systems. Microb. Ecol. 2013, 66, 593–607. [Google Scholar] [CrossRef]
- Ali, I.; Yuan, P.; Ullah, S.; Iqbal, A.; Zhao, Q.; Liang, H.; Jiang, L. Biochar Amendment and Nitrogen Fertilizer Contribute to the Changes in Soil Properties and Microbial Communities in a Paddy Field. Front. Microbiol. 2022, 13, 834751. [Google Scholar] [CrossRef]
- Tecon, R.; Or, D. Biophysical processes supporting the diversity of microbial life in soil. FEMS Microbiol. Rev. 2017, 41, 599–623. [Google Scholar] [CrossRef] [Green Version]
- Gans, J.; Wolinsky, M.; Dunbar, J. Computational improvements reveal great bacterial diversity and high metal toxicity in soil. Science 2005, 309, 1387–1390. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Chen, D.; Shen, J.; Yuan, Q.; Fan, F.; Wei, W.; Wu, J. Biochar alters soil microbial communities and potential functions 3–4 years after amendment in a double rice cropping system. Agric. Ecosyst. Environ. 2021, 311, 107291. [Google Scholar] [CrossRef]
- Young, I.M.; Crawford, J.W. Interactions and self-organization in the soil-microbe complex. Science 2004, 304, 1634–1637. [Google Scholar] [CrossRef] [PubMed]
- Liang, B.; Ma, C.; Fan, L.; Wang, Y.; Yuan, Y. Soil amendment alters soil physicochemical properties and bacterial community structure of a replanted apple orchard. Microbiol. Res. 2018, 216, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Yao, S.; Merwin, I.A.; Abawi, G.S.; Thies, J.E. Soil fumigation and compost amendment alter soil microbial community composition but do not improve tree growth or yield in an apple replant site. Soil Biol. Biochem. 2006, 38, 587–599. [Google Scholar] [CrossRef]
- Sun, J.; Zhang, Q.; Zhou, J.; Wei, Q. Pyrosequencing technology reveals the impact of different manure doses on the bacterial community in apple rhizosphere soil. Appl. Soil Ecol. 2014, 78, 28–36. [Google Scholar] [CrossRef]
- Chen, Y.; Wen, X.; Sun, Y.; Zhang, J.; Wu, W.; Liao, Y. Mulching practices altered soil bacterial community structure and improved orchard productivity and apple quality after five growing seasons. Sci. Hortic. 2014, 172, 248–257. [Google Scholar] [CrossRef]
- Jien, S.H.; Wang, C.S. Effects of biochar on soil properties and erosion potential in a highly weathered soil. Catena 2013, 110, 225–233. [Google Scholar] [CrossRef] [Green Version]
- Shi, W.; Ju, Y.; Bian, R.; Li, L.; Joseph, S.; Mitchell, D.R.; Pan, G. Biochar bound urea boosts plant growth and reduces nitrogen leaching. Sci. Total Environ. 2020, 701, 134424. [Google Scholar] [CrossRef]
- Liu, M.; Linna, C.; Ma, S.; Ma, Q.; Guo, J.; Wang, F.; Wang, L. Effects of Biochar With Inorganic and Organic Fertilizers on Agronomic Traits and Nutrient Absorption of Soybean and Fertility and Microbes in Purple Soil. Front. Plant Sci. 2022, 13, 871021. [Google Scholar] [CrossRef]
- Ali, I.; Ullah, S.; He, L.; Zhao, Q.; Iqbal, A.; Wei, S.; Jiang, L. Combined application of biochar and nitrogen fertilizer improves rice yield, microbial activity and N-metabolism in a pot experiment. PeerJ 2020, 8, e10311. [Google Scholar] [CrossRef] [PubMed]
- Das, S.K.; Ghosh, G.K.; Avasthe, R. Applications of biomass derived biochar in modern science and technology. Environ. Technol. Innov. 2021, 21, 101306. [Google Scholar] [CrossRef]
- Qiang, M.; Gao, J.E.; Han, J.; Zhang, H.; Lin, T.; Long, S. How adding biochar improves loessal soil fertility and sunflower yield on consolidation project land on the Chinese loess plateau. Pol. J. Environ. Stud. 2020, 29, 3759–3769. [Google Scholar] [CrossRef]
- Khan, A.; Khan, S.; Lei, M.; Alam, M.; Khan, M.A.; Khan, A. Biochar characteristics, applications and importance in health risk reduction through metal immobilization. Environ. Technol. Innov. 2020, 20, 101121. [Google Scholar] [CrossRef]
- Kumar, M.; Xiong, X.; Wan, Z.; Sun, Y.; Tsang, D.C.; Gupta, J.; Ok, Y.S. Ball milling as a mechanochemical technology for fabrication of novel biochar nanomaterials. Bioresour. Technol. 2020, 312, 123613. [Google Scholar] [CrossRef] [PubMed]
- Kumar, M.; Dutta, S.; You, S.; Luo, G.; Zhang, S.; Show, P.L.; Tsang, D.C. A critical review on biochar for enhancing biogas production from anaerobic digestion of food waste and sludge. J. Clean. Prod. 2021, 305, 127143. [Google Scholar] [CrossRef]
- Dai, Z.; Xiong, X.; Zhu, H.; Xu, H.; Leng, P.; Li, J.; Xu, J. Association of biochar properties with changes in soil bacterial, fungal and fauna communities and nutrient cycling processes. Biochar 2021, 3, 239–254. [Google Scholar] [CrossRef]
- Huang, R.; Zhang, Z.; Xiao, X.; Zhang, N.; Wang, X.; Yang, Z.; Liang, Y. Structural changes of soil organic matter and the linkage to rhizosphere bacterial communities with biochar amendment in manure fertilized soils. Sci. Total Environ. 2019, 692, 333–343. [Google Scholar] [CrossRef]
- Ajeng, A.A.; Abdullah, R.; Ling, T.C.; Ismail, S.; Lau, B.F.; Ong, H.C.; Chang, J.S. Bioformulation of biochar as a potential inoculant carrier for sustainable agriculture. Environ. Technol. Innov. 2020, 20, 101168. [Google Scholar] [CrossRef]
- Calderón, F.J.; Benjamin, J.; Vigil, M.F. A comparison of corn (Zea mays L.) residue and its biochar on soil C and plant growth. PLoS ONE 2015, 10, e0121006. [Google Scholar] [CrossRef] [Green Version]
- Knox, O.G.G.; Oghoro, C.O.; Burnett, F.J.; Fountaine, J.M. Biochar increases soil pH, but is as ineffective as liming at controlling clubroot. Plant Pathol. J. 2015, 97, 149–152. [Google Scholar]
- Chen, L.F.; He, Z.B.; Wu, X.R.; Du, J.; Zhu, X.; Lin, P.F.; Kong, J.Q. Linkages between soil respiration and microbial communities following afforestation of alpine grasslands in the northeastern Tibetan Plateau. Appl. Soil Ecol. 2021, 161, 103882. [Google Scholar] [CrossRef]
- Guan, S.Y.; Zhang, D.; Zhang, Z. Soil Enzyme and Its Research Methods; China Agriculture Press: Beijing, China, 1986; pp. 274–297. [Google Scholar]
- Chen, B.; Du, K.; Sun, C.; Vimalanathan, A.; Liang, X.; Li, Y.; Shao, Y. Gut bacterial and fungal communities of the domesticated silkworm (Bombyx mori) and wild mulberry-feeding relatives. ISME J. 2018, 12, 2252–2262. [Google Scholar] [CrossRef] [Green Version]
- Zeng, X.Y.; Li, S.W.; Leng, Y.; Kang, X.H. Structural and functional responses of bacterial and fungal communities to multiple heavy metal exposure in arid loess. Sci. Total Environ. 2020, 723, 138081. [Google Scholar] [CrossRef]
- Gdanetz, K.; Benucci, G.M.N.; Vande Pol, N.; Bonito, G. CONSTAX: A tool for improved taxonomic resolution of environmental fungal ITS sequences. BMC Bioinform. 2017, 18, 538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, L.; Chen, H.; Wang, Y.; Mao, Q.; Zheng, M.; Su, Y.; Li, D. Responses of soil microbial resource limitation to multiple fertilization strategies. Soil Tillage Res. 2020, 196, 104474. [Google Scholar] [CrossRef]
- Dhariwal, A.; Chong, J.; Habib, S.; King, I.L.; Agellon, L.B.; Xia, J. Microbiome Analyst: A web-based tool for comprehensive statistical, visual and meta-analysis of microbiome data. Nucleic Acids Res. 2017, 45, W180–W188. [Google Scholar] [CrossRef] [PubMed]
- Palansooriya, K.N.; Sang, M.K.; Igalavithana, A.D.; Zhang, M.; Hou, D.; Oleszczuk, P.; Ok, Y.S. Biochar alters chemical and microbial properties of microplastic-contaminated soil. Environ. Res. 2022, 209, 112807. [Google Scholar] [CrossRef]
- Qu, J.; Yuan, Y.; Zhang, X.; Wang, L.; Tao, Y.; Jiang, Z.; Zhang, Y. Stabilization of lead and cadmium in soil by sulfur-iron functionalized biochar: Performance, mechanisms and microbial community evolution. J. Hazard. Mater. 2022, 425, 127876. [Google Scholar] [CrossRef] [PubMed]
- Pandian, K.; Subramaniayan, P.; Gnasekaran, P.; Chitraputhirapillai, S. Effect of biochar amendment on soil physical, chemical and biological properties and groundnut yield in rainfed Alfisol of semi-arid tropics. Arch. Agron. Soil Sci. 2016, 62, 1293–1310. [Google Scholar] [CrossRef]
- Zhang, C.; Liu, G.; Xue, S.; Wang, G. Soil bacterial community dynamics reflect changes in plant community and soil properties during the secondary succession of abandoned farmland in the Loess Plateau. Soil Biol. Biochem. 2016, 97, 40–49. [Google Scholar] [CrossRef]
- Ren, F.; Sun, N.; Xu, M.; Zhang, X.; Wu, L.; Xu, M. Changes in soil microbial biomass with manure application in cropping systems: A meta-analysis. Soil Tillage Res. 2019, 194, 104291. [Google Scholar] [CrossRef] [Green Version]
- Lehmann, J.; Rillig, M.C.; Thies, J.; Masiello, C.A.; Hockaday, W.C.; Crowley, D. Biochar effects on soil biota–a review. Soil Biol. Biochem. 2011, 43, 1812–1836. [Google Scholar] [CrossRef]
- Gul, S.; Whalen, J.K.; Thomas, B.W.; Sachdeva, V.; Deng, H. Physico-chemical properties and microbial responses in biochar-amended soils: Mechanisms and future directions. Agric. Ecosyst. Environ. 2015, 206, 46–59. [Google Scholar] [CrossRef]
- Gao, Y.; Lu, Y.; Lin, W.; Tian, J.; Cai, K. Biochar suppresses bacterial wilt of tomato by improving soil chemical properties and shifting soil microbial community. Microorganisms 2019, 7, 676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yao, X.H.; Min, H.; Lü, Z.H.; Yuan, H.P. Influence of acetamiprid on soil enzymatic activities and respiration. Eur. J. Soil Biol. 2006, 42, 120–126. [Google Scholar] [CrossRef]
- Eivazi, F.; Bayan, M.R.; Schmidt, K. Select soil enzyme activities in the historic Sanborn Field as affected by long-term cropping systems. Commun. Soil Sci. Plant Anal. 2003, 34, 2259–2275. [Google Scholar] [CrossRef]
- Das, S.K.; Varma, A. Role of enzymes in maintaining soil health. In Soil Enzymology; Springer: Berlin/Heidelberg, Germany, 2010; pp. 25–42. [Google Scholar] [CrossRef]
- Ma, H.; Egamberdieva, D.; Wirth, S.; Bellingrath-Kimura, S.D. Effect of biochar and irrigation on soybean-rhizobium symbiotic performance and soil enzymatic activity in field rhizosphere. Agronomy 2019, 9, 626. [Google Scholar] [CrossRef] [Green Version]
- Jindo, K.; Suto, K.; Matsumoto, K.; García, C.; Sonoki, T.; Sanchez-Monedero, M.A. Chemical and biochemical characterisation of biochar-blended composts prepared from poultry manure. Bioresour. Technol. 2012, 110, 396–404. [Google Scholar] [CrossRef] [PubMed]
- Makoi, J.H.; Ndakidemi, P.A. Selected soil enzymes: Examples of their potential roles in the ecosystem. Afr. J. Biotechnol. 2008, 7, 181–191. [Google Scholar]
- Zhao, D.; Li, F.; Wang, R. The effects of different urban land use patterns on soil microbial biomass nitrogen and enzyme activities in urban area of Beijing, China. Acta Ecol. Sin. 2012, 32, 144–149. [Google Scholar] [CrossRef]
- Bhaduri, D.; Saha, A.; Desai, D.; Meena, H.N. Restoration of carbon and microbial activity in salt-induced soil by application of peanut shell biochar during short-term incubation study. Chemosphere 2016, 148, 86–98. [Google Scholar] [CrossRef]
- Oleszczuk, P.; Jośko, I.; Futa, B.; Pasieczna-Patkowska, S.; Pałys, E.; Kraska, P. Effect of pesticides on microorganisms, enzymatic activity and plant in biochar-amended soil. Geoderma 2014, 214, 10–18. [Google Scholar] [CrossRef]
- Chen, J.; Liu, X.; Zheng, J.; Zhang, B.; Lu, H.; Chi, Z.; Yu, X. Biochar soil amendment increased bacterial but decreased fungal gene abundance with shifts in community structure in a slightly acid rice paddy from Southwest China. Appl. Soil Ecol. 2013, 71, 33–44. [Google Scholar] [CrossRef]
- Noyce, G.L.; Jones, T.; Fulthorpe, R.; Basiliko, N. Phosphorus uptake and availability and short-term seedling growth in three Ontario soils amended with ash and biochar. Can. J. Soil Sci. 2017, 97, 678–691. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Masto, R.E.; Ram, L.C.; Sarkar, P.; George, J.; Selvi, V.A. Biochar preparation from Parthenium hysterophorus and its potential use in soil application. Ecol. Eng. 2013, 55, 67–72. [Google Scholar] [CrossRef]
- Khadem, A.; Raiesi, F. Influence of biochar on potential enzyme activities in two calcareous soils of contrasting texture. Geoderma 2017, 308, 149–158. [Google Scholar] [CrossRef]
- Jiang, Y.; Wang, X.; Zhao, Y.; Zhang, C.; Jin, Z.; Shan, S.; Ping, L. Effects of Biochar Application on Enzyme Activities in Tea Garden Soil. Front. Bioeng. Biotechnol. 2021, 9, 728530. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Yao, S.; Bian, Y.; Jiang, X.; Song, Y. The combination of biochar and plant roots improves soil bacterial adaptation to PAH stress: Insights from soil enzymes, microbiome, and metabolome. J. Hazard. Mater. 2020, 400, 123227. [Google Scholar] [CrossRef] [PubMed]
- Hardy, B.; Sleutel, S.; Dufey, J.E.; Cornelis, J.T. The long-term effect of biochar on soil microbial abundance, activity and community structure is overwritten by land management. Front. Environ. Sci. 2019, 7, 110. [Google Scholar] [CrossRef] [Green Version]
- Ji, F.Y.; Ming, H.X.; Li, H.B.; Zan, S.J.; Wang, J.N.; Su, J.; Fan, J. Diversity of CO2 fixation gene in the surface waters of northern South China Sea in the Calvin cycle. Acta Sci. Circumst. 2016, 36, 4037–4043. [Google Scholar]
- Yin, D.; Li, H.; Wang, H.; Guo, X.; Wang, Z.; Lv, Y.; Lan, Y. Impact of different biochars on microbial community structure in the rhizospheric soil of rice grown in albic soil. Molecules 2021, 26, 4783. [Google Scholar] [CrossRef]
- Fierer, N.; Bradford, M.A.; Jackson, R.B. Toward an ecological classification of soil bacteria. Ecology 2007, 88, 1354–1364. [Google Scholar] [CrossRef]
- Kolton, M.; Meller Harel, Y.; Pasternak, Z.; Graber, E.R.; Elad, Y.; Cytryn, E. Impact of biochar application to soil on the root-associated bacterial community structure of fully developed greenhouse pepper plants. Appl. Environ. Microbiol. 2011, 77, 4924–4930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mackie, K.A.; Marhan, S.; Ditterich, F.; Schmidt, H.P.; Kandeler, E. The effects of biochar and compost amendments on copper immobilization and soil microorganisms in a temperate vineyard. Agric. Ecosyst. Environ. 2015, 201, 58–69. [Google Scholar] [CrossRef]
- Nielsen, S.; Minchin, T.; Kimber, S.; Van Zwieten, L.; Gilbert, J.; Munroe, P.; Thomas, T. Comparative analysis of the microbial communities in agricultural soil amended with enhanced biochars or traditional fertilizers. Agric. Ecosyst. Environ. 2014, 191, 73–82. [Google Scholar] [CrossRef]
- Cottrell, M.T.; Kirchman, D.L. Natural assemblages of marine proteobacteria and members of the Cytophaga-Flavobacter cluster consuming low-and high-molecular-weight dissolved organic matter. Appl. Environ. Microbiol. 2000, 66, 1692–1697. [Google Scholar] [CrossRef] [PubMed]
- Wei, M.; Liu, X.; He, Y.; Xu, X.; Wu, Z.; Yu, K.; Zheng, X. Biochar inoculated with Pseudomonas putida improves grape (Vitis vinifera L.) fruit quality and alters bacterial diversity. Rhizosphere 2020, 16, 100261. [Google Scholar] [CrossRef]
- He, Y.; Wu, Z.; Wang, W.; Liu, X.; Ye, B.C. Bacterial community and phosphorus species changes in pepper rhizosphere soils after Pseudomonas putida Rs-198 inoculation. Rhizosphere 2019, 11, 100164. [Google Scholar] [CrossRef]
- Nguyen, T.T.N.; Wallace, H.M.; Xu, C.Y.; Zwieten, L.V.; Weng, Z.H.; Xu, Z.; Bai, S.H. The effects of short term, long term and reapplication of biochar on soil bacteria. Sci. Total Environ. 2018, 636, 142–151. [Google Scholar] [CrossRef]
- Khan, A.; Jiang, H.; Bu, J.; Adnan, M.; Gillani, S.W.; Hussain, M.A.; Zhang, M. Untangling the Rhizosphere Bacterial Community Composition and Response of Soil Physiochemical Properties to Different Nitrogen Applications in Sugarcane Field. Front. Microbiol. 2022, 13, 856078. [Google Scholar] [CrossRef]
- Zhu, J.; Cao, A.; Wu, J.; Fang, W.; Huang, B.; Yan, D.; Li, Y. Effects of chloropicrin fumigation combined with biochar on soil bacterial and fungal communities and Fusarium oxysporum. Ecotoxicol. Environ. 2021, 220, 112414. [Google Scholar] [CrossRef]
- Wang, Q.; Chen, Z.; Zhao, J.; Ma, J.; Yu, Q.; Zou, P.; Ma, J. Fate of heavy metals and bacterial community composition following biogas slurry application in a single rice cropping system. J. Soils Sediments 2022, 22, 968–981. [Google Scholar] [CrossRef]
- Xia, S.; Shi, Y.; Fu, Y.; Ma, X. DGGE analysis of 16S rDNA of ammonia-oxidizing bacteria in chemical–biological flocculation and chemical coagulation systems. Appl. Microbiol. Biotechnol. 2005, 69, 99–105. [Google Scholar] [CrossRef]
- Ye, J.; Zhang, R.; Nielsen, S.; Joseph, S.D.; Huang, D.; Thomas, T. A combination of biochar–mineral complexes and compost improves soil bacterial processes, soil quality, and plant properties. Front. Microbiol. 2016, 7, 372. [Google Scholar] [CrossRef]
- Chuang, H.-P.; Imachi, H.; Tandukar, M.; Kawakami, S.; Harada, H.; Ohashi, A. Microbial community that catalyzes partial nitrification at low oxygen atmosphere as revealed by 16S rRNA and amoA genes. J. Biosci. Bioeng. 2007, 104, 525–528. [Google Scholar] [CrossRef] [PubMed]
- Kolton, M.; Graber, E.R.; Tsehansky, L.; Elad, Y.; Cytryn, E. Biochar-stimulated plant performance is strongly linked to microbial diversity and metabolic potential in the rhizosphere. New Phytol. 2017, 213, 1393–1404. [Google Scholar] [CrossRef] [PubMed]
- He, L.L.; Zhong, Z.K.; Yang, H.M. Effects on soil quality of biochar and straw amendment in conjunction with chemical fertilizers. J. Integr. Agric. 2017, 16, 704–712. [Google Scholar] [CrossRef] [Green Version]
- Gao, M.; Yang, J.; Liu, C.; Gu, B.; Han, M.; Li, J.; Li, N.; Liu, N.; An, N.; Dai, J.; et al. Effects of long-term biochar and biochar-based fertilizer application on brown earth soil bacterial communities. Agric. Ecosyst. Environ. 2021, 309, 107285. [Google Scholar] [CrossRef]
- Abujabhah, I.S.; Bound, S.A.; Doyle, R.; Bowman, J.P. Effects of biochar and compost amendments on soil physicochemical properties and the total community within a temperate agricultural soil. Appl. Soil Ecol. 2016, 98, 243–253. [Google Scholar] [CrossRef]
- Yao, Q.; Liu, J.; Yu, Z.; Li, Y.; Jin, J.; Liu, X.; Wang, G. Three years of biochar amendment alters soil physiochemical properties and fungal community composition in a black soil of northeast China. Soil Biol. Biochem. 2017, 110, 56–67. [Google Scholar] [CrossRef]
- Lin, Y.; Ye, Y.; Liu, S.; Wen, J.; Chen, D. Effect Mechanism of Land Consolidation on Soil Bacterial Community: A Case Study in Eastern China. Int. J. Environ. Res. Public Health 2022, 19, 845. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Ullah, F.; Ahmad, R.; Ali Shah, S.U.; Khan, A.; Adnan, M. Response of Soil Proteobacteria to Biochar Amendment in Sustainable Agriculture—A Mini Review. J. Soil Plant Environ. 2022, 1, 16–30. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PCR Reaction System (50 µL) | Addition (µL) |
|---|---|
| 2X Premix Taq | 25 |
| Forward primer (5 µM) | 1 |
| Reverse primer (5 µM) | 1 |
| DNA Template | 3 |
| Deionized distilled H2O | 20 |
| Stages | No. of Cycle | Temperature (°C) | Time (min) |
|---|---|---|---|
| Initial denaturation | 1 | 94 | 5 |
| Second denaturation | 30 | 94 | 0.3 |
| Annealing | 1 | 52 | 0.3 |
| Initial extension | 1 | 72 | 0.3 |
| Final extension | 1 | 72 | 10 |
| 3 Months | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Treat | Soil pH | SOM | STN | NO3−-N | NH4+-N | UR | CAT | ALP | SC |
| Ck | 8.91 ± 0.06 a | 8.48 ± 0.19 a | 0.42 ± 0.05 a | 3.93 ± 0.15 a | 67.50 ± 3.90 a | 0.30 ± 0.03 a | 3.95 ± 0.09 a | 0.99 ± 0.05 a | 6.11 ± 0.35 a |
| T1 | 8.93 ± 0.14 a | 8.75 ± 0.45 a | 0.53 ± 0.06 abc | 4.43 ± 0.82 a | 71.68 ± 2.39 a | 0.33 ± 0.05 a | 4.01 ± 0.07 ab | 1.09 ± 0.03 ab | 6.13 ± 0.40 a |
| T2 | 8.98 ± 0.05 ab | 9.41 ± 0.30 b | 0.47 ± 0.07 ab | 6.43 ± 1.02 b | 78.69 ± 3.33 a | 0.51 ± 0.03 c | 4.09 ± 0.05 b | 1.12 ± 0.04 bc | 6.45 ± 0.16 ab |
| T3 | 9.03 ± 0.02 ab | 10.30 ± 0.15 c | 0.55 ± 0.06 abc | 7.63 ± 1.04 b | 81.47 ± 5.20 b | 0.42 ± 0.06 b | 4.14 ± 0.06 bc | 1.19 ± 0.02 bc | 6.70 ± 1.18 ab |
| T4 | 9.09 ± 0.02 b | 10.89 ± 0.22 d | 0.61 ± 0.13 bc | 9.43 ± 0.72 c | 83.82 ± 2.50 b | 0.45 ± 0.05 bc | 4.25 ± 0.04 c | 1.22 ± 0.09 cd | 7.11 ± 0.44 ab |
| T5 | 9.11 ± 0.06 b | 11.52 ± 0.22 e | 0.67 ± 0.10 c | 10.42 ± 1.13 c | 84.77 ± 2.64 b | 0.53 ± 0.03 c | 4.30 ± 0.05 cd | 1.31 ± 0.06 d | 7.52 ± 0.27 b |
| 6 Months | |||||||||
| Ck | 8.95 ± 0.06 a | 8.56 ± 0.26 a | 0.46 ± 0.04 a | 3.98 ± 0.14 a | 68.20 ± 5.03 a | 0.32 ± 0.03 a | 3.94 ± 0.05 a | 1.02 ± 0.02 a | 6.15 ± 0.49 a |
| T1 | 8.98 ± 0.06 ab | 8.78 ± 0.39 a | 0.52 ± 0.07 ab | 4.49 ± 0.20 a | 73.44 ± 8.14 a | 0.35 ± 0.07 ab | 4.05 ± 0.07 b | 1.12 ± 0.03 ab | 6.20 ± 0.52 a |
| T2 | 9.02 ± 0.07 abc | 9.95 ± 0.35 b | 0.55 ± 0.07 abc | 6.60 ± 0.84 b | 81.95 ± 2.49 b | 0.54 ± 0.04 d | 4.09 ± 0.03 b | 1.17 ± 0.04 bc | 6.58 ± 0.22 ab |
| T3 | 9.08 ± 0.04 bc | 10.62 ± 0.55 bc | 0.59 ± 0.05 bc | 7.77 ± 1.11 b | 84.04 ± 4.51 b | 0.43 ± 0.06 bc | 4.18 ± 0.06 c | 1.22 ± 0.03 bcd | 6.79 ± 1.13 ab |
| T4 | 9.11 ± 0.04 c | 11.28 ± 0.17 cd | 0.67 ± 0.08 cd | 9.55 ± 0.58 c | 88.52 ± 1.50 b | 0.49 ± 0.06 cd | 4.27 ± 0.05 cd | 1.27 ± 0.10 cd | 7.28 ± 0.93 ab |
| T5 | 9.14 ± 0.08 c | 11.69 ± 0.50 d | 0.74 ± 0.06 d | 10.58 ± 1.55 c | 88.57 ± 3.29 b | 0.57 ± 0.05 d | 4.32 ± 0.06 d | 1.33 ± 0.07 d | 7.85 ± 1.22 b |
| 9 Months | |||||||||
| Ck | 8.99 ± 0.01 a | 8.75 ± 0.33 a | 0.49 ± 0.05 a | 4.10 ± 0.33 a | 69.13 ± 14.07 a | 0.34 ± 0.04 a | 3.93 ± 0.09 a | 1.03 ± 0.06 a | 6.18 ± 1.11 a |
| T1 | 9.04 ± 0.03 a | 8.86 ± 1.45 a | 0.57 ± 0.08 a | 4.53 ± 1.16 a | 74.79 ± 7.56 ab | 0.36 ± 0.03 a | 4.04 ± 0.07 ab | 1.13 ± 0.09 ab | 6.24 ± 0.39 a |
| T2 | 9.06 ± 0.03 ab | 10.19 ± 0.28 b | 0.56 ± 0.07 a | 6.73 ± 0.92 b | 84.06 ± 4.54 bc | 0.60 ± 0.07 c | 4.13 ± 0.05 bc | 1.21 ± 0.03 bc | 6.67 ± 0.41 b |
| T3 | 9.13 ± 0.01 bc | 11.56 ± 0.19 c | 0.62 ± 0.03 ab | 8.09 ± 1.68 bc | 85.12 ± 3.94 bc | 0.47 ± 0.05 b | 4.20 ± 0.06 cd | 1.27 ± 0.04 bc | 6.82 ± 1.06 b |
| T4 | 9.16 ± 0.09 bc | 11.68 ± 0.40 c | 0.71 ± 0.09 bc | 9.78 ± 1.22 cd | 89.66 ± 2.81 c | 0.52 ± 0.06 bc | 4.31 ± 0.07 de | 1.32 ± 0.14 c | 7.43 ± 2.03 bc |
| T5 | 9.19 ± 0.08 c | 12.03 ± 0.57 c | 0.81 ± 0.07 c | 10.82 ± 0.69 d | 90.30 ± 4.82 c | 0.61 ± 0.04 c | 4.39 ± 0.09 e | 1.35 ± 0.08 c | 7.97 ± 0.87 c |
| 12 Months | |||||||||
| Ck | 9.01 ± 0.01 a | 8.82 ± 0.21 a | 0.53 ± 0.06 a | 4.14 ± 1.82 a | 69.81 ± 30.94 a | 0.35 ± 0.05 a | 3.95 ± 0.11 a | 1.06 ± 0.03 a | 6.20 ± 1.37 a |
| T1 | 9.08 ± 0.04 ab | 8.92 ± 1.52 a | 0.59 ± 0.16 ab | 4.64 ± 2.66 a | 76.16 ± 9.01 a | 0.38 ± 0.05 a | 4.08 ± 0.12 ab | 1.19 ± 0.28 a | 6.33 ± 1.35 a |
| T2 | 9.10 ± 0.05 abc | 10.47 ± 0.56 b | 0.59 ± 0.09 ab | 7.32 ± 3.10 ab | 86.56 ± 5.01 b | 0.63 ± 0.06 d | 4.15 ± 0.11 abc | 1.33 ± 0.09 b | 6.83 ± 0.25 b |
| T3 | 9.16 ± 0.01 bc | 11.65 ± 0.27 bc | 0.65 ± 0.07 ab | 8.52 ± 5.20 ab | 87.15 ± 8.11 b | 0.51 ± 0.04 b | 4.24 ± 0.11 bcd | 1.31 ± 0.09 b | 6.90 ± 1.16 b |
| T4 | 9.19 ± 0.12 bc | 12.09 ± 0.27 c | 0.73 ± 0.12 bc | 10.13 ± 3.41 ab | 91.11 ± 4.58 b | 0.58 ± 0.10 bc | 4.33 ± 0.09 cd | 1.35 ± 0.13 b | 7.58 ± 0.94 c |
| T5 | 9.22 ± 0.09 c | 12.38 ± 0.15 c | 0.87 ± 0.08 c | 11.17 ± 2.85 b | 92.97 ± 5.94 b | 0.65 ± 0.04 d | 4.40 ± 0.08 d | 1.41 ± 0.31 bc | 8.04 ± 1.41 cd |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ahmad, R.; Gao, J.; Gao, Z.; Khan, A.; Ali, I.; Fahad, S. Influence of Biochar on Soil Nutrients and Associated Rhizobacterial Communities of Mountainous Apple Trees in Northern Loess Plateau China. Microorganisms 2022, 10, 2078. https://doi.org/10.3390/microorganisms10102078
Ahmad R, Gao J, Gao Z, Khan A, Ali I, Fahad S. Influence of Biochar on Soil Nutrients and Associated Rhizobacterial Communities of Mountainous Apple Trees in Northern Loess Plateau China. Microorganisms. 2022; 10(10):2078. https://doi.org/10.3390/microorganisms10102078
Chicago/Turabian StyleAhmad, Rafiq, Jianen Gao, Zhe Gao, Abdullah Khan, Izhar Ali, and Shah Fahad. 2022. "Influence of Biochar on Soil Nutrients and Associated Rhizobacterial Communities of Mountainous Apple Trees in Northern Loess Plateau China" Microorganisms 10, no. 10: 2078. https://doi.org/10.3390/microorganisms10102078